
Systematic Comparison of Beetle Luciferase-Luciferin Pairs as Sources of Near-Infrared Light for In Vitro and In Vivo Applications


Abstract
:1. Introduction
2. Results and Discussion
2.1. Luciferin Analog Synthesis
2.2. Spectral Characterization of Substrates
2.3. Characterization of BL Properties of Luc/Substrate Analog Pairs
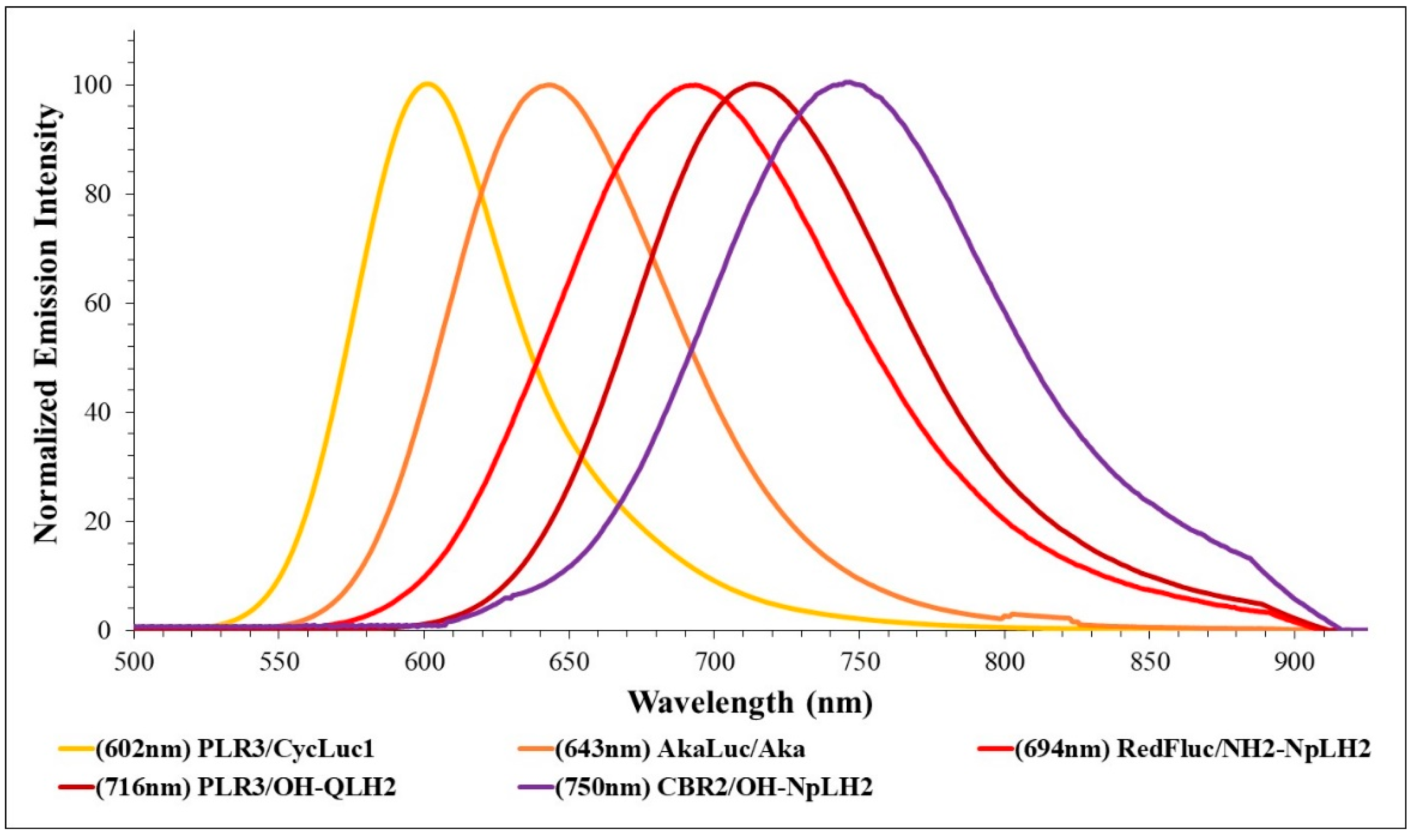
2.3.1. Systematic Study
2.3.2. BL Properties of Luc2 with LH2 and Substrate Analogs In Vitro
2.3.3. BL Properties of LH2 with Lucs In Vitro
2.3.4. Enhanced In Vitro nIR Sources for Biosensor and Biomarker Applications
2.3.5. BL Properties of Luc2 with LH2 and Substrate Analogs in Live Cells
2.3.6. BL Properties of LH2 with Lucs in Live Cells
2.3.7. BL Properties of Optimized and Novel Luc/Substrate Combinations in Live Cells
2.3.8. Enhanced nIR Sources for BLI Applications
3. Materials and Methods
3.1. Materials
3.2. General Methods
3.3. Bioluminescence Emission Spectra
3.4. In Vitro Specific Activities
3.5. In Vitro Bioluminescence Wavelength Emission Distribution
3.6. Cell Culture and Transfection
3.7. Live Cell Imaging in Transfected HEK293T Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mofford, D.M.; Reddy, G.R.; Miller, S.C. Latent luciferase activity in the fruit fly revealed by a synthetic luciferin. Proc. Natl. Acad. Sci. USA 2014, 111, 4443–4448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branchini, B.R.; Behney, C.E.; Southworth, T.L.; Fontaine, D.M.; Gulick, A.M.; Vinyard, D.J.; Brudvig, G.W. Experimental Support for a Single Electron-Transfer Oxidation Mechanism in Firefly Bioluminescence. J. Am. Chem. Soc. 2015, 137, 7592–7595. [Google Scholar] [CrossRef] [PubMed]
- Fetzner, S.; Steiner, R.A. Cofactor-independent oxidases and oxygenases. Appl. Microbiol. Biotechnol. 2010, 86, 791–804. [Google Scholar] [CrossRef] [PubMed]
- Viviani, V.R. The origin, diversity, and structure function relationships of insect luciferases. Cell. Mol. Life Sci. 2002, 59, 1833–1850. [Google Scholar] [CrossRef]
- Branchini, B.R.; Ablamsky, D.M.; Davis, A.L.; Southworth, T.L.; Butler, B.; Fan, F.; Jathoul, A.P.; Pule, M.A. Red-emitting luciferases for bioluminescence reporter and imaging applications. Anal. Biochem. 2010, 396, 290–297. [Google Scholar] [CrossRef]
- Viviani, V.R.; Amaral, D.; Prado, R.; Arnoldi, F.G.C. A new blue-shifted luciferase from the Brazilian Amydetes fanestratus (Coleoptera: Lampyridae) firefly: Molecular evolution and structural/functional properties. Photochem. Photobiol. Sci. 2011, 10, 1879–1886. [Google Scholar] [CrossRef]
- Nishiguchi, T.; Yamada, T.; Nasu, Y.; Ito, M.; Yoshimura, H.; Ozawa, T. Development of red-shifted mutants derived from luciferase of Brazilian click beetle Pyrearinus termitilluminans. J. Biomed. Opt. 2015, 20, 101205. [Google Scholar] [CrossRef]
- Naumov, P.; Ozawa, Y.; Ohkubo, K.; Fukuzumi, S. Structure and Spectroscopy of Oxyluciferin, the Light Emitter of the Firefly Bioluminescence. J. Am. Chem. Soc. 2009, 131, 11590–11605. [Google Scholar] [CrossRef]
- Woodroofe, C.C.; Meisenheimer, P.L.; Klaubert, D.H.; Kovic, Y.; Rosenberg, J.C.; Behney, C.E.; Southworth, T.L.; Branchini, B.R. Novel Heterocyclic Analogues of Firefly Luciferin. Biochemistry 2012, 51, 9807–9813. [Google Scholar] [CrossRef]
- Iwano, S.; Obata, R.; Miura, C.; Kiyama, M.; Hama, K.; Nakamura, M.; Amano, Y.; Kojima, S.; Hirano, T.; Maki, S.; et al. Development of simple firefly luciferin analogs emitting blue, green, red, and near-infrared biological window light. Tetrahedron 2013, 69, 3847–3856. [Google Scholar] [CrossRef]
- Miura, C.; Kiyama, M.; Iwano, S.; Ito, K.; Obata, R.; Hirano, T.; Maki, S.; Niwa, H. Synthesis and luminescence properties of biphenyl-type firefly luciferin analogs with a new, near-infrared light-emitting bioluminophore. Tetrahedron 2013, 69, 9726–9734. [Google Scholar] [CrossRef]
- Jathoul, A.P.; Grounds, H.; Anderson, J.C.; Pule, M.A. A Dual-Color Far-Red to Near-Infrared Firefly Luciferin Analogue Designed for Multiparametric Bioluminescence Imaging. Angew. Chem. Int. Ed. 2014, 53, 13059–13063. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.C.; Grounds, H.; Jathoul, A.P.; Murray, J.A.H.; Pacman, S.J.; Tisi, L. Convergent synthesis and optical properties of near-infrared emitting bioluminescent infra-luciferins. Rsc Adv. 2017, 7, 3975–3982. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.P.; Woodroofe, C.C.; Wood, M.G.; Que, I.; Van’T Root, M.; Ridwan, Y.; Shi, C.; Kirkland, T.A.; Encell, L.P.; Wood, K.V.; et al. Click beetle luciferase mutant and near infrared naphthyl-luciferins for improved bioluminescence imaging. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Iwano, S.; Sugiyama, M.; Hama, H.; Watakabe, A.; Hasegawa, N.; Kuchimaru, T.; Tanaka, K.Z.; Takahashi, M.; Ishida, Y.; Hata, J.; et al. Single-cell bioluminescence imaging of deep tissue in freely moving animals. Science 2018, 359, 935–939. [Google Scholar] [CrossRef] [Green Version]
- Jathoul, A.; Law, E.; Gandelman, O.A.; Pule, M.; Tisi, L.; Murray, J. Development of a pH-Tolerant Thermostable Photinus pyralis Luciferase for Brighter In Vivo Imaging; InTech Open: London, UK, 2012. [Google Scholar]
- Stowe, C.L.; Burley, T.A.; Allan, H.; Vinci, M.; Kramer-Marek, G.; Ciobota, D.M.; Parkinson, G.N.; Southworth, T.L.; Agliardi, G.; Hotblack, A.; et al. Near-infrared dual bioluminescence imaging in mouse models of cancer using infraluciferin. ELife 2019, 8, e45801. [Google Scholar] [CrossRef]
- Steinhardt, R.C.; Rathbun, C.M.; Krull, B.T.; Yu, J.M.; Yang, Y.H.; Nguyen, B.D.; Kwon, J.; McCutcheon, D.C.; Jones, K.A.; Furche, F.; et al. Brominated Luciferins Are Versatile Bioluminescent Probes. ChemBioChem 2017, 18, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.D.; Warner, E.A.; Morrissey, C.E.; Fick, C.W.; Wu, T.S.; Ornelas, M.Y.; Ochoa, G.V.; Zhang, B.S.; Rathbun, C.M.; Porterfield, W.B.; et al. Statistical Coupling Analysis-Guided Library Design for the Discovery of Mutant Luciferases. Biochemistry 2018, 57, 663–671. [Google Scholar] [CrossRef]
- Adams, S.T.; Miller, S.C. Beyond D-luciferin: Expanding the scope of bioluminescence imaging in vivo. Curr. Opin. Chem. Biol. 2014, 21, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Williams, S.J.; Prescher, J.A. Building Biological Flashlights: Orthogonal Luciferases and Luciferins for In Vivo Imaging. Acc. Chem. Res. 2019, 52, 3039–3050. [Google Scholar] [CrossRef]
- Liu, S.; Su, Y.; Lin, M.Z.; Ronald, J.A. Brightening up Biology: Advances in Luciferase Systems for in Vivo Imaging. ACS Chem. Biol. 2021, 16, 2707–2718. [Google Scholar] [CrossRef] [PubMed]
- Contag, C.H.; Bachmann, M.H. Advances in vivo bioluminescence imaging of gene expression. Annu. Rev. Biomed. Eng. 2002, 4, 235–260. [Google Scholar] [CrossRef] [PubMed]
- Nasu, Y.; Campbell, R.E. Unnaturally aglow with a bright inner light. Science 2018, 359, 868–869. [Google Scholar] [CrossRef] [PubMed]
- Abe, M.; Nishihara, R.; Ikeda, Y.; Nakajima, T.; Sato, M.; Iwasawa, N.; Nishiyama, S.; Paulmurugan, R.; Citterio, D.; Kim, S.B.; et al. Near-Infrared Bioluminescence Imaging with a through-Bond Energy Transfer Cassette. ChemBioChem 2019, 20, 1919–1923. [Google Scholar] [CrossRef] [PubMed]
- Weissleder, R.; Ntziachristos, V. Shedding light onto live molecular targets. Nat. Med. 2003, 9, 123–128. [Google Scholar] [CrossRef]
- Saito, R.; Kuchimaru, T.; Higashi, S.; Lu, S.W.; Kiyama, M.; Iwano, S.; Obata, R.; Hirano, T.; Kizaka-Kondoh, S.; Maki, S.A. Synthesis and luminescence properties of near-infrared N-heterocyclic luciferin analogues for in vivo optical imaging. Bull. Chem. Soc. Jpn. 2019, 92, 608–618. [Google Scholar] [CrossRef]
- Branchini, B.R.; Ablamsky, D.M.; Rosenberg, J.C. Chemically Modified Firefly Luciferase Is an Efficient Source of Near-Infrared Light. Bioconjugate Chem. 2010, 21, 2023–2030. [Google Scholar] [CrossRef]
- Branchini, B.R.; Southworth, T.L.; Fontaine, D.M.; Kohrt, D.; Florentine, C.M.; Grossel, M.J. A Firefly Luciferase Dual Color Bioluminescence Reporter Assay Using Two Substrates to Simultaneously Monitor Two Gene Expression Events. Sci. Rep. 2018, 8, 5990. [Google Scholar]
- Rathbun, C.M.; Porterfield, W.B.; Jones, K.A.; Sagoe, M.J.; Reyes, M.R.; Hua, C.T.; Prescher, J.A. Parallel Screening for Rapid Identification of Orthogonal Bioluminescent Tools. Acs Cent. Sci. 2017, 3, 1254–1261. [Google Scholar] [CrossRef]
- Branchini, B.R.; Southworth, T.L.; Fontaine, D.M.; Kohrt, D.; Welcome, F.S.; Florentine, C.M.; Henricks, E.R.; DeBartolo, D.B.; Michelini, E.; Cevenini, L.; et al. Red-emitting chimeric firefly luciferase for in vivo imaging in low ATP cellular environments. Anal. Biochem. 2017, 534, 36–39. [Google Scholar] [CrossRef] [Green Version]
- Shinde, R.; Perkins, J.; Contag, C.H. Luciferin derivatives for enhanced in vitro and in vivo bioluminescence assays. Biochemistry 2006, 45, 11103–11112. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.S.; Chaurette, J.P.; Adams, S.T.; Reddy, G.R.; Paley, M.A.; Aronin, N.; Prescher, J.A.; Miller, S.C. A synthetic luciferin improves bioluminescence imaging in live mice. Nat. Methods 2014, 11, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Simonyan, H.; Hurr, C.; Young, C.N. A synthetic luciferin improves in vivo bioluminescence imaging of gene expression in cardiovascular brain regions. Physiol. Genom. 2016, 48, 762–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Z.; Li, G.Y.; Wu, C.F.; Zhang, M.M.; Zhao, Y.; Liang, G.L. Intracellular synthesis of D-aminoluciferin for bioluminescence generation. Chem. Commun. 2017, 53, 3567–3570. [Google Scholar] [CrossRef]
- Abakumov, M.; Kilpeläinen, A.; Petkov, S.; Belikov, S.; Klyachko, N.; Chekhonin, V.; Isaguliants, M. Evaluation of cyclic luciferin as a substrate for luminescence measurements in in vitro and in vivo applications. Biochem. Biophys. Res. Commun. 2019, 513, 535–539. [Google Scholar] [CrossRef]
- Parkins, K.M.; Dubois, V.P.; Hamilton, A.M.; Makela, A.V.; Ronald, J.A.; Foster, P.J. Multimodality cellular and molecular imaging of concomitant tumour enhancement in a syngeneic mouse model of breast cancer metastasis. Sci. Rep. 2018, 8, 8930. [Google Scholar] [CrossRef]
- Zhang, X.; Lin, C.; Chan, W.; Liu, K.; Lu, A.; Lin, G.; Hu, R.; Shi, H.; Zhang, H.; Yang, Z. Dual-Functional Liposomes with Carbonic Anhydrase IX Antibody and BR2 Peptide Modification Effectively Improve Intracellular Delivery of Cantharidin to Treat Orthotopic Hepatocellular Carcinoma Mice. Molecules 2019, 24, 3332. [Google Scholar] [CrossRef] [Green Version]
- Zambito, G.; Gaspar, N.; Ridwan, Y.; Hall, M.P.; Shi, C.; Kirkland, T.A.; Encell, L.P.; Löwik, C.; Mezzanotte, L. Evaluating Brightness and Spectral Properties of Click Beetle and Firefly Luciferases Using Luciferin Analogues: Identification of Preferred Pairings of Luciferase and Substrate for In Vivo Bioluminescence Imaging. Mol. Imaging Biol. 2020, 22, 1523–1531. [Google Scholar] [CrossRef]
- Viviani, V.R.; Bevilaqua, V.R.; de Souza, D.R.; Pelentir, G.F.; Kakiuchi, M.; Hirano, T. A very bright far-red bioluminescence emitting combination based on engineered railroad worm luciferase and 6′-amino-analogs for bioimaging purposes. Int. J. Mol. Sci. 2021, 22, 1–13. [Google Scholar]
- Anderson, J.C.; Chang, C.H.; Jathoul, A.P.; Syed, A.J. Synthesis and bioluminescence of electronically modified and rotationally restricted colour-shifting infraluciferin analogues. Tetrahedron 2019, 75, 347–356. [Google Scholar] [CrossRef]
- Ikeda, Y.; Nomoto, T.; Hiruta, Y.; Nishiyama, N.; Citterio, D. Ring-Fused Firefly Luciferins: Expanded Palette of Near-Infrared Emitting Bioluminescent Substrates. Anal. Chem. 2020, 92, 4235–4243. [Google Scholar] [CrossRef] [PubMed]
- Saito-moriya, R.; Nakayama, J.; Kamiya, G.; Kitada, N.; Obata, R.; Maki, S.A.; Aoyama, H. How to select firefly luciferin analogues for in vivo imaging. Int. J. Mol. Sci. 2021, 22, 1848. [Google Scholar] [CrossRef] [PubMed]
- Branchini, B.R.; Hayward, M.M.; Bamford, S.; Brennan, P.M.; Lajiness, E.J. Naphthylluciferin and Quinolylluciferin-Green and Red-Light Emitting Firefly Luciferin Analogs. Photochem. Photobiol. 1989, 49, 689–695. [Google Scholar] [CrossRef]
- Takakura, H.; Sasakura, K.; Ueno, T.; Urano, Y.; Terai, T.; Hanaoka, K.; Tsuboi, T.; Nagano, T. Development of Luciferin Analogues Bearing an Amino Group and Their Application as BRET Donors. Chem.-Asian J. 2010, 5, 2053–2061. [Google Scholar] [CrossRef]
- McCutcheon, D.C.; Porterfield, W.B.; Prescher, J.A. Rapid and scalable assembly of firefly luciferase substrates. Org. Biomol. Chem. 2015, 13, 2117–2121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mofford, D.M.; Reddy, G.R.; Miller, S.C. Aminoluciferins Extend Firefly Luciferase Bioluminescence into the Near-Infrared and Can Be Preferred Substrates over D-Luciferin. J. Am. Chem. Soc. 2014, 136, 13277–13282. [Google Scholar] [CrossRef] [Green Version]
- Harwood, K.R.; Mofford, D.M.; Reddy, G.R.; Miller, S.C. Identification of Mutant Firefly Luciferases that Efficiently Utilize Aminoluciferins. Chem. Biol. 2011, 18, 1649–1657. [Google Scholar] [CrossRef] [Green Version]
- Ji, X.; Adams, S.T., Jr.; Miller, S.C. Bioluminescence imaging in mice with synthetic luciferin analogues. Methods Enzymol. 2020, 640, 165–183. [Google Scholar]
- Branchini, B.R.; Ablamsky, D.M.; Rosenman, J.M.; Uzasci, L.; Southworth, T.L.; Zimmer, M. Synergistic mutations produce blue-shifted bioluminescence in firefly luciferase. Biochemistry 2007, 46, 13847–13855. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Specific Activity 1 | BL, 23 °C (λmax ± 2 nm) 2 | BL, 37 °C (λmax ± 2 nm) 2 | Live Cell Activity 3 | Km (μM) 4 |
|---|---|---|---|---|---|
| LH2 | 100 ± 4 | 562 (84) | 600, 574 (105) | 100 ± 9.9 | 15 ± 2 |
| NH2-LH2 | 9.1 ± 0.48 | 603 (85) | 602 (80) | 36 ± 3.9 | 5 ± 1.2 |
| CycLuc1 | 8.3 ± 1.9 | 609 (61) | 606 (64) | 47 ± 4.3 | 0.61 ± 0.2 |
| Aka | 1.9 ± 0.01 | 677 (90) | 674 (88) | 10 ± 1.0 | 2.2 ± 0.2 |
| iLH2 | 0.07 ± 0.001 | 709 (96) | 708 (99) | 0.06 ± 0.007 | 0.13 ± 0.05 |
| 4′-BrLuc | 8.3 ± 0.78 | 617 (70) | 618 (70) | 5.7 ± 1.7 | 44 ± 11 |
| NH2-NpLH2 | 0.18 ± 0.003 | 708 (135) | 706 (126) | 0.58 ± 0.06 | 3.3 ± 0.6 |
| OH-NpLH2 | b.d. | b.d. | b.d. | b.d. | b.d. |
| NH2-QLH2 | 0.08 ± 0.001 | 678 (139) | 678 (130) | 0.18 ± 0.01 | 1.5 ± 0.2 |
| OH-QLH2 | 0.03 ± 0.003 | 720 (154) | 719 (133) | 0.03 ± 0.003 | 1.8 ± 0.6 |
| Luc/LH2 | Specific Activity 1 | BL, 23 °C (λmax ± 2 nm) 2 | BL, 37 °C (λmax ± 2 nm) 2 | Live Cell Activity 3 | Km (μM) 4 |
|---|---|---|---|---|---|
| Luc2 | 100 ± 4 | 562 (84) | 600, 574 (105) | 100 ± 9.9 | 15 ± 2 |
| RedFluc | 51 ± 1.5 | 612 (60) | 613 (63) | 27 ± 2.5 | 66 ± 7 |
| Akaluc | 0.1 ± 0.01 | 598 (73) | 597 (73) | 4.2 ± 0.6 | 64 ± 10 |
| Fluc_red | 47 ± 2.6 | 602 (76) | 604 (68) | 272 ± 27 | 11 ± 0.9 |
| Mut51 | 1.9 ± 0.02 | 568 (101) | 612 (86) | 0.9 ± 0.03 | 98 ± 7 |
| CBR2 | 52 ± 0.26 | 621 (65) | 619 (64) | 156 ± 15 | 250 ± 24 |
| PLR3 | 74 ± 8 | 613 (61) | 611 (62) | 149 ± 17 | 15 ± 4 |
| Luc/Substrate | Specific Activity 1 | BL, 23 °C (λmax ± 2 nm) 2 | BL, 37 °C (λmax ± 2 nm) 2 | Live Cell Activity 3 | Km (μM) 4 |
|---|---|---|---|---|---|
| Luc2/LH2 | 100 ± 4 | 562 (84) | 600, 574 (105) | 100 ± 9.9 | 15 ± 2 |
| CBR2/NH2-LH2 | 10 ± 0.24 | 598 (71) | 600 (72) | 168 ± 30 | 9.5 ± 1.3 |
| PLR3/CycLuc1 | 13.2 ± 0.24 | 606 (62) | 602 (65) | 257 ± 13 | 0.45 ± 0.1 |
| Fluc_red/iLH2 | 0.14 ± 0.02 | 707 (93) | 707 (86) | 0.56 ± 0.03 | 0.17 ± 0.05 |
| CBR2/iLH2 | 0.14 ± 0.002 | 730 (89) | 727 (87) | 1.3 ± 0.04 | 0.9 ± 0.08 |
| Akaluc/Aka | 1.6 ± 0.03 | 642 (93) | 643 (89) | 152 ± 18 | 0.3 ± 0.05 |
| CBR2/NH2-NpLH2 | 0.42 ± 0.01 | 721 (120) | 718 (117) | 3.2 ± 0.09 | 0.8 ± 0.1 |
| CBR2/OH-NpLH2 | 0.48 ± 0.01 | 750 (117) | 749 (116) | 2.2 ± 0.13 | 0.93 ± 0.13 |
| PLR3/NH2-QLH2 | 1.23 ± 0.003 | 649 (111) | 649 (113) | 2.9 ± 0.14 | 0.7 ± 0.15 |
| PLR3/OH-QLH2 | 2.8 ± 0.06 | 718 (107) | 716 (107) | 4.1 ± 0.35 | 1.6 ± 0.1 |
| Mut51/4′-BrLuc | 1.9 ± 0.08 | 617 (77) | 618 (76) | 2.1 ± 0.34 | 2.3 ± 0.2 |
| PLR3/4′-BrLuc | 44 ± 2 | 615 (63) | 615 (69) | 68 ± 5.8 | 4.5 ± 0.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Branchini, B.R.; Fontaine, D.M.; Kohrt, D.; Huta, B.P.; Racela, A.R.; Fort, B.R.; Southworth, T.L.; Roda, A. Systematic Comparison of Beetle Luciferase-Luciferin Pairs as Sources of Near-Infrared Light for In Vitro and In Vivo Applications. Int. J. Mol. Sci. 2022, 23, 2451. https://doi.org/10.3390/ijms23052451
Branchini BR, Fontaine DM, Kohrt D, Huta BP, Racela AR, Fort BR, Southworth TL, Roda A. Systematic Comparison of Beetle Luciferase-Luciferin Pairs as Sources of Near-Infrared Light for In Vitro and In Vivo Applications. International Journal of Molecular Sciences. 2022; 23(5):2451. https://doi.org/10.3390/ijms23052451
Chicago/Turabian StyleBranchini, Bruce R., Danielle M. Fontaine, Dawn Kohrt, Brian P. Huta, Allison R. Racela, Benjamin R. Fort, Tara L. Southworth, and Aldo Roda. 2022. "Systematic Comparison of Beetle Luciferase-Luciferin Pairs as Sources of Near-Infrared Light for In Vitro and In Vivo Applications" International Journal of Molecular Sciences 23, no. 5: 2451. https://doi.org/10.3390/ijms23052451