
HOXB9 Overexpression Promotes Colorectal Cancer Progression and Is Associated with Worse Survival in Liver Resection Patients for Colorectal Liver Metastases

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Gene Expression Bioinformatics Analysis
2.2. Gene Expression Editing Mechanistic Studies
2.3. In Silico Transcriptional Regulation Prediction of HOXB9 and Gene Set Enrichment Analysis
2.4. Patient Tissue Samples, Clinicopathological Variables and Immunohistochemistry
3. Results
3.1. HOXB9 Differential Expression in CRC
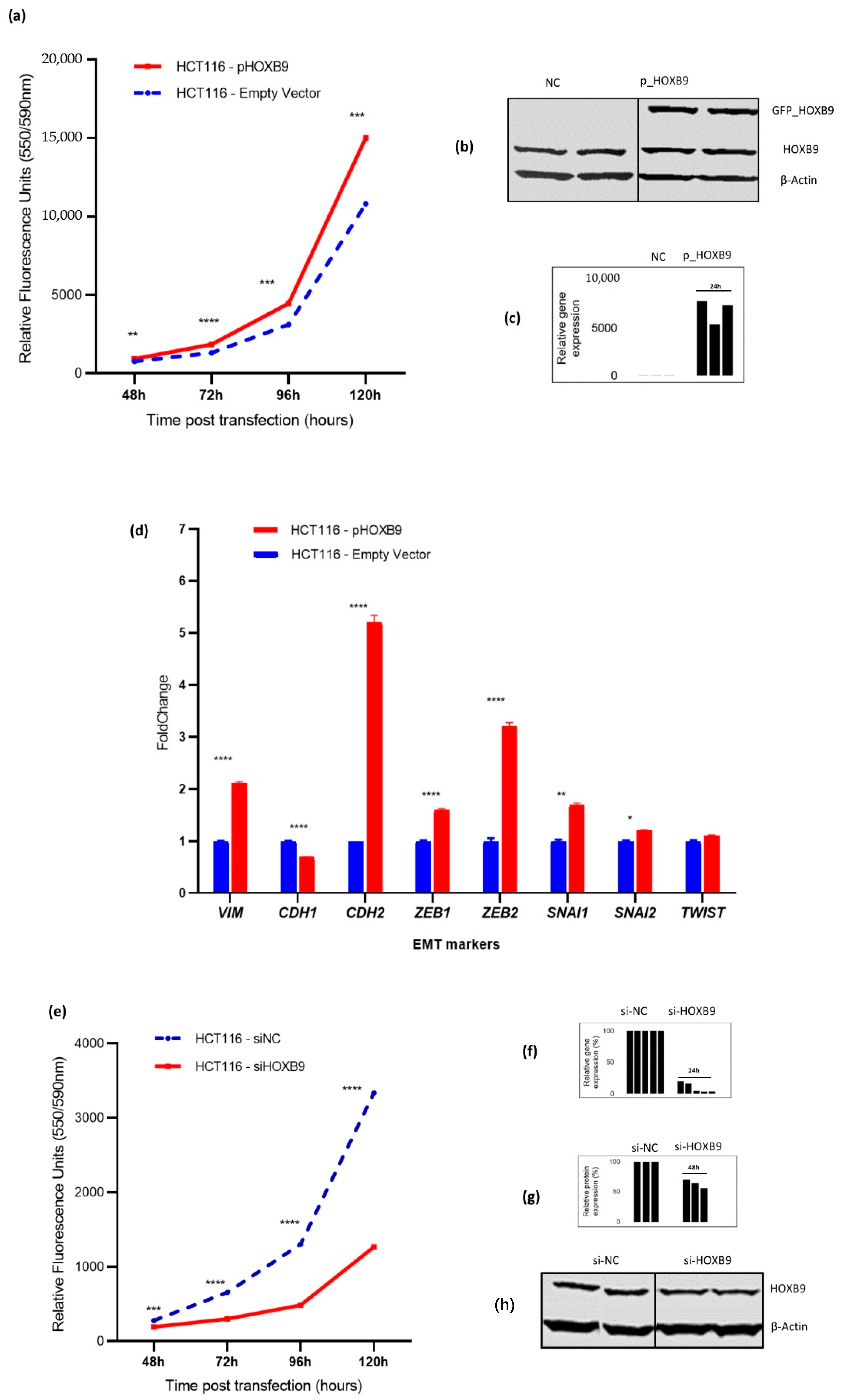
3.2. Impact of HOXB9 Dysregulation in CRC Progression In Vitro
3.3. Predicted HOXB9 Regulators and Related Biological Processes
3.4. Association of HOXB9 with OS in Patients with CRLM
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Chow, F.C.-L.; Chok, K.S.-H. Colorectal liver metastases: An update on multidisciplinary approach. World J. Hepatol. 2019, 11, 150–172. [Google Scholar] [CrossRef]
- Singh, M.P.; Rai, S.; Pandey, A.; Singh, N.K.; Srivastava, S. Molecular subtypes of colorectal cancer: An emerging therapeutic opportunity for personalized medicine. Genes Dis. 2021, 8, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Liu, L.; Li, W.; Zou, D.; Yu, J.; Wang, L.; Wong, C.C. Transcription factors in colorectal cancer: Molecular mechanism and therapeutic implications. Oncogene 2021, 40, 1555–1569. [Google Scholar] [CrossRef]
- Castelli-Gair Hombría, J.; Lovegrove, B. Beyond homeosis-HOX function in morphogenesis and organogenesis. Differentiation 2003, 71, 461–476. [Google Scholar] [CrossRef]
- Li, B.; Huang, Q.; Wei, G.-H. The Role of HOX Transcription Factors in Cancer Predisposition and Progression. Cancers 2019, 11, 528. [Google Scholar] [CrossRef] [Green Version]
- Paço, A.; de Bessa Garcia, S.A.; Castro, J.L.; Costa-Pinto, A.; Freitas, R. Roles of the HOX Proteins in Cancer Invasion and Metastasis. Cancers 2020, 13, 10. [Google Scholar] [CrossRef] [PubMed]
- Contarelli, S.; Fedele, V.; Melisi, D. HOX Genes Family and Cancer: A Novel Role for Homeobox B9 in the Resistance to Anti-Angiogenic Therapies. Cancers 2020, 12, 3299. [Google Scholar] [CrossRef]
- Paschos, K.A.; Majeed, A.W.; Bird, N.C. Natural history of hepatic metastases from colorectal cancer - Pathobiological pathways with clinical significance. World J. Gastroenterol. 2014, 20, 3719–3737. [Google Scholar] [CrossRef] [PubMed]
- Martinou, E.; Falgari, G.; Bagwan, I.; Angelidi, A.M. A Systematic Review on HOX Genes as Potential Biomarkers in Colorectal Cancer: An Emerging Role of HOXBInt. J. Mol. Sci. 2021, 22, 13429. [Google Scholar] [CrossRef]
- Huang, K.; Yuan, R.; Wang, K.; Hu, J.; Huang, Z.; Yan, C.; Shen, W.; Shao, J. Overexpression of HOXB9 promotes metastasis and indicates poor prognosis in colon cancer. Chin. J. Cancer Res. 2014, 26, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, Y.; Hayashida, T.; Hirata, A.; Takahashi, H.; Chiba, N.; Ohmura, M.; Wakui, M.; Jinno, H.; Hasegawa, H.; Maheswaran, S.; et al. Bevacizumab terminates homeobox B9-induced tumor proliferation by silencing microenvironmental communication. Mol. Cancer 2014, 13, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, J.; Niu, M.; Wang, P.; Zhu, X.; Li, S.; Song, J.; He, H.; Wang, Y.; Xue, L.; Fang, W.; et al. Elevated HOXB9 expression promotes differentiation and predicts a favourable outcome in colon adenocarcinoma patients. Br. J. Cancer 2014, 111, 883–893. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, D.R.; Kalyana-Sundaram, S.; Mahavisno, V.; Varambally, R.; Yu, J.; Briggs, B.B.; Barrette, T.R.; Anstet, M.J.; Kincead-Beal, C.; Kulkarni, P.; et al. Oncomine 3.0: Genes, Pathways, and Networks in a Collection of 18,000 Cancer Gene Expression Profiles. Neoplasia 2007, 9, 166–180. [Google Scholar] [CrossRef] [Green Version]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.V.S.K.; Varambally, S. UALCAN: A portal for facilitating tumor subgroup gene expression and survival analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef]
- Wu, C.; Huang, B.E.; Chen, G.; Lovenberg, T.W.; Pocalyko, D.J.; Yao, X. Integrative Analysis of DiseaseLand Omics Database for Disease Signatures and Treatments: A Bipolar Case Study. Front. Genet. 2019, 10, 10–3389. [Google Scholar] [CrossRef]
- Ahmed, D.; Eide, P.W.; Eilertsen, I.A.; Danielsen, S.A.; Eknaes, M.; Hektoen, M.; Lind, G.E.; Lothe, R.A. Epigenetic and genetic features of 24 colon cancer cell lines. Oncogenesis 2013, 2, e71. [Google Scholar] [CrossRef]
- Menyhárt, O.; Harami-Papp, H.; Sukumar, S.; Schäfer, R.; Magnani, L.; de Barrios, O.; Győrffy, B. Guidelines for the selection of functional assays to evaluate the hallmarks of cancer. Biochim. Biophys. Acta Bioenerg. 2016, 1866, 300–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zachari, M.; Chondrou, P.S.; Pouliliou, S.; Mitrakas, A.G.; Abatzoglou, I.; Zois, C.E.; Koukourakis, M.I. Evaluation of The Alamarblue Assay for Adherent Cell Irradiation Experiments. Dose-Response 2013, 12, 246–258. [Google Scholar] [CrossRef] [Green Version]
- Weiser, M.R. AJCC 8th ed.; Colorectal Cancer. Ann. Surg. Oncol. 2018, 25, 1454–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenhauer, E.A.; Therasse, P.; Bogaerts, J.; Schwartz, L.H.; Sargent, D.; Ford, R.; Dancey, J.; Arbuck, S.; Gwyther, S.; Mooney, M.; et al. New response evaluation criteria in solid tumours: Revised RECIST guideline (version 1.1). Eur. J. Cancer 2009, 45, 228–247. [Google Scholar] [CrossRef]
- Leighton, X.; Bera, A.; Eidelman, O.; Bubendorf, L.; Zellweger, T.; Banerjee, J.; Gelmann, E.P.; Pollard, H.B.; Srivastava, M. Tissue microarray analysis delineate potential prognostic role of Annexin A7 in prostate cancer progression. PLoS ONE 2018, 13, e0205837. [Google Scholar] [CrossRef] [Green Version]
- Altman, D.G.; McShane, L.M.; Sauerbrei, W.; Taube, S.E. Reporting Recommendations for Tumor Marker Prognostic Studies (REMARK): Explanation and Elaboration. PLoS Med. 2012, 9, e1001216. [Google Scholar] [CrossRef] [Green Version]
- Numata, M.; Morinaga, S.; Watanabe, T.; Tamagawa, H.; Yamamoto, N.; Shiozawa, M.; Nakamura, Y.; Kameda, Y.; Okawa, S.; Rino, Y.; et al. The clinical significance of SWI/SNF complex in pancreatic cancer. Int. J. Oncol. 2012, 42, 403–410. [Google Scholar] [CrossRef]
- Carbone, C.; Piro, G.; Simionato, F.; Ligorio, F.; Cremolini, C.; Loupakis, F.; Alì, G.; Rossini, D.; Merz, V.; Santoro, R.; et al. Homeobox B9 Mediates Resistance to Anti-VEGF Therapy in Colorectal Cancer Patients. Clin. Cancer Res. 2017, 23, 4312–4322. [Google Scholar] [CrossRef] [Green Version]
- Pentheroudakis, G.; Kalogeras, K.T.; Wirtz, R.M.; Grimani, I.; Zografos, G.; Gogas, H.; Stropp, U.; Pectasides, D.; Skarlos, D.; Hennig, G.; et al. Gene expression of estrogen receptor, progesterone receptor and microtubule-associated protein Tau in high-risk early breast cancer: A quest for molecular predictors of treatment benefit in the context of a Hellenic Cooperative Oncology Group trial. Breast Cancer Res. Treat. 2008, 116, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Pallant, J.; Pallant, J. Logistic Regression. In SPSS Survival Manual; Open University Press: Berkshire, UK, 2020; p. 176. [Google Scholar]
- Liao, W.; Overman, M.J.; Boutin, A.T.; Shang, X.; Zhao, D.; Dey, P.; Li, J.; Wang, G.; Lan, Z.; Li, J.; et al. KRAS-IRF2 Axis Drives Immune Suppression and Immune Therapy Resistance in Colorectal Cancer. Cancer Cell 2019, 35, 559–572.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margonis, G.A.; Buettner, S.; Andreatos, N.; Kim, Y.; Wagner, D.; Sasaki, K.; Beer, A.; Schwarz, C.; Løes, I.M.; Smolle, M.; et al. Association of BRAF Mutations With Survival and Recurrence in Surgically Treated Patients With Metastatic Colorectal Liver Cancer. JAMA Surg. 2018, 153, e180996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Wang, T.; Xu, W.; Wang, P.; Wan, J.; Wang, Y.; Zhan, J.; Zhang, H. HOXB9 acetylation at K27 is responsible for its suppression of colon cancer progression. Cancer Lett. 2018, 426, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Margonis, G.A.; Sergentanis, T.N.; Ntanasis-Stathopoulos, I.; Andreatos, N.; Tzanninis, I.G.; Sasaki, K.; Psaltopoulou, T.; Wang, J.; Buettner, S.; He, J.; et al. Impact of Surgical Margin Width on Recurrence and Overall Survival Following R0 Hepatic Resection of Colorectal Metastases: A Systematic Review and Meta-analysis. Ann. Surg. 2018, 267, 1047–1055. [Google Scholar] [CrossRef]
- Yu, M.; Zhan, J.; Zhang, H. HOX family transcription factors: Related signaling pathways and post-translational modifications in cancer. Cell. Signal. 2020, 66, 109469. [Google Scholar] [CrossRef]
- Wan, J.; Xu, W.; Zhan, J.; Ma, J.; Li, X.; Xie, Y.; Wang, J.; Zhu, W.-G.; Luo, J.; Zhang, H. PCAF-mediated acetylation of transcriptional factor HOXB9 suppresses lung adenocarcinoma progression by targeting oncogenic protein JMJD. Nucleic Acids Res. 2016, 44, 10662–10675. [Google Scholar] [CrossRef] [Green Version]
- Brotto, D.B.; Siena, Á.D.D.; de Barros, I.I.; Carvalho, S.; Muys, B.; Goedert, L.; Cardoso, C.; Plaça, J.R.; Ramão, A.; Squire, J.A.; et al. Contributions of HOX genes to cancer hallmarks: Enrichment pathway analysis and review. Tumor Biol. 2020, 42, 1010428320918050. [Google Scholar] [CrossRef]
- Loh, C.-Y.; Chai, J.Y.; Tang, T.F.; Wong, W.F.; Sethi, G.; Shanmugam, M.K.; Chong, P.P.; Looi, C.Y. The E-Cadherin and N-Cadherin Switch in Epithelial-to-Mesenchymal Transition: Signaling, Therapeutic Implications, and Challenges. Cells 2019, 8, 1118. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Definition |
|---|---|
| Age (years) | [Date of Operation–Date of Birth] |
| T | T1–T4, Tumour depth as per American Joint Committee on Cancer (AJCC) 8th edition |
| N | N0, N1, N2, Lymph nodal invasion as per AJCC 8th edition |
| M | M0: No metastatic disease at the time of diagnosis of CRC, (liver metastases were developed later: metachronous) M1: Liver metastatic disease present at the time of diagnosis of colorectal cancer (synchronous) |
| Stage | I–IV, as per AJCC 8th edition |
| Grade | 1: Low differentiation of CRC cells 2: Moderate differentiation of CRC cells 3: High differentiation of CRC cells |
| Primary Tumour Location | Right site: CRC located from the caecum to the transverse colon up to the splenic flexure Left site: CRC located from the splenic flexure to the rectum |
| CRLM location | Unilobar: metastases present at either the left or right liver lobe Bilobar: metastases present at both liver lobes |
| Size of CLRM | Size of largest metastatic deposit measured at histopathological examination (measured in cm) |
| Number of CRLM | Number of metastatic deposits mentioned at histopathology report |
| CEA | CEA level measured at the time of the diagnosis of metastatic liver disease (ng/mL) |
| Response to neoadjuvant chemotherapy | Yes: Patient demonstrating either complete or partial response to chemo on CT according to Response evaluation criteria in solid tumours (RECIST) criteria No: Patient demonstrating either stable disease or disease progression on CT according to RECIST criteria |
| Resection | R0: resection margin ≥1 mm R1: resection margin <1 mm |
| Local Recurrence | Patient demonstrating new intrahepatic disease after first liver resection |
| Overall Survival | Date of death or the date of status checked in the NHS Spine (28 October 2020) minus the date of discharge. |
| Total (n = 96) | Neg: <10 (n = 46) | Low: [10–50] (n = 39) | High: ≥50 (n = 11) | p-Value * | |
|---|---|---|---|---|---|
| Age (mean, SD), [range] | 66 (33), [32–81] | 68 (11), [32–89] | 64 (11), [35–81] | 66 (10), [52–82] | 0.187 |
| Gender, n (%) | |||||
| Male | 63 (67%) | 28 (61%) | 25 (64%) | 10 (91%) | 0.164 |
| Female | 33 (33%) | 18 (39%) | 14 (36%) | 1 (9%) | |
| Deceased | 74 (77%) | 40 (87%) | 25 (64%) | 9 (82%) | 0.195 |
| Primary CRC characteristics | |||||
| Tumour Location, n (%) | |||||
| Right colon | 15 (16%) | 9 (20%) | 5 (13%) | 1 (9%) | 0.402 |
| Left colon | 81 (84%) | 37 (80%) | 34 (87%) | 10 (91%) | |
| Tumour Depth, n (%) | |||||
| T1/2 | 18 (19%) | 8 (17%) | 9 (23%) | 1 (9%) | 0.546 |
| T3/4 | 78 (81%) | 38 (83%) | 30 (77%) | 10 (91%) | |
| Lymph node status, n (%) | |||||
| Negative | 40 (42%) | 19 (41%) | 20 (51%) | 1 (9%) | 0.035 |
| Positive | 56 (58%) | 28 (59%) | 18 (49%) | 10 (91%) | |
| Metastases, n (%) | |||||
| M0 | 60 (63%) | 28 (61%) | 23 (59%) | 9 (82%) | 0.366 |
| M1 | 36 (37%) | 18 (39%) | 16 (41%) | 2 (18%) | |
| Stage, n (%) | |||||
| I/II | 17 (18%) | 8 (17%) | 8 (21%) | 1 (9%) | 0.680 |
| III/IV | 79 (82%) | 38 (83%) | 31 (79%) | 10 (91%) | |
| Grade, n (%) | |||||
| Well/Moderate | 70 (73%) | 36 (84%) | 27 (82%) | 7 (88%) | 0.923 |
| Poor | 14 (15%) | 7 (16%) | 6 (18%) | 1 (12%) | |
| CRLM characteristics | |||||
| CRLM Location, n (%) | |||||
| Unilobar | 65 (68%) | 31 (67%) | 25 (64%) | 9 (82%) | 0.537 |
| Bilobar | 31 (35%) | 15 (33%) | 14 (36%) | 2 (18%) | |
| Number of CRLM, n (%) | |||||
| <4 | 77 (80%) | 37 (80%) | 31 (80%) | 9 (82%) | 0.985 |
| ≥4 | 19 (20%) | 9 (20%) | 8 (20%) | 2 (8%) | |
| Size of CRLM (cm), n (%) | |||||
| <5 | 77 (80%) | 37 (80%) | 30 (77%) | 10 (91%) | 0.589 |
| ≥5 | 19 (20%) | 9 (20%) | 9 (23%) | 1 (9%) | |
| CEA (ng/mL), n (%) | |||||
| <20 | 33 (34%) | 20 (77%) | 12 (100%) | 1 (50%) | 0.387 |
| ≥20 | 7 (7%) | 6 (23%) | 0 (0%) | 1 (50%) | |
| Neoadjuvant Chemo, n (%) | |||||
| Yes | 74 (77%) | 35 (76%) | 30 (77%) | 9 (82%) | 0.919 |
| No | 22 (23%) | 11 (24%) | 9 (23%) | 2 (18%) | |
| Local Recurrence, n (%) | |||||
| Yes | 31 (32%) | 14 (30%) | 14 (36%) | 3 (27%) | 0.865 |
| No | 61 (64%) | 28 (70%) | 25 (44%) | 8 (73%) | |
| Variables | Univariable | Multivariable (1) | Multivariable (2) | Multivariable (3) |
|---|---|---|---|---|
| HR (95% CI) p-Value | HR (95% CI) p-Value | HR (95% CI) p-Value | HR (95% CI) p-Value | |
| Age | 1.02 (0.10–1.04) p = 0.121 | 1.04 (1.00–1.08) p = 0.048 | 1.02 (0.98–1.07) p = 0.333 | |
| Gender (Male) | 1.29 (0.79–2.09) p = 0.303 | |||
| Local Recurrence * | 2.29 (1.40–3.56) p = 0.001 | 4.28 (1.88–9.72) p = 0.001 | 5.73 (2.33–14.08) p < 0.001 | 5.83 (2.11–16.11) p = 0.001 |
| HOXB9 staining (2+) | 1.18 (0.58–2.43) p = 0.648 | |||
| HOXB9 H-Score (High) | 2.13 (0.98–4.63) p = 0.056 | 3.82 (1.59–9.19) p = 0.003 | 4.15 (1.71–10.06) p = 0.002 | 3.79 (1.20–11.98) p = 0.023 |
| Tumour Location * (left) | 0.48 (0.26–0.87) p = 0.017 | 0.39 (0.13–1.13) p = 0.083 | 0.38 (0.13–1.10) p = 0.074 | |
| Number of CRLM * (≥4) | 1.78 (1.03–3.08) p = 0.040 | 1.25 (0.45–3.45) p = 0.665 | 1.41 (0.54–3.71) p = 0.489 | 1.83 (0.58–5.74) p = 0.302 |
| Size of CRLM *(≥5 cm) | 1.87 (1.08–3.25) p = 0.027 | 2.27 (0.88–5.88) p = 0.091 | 2.76 (1.06–7.20) p = 0.038 | 4.44 (1.11–17.75) p = 0.035 |
| T3/4 | 1.34 (0.64–2.81) p = 0.438 | |||
| N1/2 | 1.41 (0.87–2.29) p = 0.168 | 1.04 (0.33–3.28) p = 0.946 | ||
| M1 | 0.99 (0.51–1.90) p = 0.970 | |||
| Stage (III/IV) | 1.23 (0.64–1.97) p = 0.535 | |||
| Grade 2/3 | 1.18 (0.71–1.97) p = 0.518 | |||
| CRLM Location (bilobar) | 1.26 (0.78–2.02) p = 0.342 | 0.42 (0.12–1.46) p = 0.170 | ||
| CEA(≥20 ng/mL) | 1.54 (0.79–3.01) p = 0.207 | |||
| R1 resection | 1.09 (0.51–2.35) p = 0.827 | |||
| Neoadjuvant Chemotherapy | 1.26 (0.72–2.23) p = 0.422 | |||
| Response to Chemotherapy | 0.83 (0.42–1.66) p = 0.598 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martinou, E.; Moller-Levet, C.; Karamanis, D.; Bagwan, I.; Angelidi, A.M. HOXB9 Overexpression Promotes Colorectal Cancer Progression and Is Associated with Worse Survival in Liver Resection Patients for Colorectal Liver Metastases. Int. J. Mol. Sci. 2022, 23, 2281. https://doi.org/10.3390/ijms23042281
Martinou E, Moller-Levet C, Karamanis D, Bagwan I, Angelidi AM. HOXB9 Overexpression Promotes Colorectal Cancer Progression and Is Associated with Worse Survival in Liver Resection Patients for Colorectal Liver Metastases. International Journal of Molecular Sciences. 2022; 23(4):2281. https://doi.org/10.3390/ijms23042281
Chicago/Turabian StyleMartinou, Eirini, Carla Moller-Levet, Dimitrios Karamanis, Izhar Bagwan, and Angeliki M. Angelidi. 2022. "HOXB9 Overexpression Promotes Colorectal Cancer Progression and Is Associated with Worse Survival in Liver Resection Patients for Colorectal Liver Metastases" International Journal of Molecular Sciences 23, no. 4: 2281. https://doi.org/10.3390/ijms23042281
APA StyleMartinou, E., Moller-Levet, C., Karamanis, D., Bagwan, I., & Angelidi, A. M. (2022). HOXB9 Overexpression Promotes Colorectal Cancer Progression and Is Associated with Worse Survival in Liver Resection Patients for Colorectal Liver Metastases. International Journal of Molecular Sciences, 23(4), 2281. https://doi.org/10.3390/ijms23042281