
ICA1L Is Associated with Small Vessel Disease: A Proteome-Wide Association Study in Small Vessel Stroke and Intracerebral Haemorrhage

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
2.1. Stringent Criterion PWAS Analysis
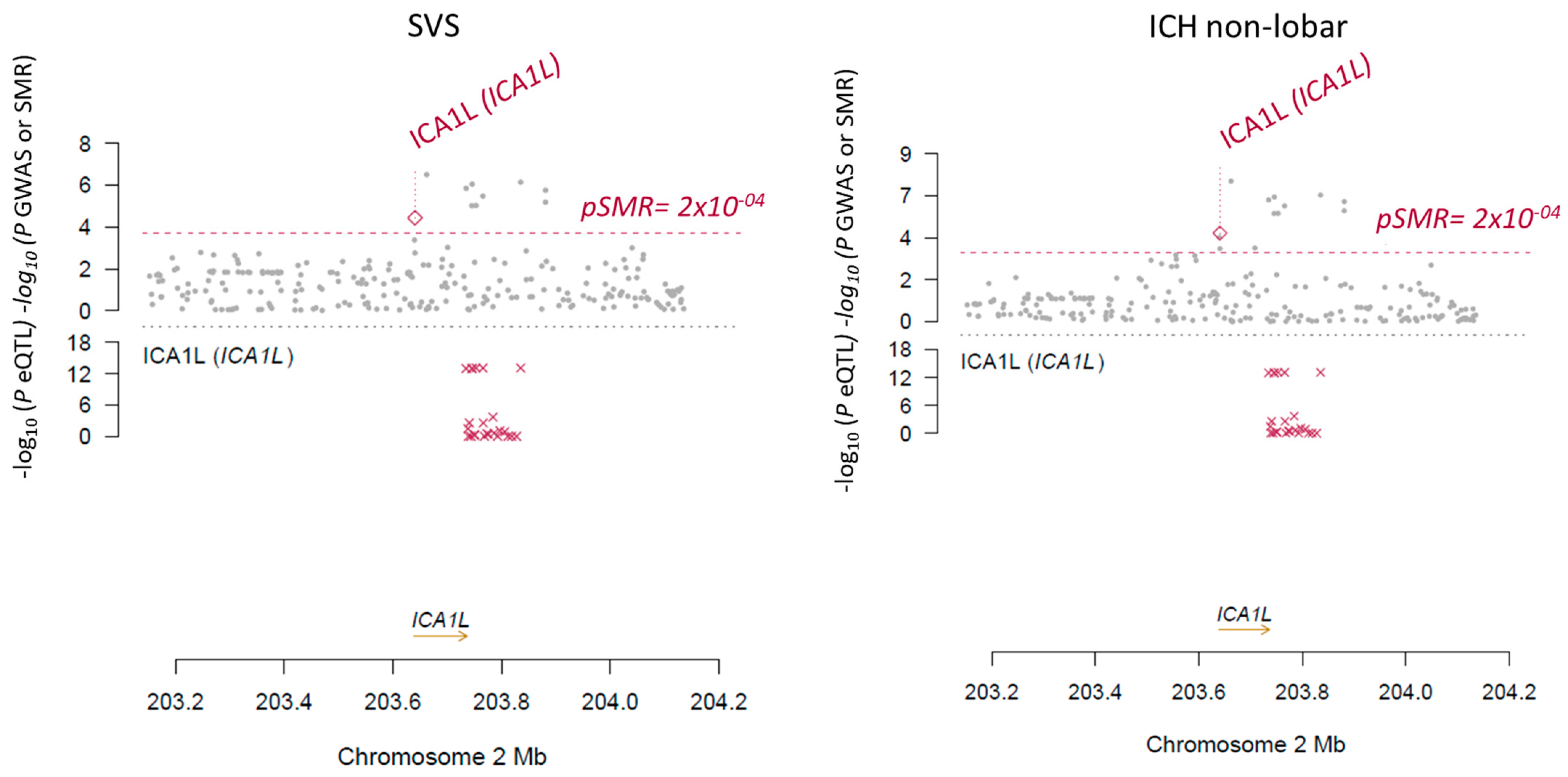
2.2. SMR and Conditional Analysis
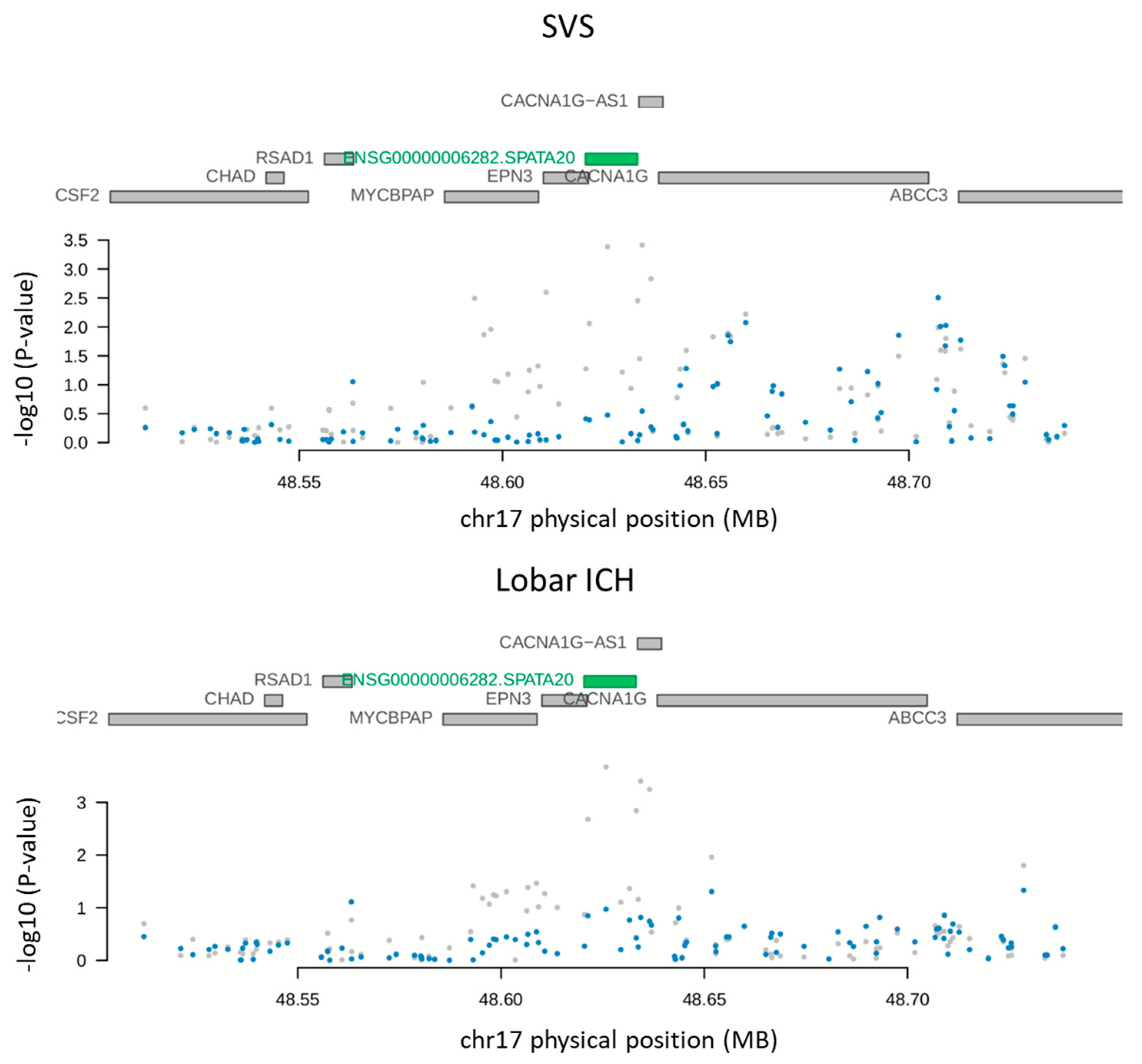
2.3. Open Criterion PWAS Analysis
3. Discussion
4. Materials and Methods
4.1. Proteome-Wide Associations Study (PWAS)
4.2. Colocalization Analysis
4.3. Replication Analysis
4.4. Summary-Based Mendelian Randomization (SMR)
4.5. Conditional Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pantoni, L. Cerebral Small Vessel Disease: From Pathogenesis and Clinical Characteristics to Therapeutic Challenges. Lancet Neurol. 2010, 9, 689–701. [Google Scholar] [CrossRef]
- Uniken Venema, S.M.; Marini, S.; Lena, U.K.; Morotti, A.; Jessel, M.; Moomaw, C.J.; Kourkoulis, C.; Testai, F.D.; Kittner, S.J.; Brouwers, H.B.; et al. Impact of Cerebral Small Vessel Disease on Functional Recovery After Intracerebral Hemorrhage. Stroke 2019, 50, 2722–2728. [Google Scholar] [CrossRef] [PubMed]
- Ter Telgte, A.; van Leijsen, E.M.C.; Wiegertjes, K.; Klijn, C.J.M.; Tuladhar, A.M.; de Leeuw, F.-E. Cerebral Small Vessel Disease: From a Focal to a Global Perspective. Nat. Rev. Neurol. 2018, 14, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.; Marini, S.; Pera, J.; Norrving, B.; Jimenez-Conde, J.; Roquer, J.; Fernandez-Cadenas, I.; Tirschwell, D.L.; Selim, M.; Brown, D.L.; et al. Genome-Wide Association Study of Cerebral Small Vessel Disease Reveals Established and Novel Loci. Brain 2019, 142, 3176–3189. [Google Scholar] [CrossRef] [PubMed]
- Woo, D.; Falcone, G.J.; Devan, W.J.; Brown, W.M.; Biffi, A.; Howard, T.D.; Anderson, C.D.; Brouwers, H.B.; Valant, V.; Battey, T.W.K.; et al. Meta-Analysis of Genome-Wide Association Studies Identifies 1q22 as a Susceptibility Locus for Intracerebral Hemorrhage. Am. J. Hum. Genet. 2014, 94, 511–521. [Google Scholar] [CrossRef] [Green Version]
- Traylor, M.; Malik, R.; Nalls, M.A.; Cotlarciuc, I.; Radmanesh, F.; Thorleifsson, G.; Hanscombe, K.B.; Langefeld, C.; Saleheen, D.; Rost, N.S.; et al. Genetic Variation at 16q24.2 Is Associated with Small Vessel Stroke. Ann. Neurol. 2017, 81, 383–394. [Google Scholar] [CrossRef] [Green Version]
- Qureshi, A.I.; Mendelow, A.D.; Hanley, D.F. Intracerebral Haemorrhage. Lancet 2009, 373, 1632–1644. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Delcourt, C.; Heeley, E.; Arima, H.; Zhang, S.; Al-Shahi Salman, R.; Stapf, C.; Woo, D.; Flaherty, M.L.; Vagal, A.; et al. Significance of Cerebral Small-Vessel Disease in Acute Intracerebral Hemorrhage. Stroke 2016, 47, 701–707. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.J.; Lee, S.-H. Prognostic Impact of Cerebral Small Vessel Disease on Stroke Outcome. J. Stroke 2015, 17, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Regenhardt, R.W.; Das, A.S.; Lo, E.H.; Caplan, L.R. Advances in Lacunar Stroke Pathophysiology: A Review. JAMA Neurol. 2018, 75, 1273–1281. [Google Scholar] [CrossRef]
- Fornage, M.; Debette, S.; Bis, J.C.; Schmidt, H.; Ikram, M.A.; Dufouil, C.; Sigurdsson, S.; Lumley, T.; DeStefano, A.L.; Fazekas, F.; et al. Genome-Wide Association Studies of Cerebral White Matter Lesion Burden: The CHARGE Consortium. Ann. Neurol. 2011, 69, 928–939. [Google Scholar] [CrossRef] [PubMed]
- Haffner, C.; Malik, R.; Dichgans, M. Genetic Factors in Cerebral Small Vessel Disease and Their Impact on Stroke and Dementia. J. Cereb. Blood Flow Metab. 2016, 36, 158–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhaaren, B.F.J.; Debette, S.; Bis, J.C.; Smith, J.A.; Ikram, M.K.; Adams, H.H.; Beecham, A.H.; Rajan, K.B.; Lopez, L.M.; Barral, S.; et al. Multiethnic Genome-Wide Association Study of Cerebral White Matter Hyperintensities on MRI. Circ. Cardiovasc. Genet. 2015, 8, 398–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traylor, M.; Zhang, C.R.; Adib-Samii, P.; Devan, W.J.; Parsons, O.E.; Lanfranconi, S.; Gregory, S.; Cloonan, L.; Falcone, G.J.; Radmanesh, F.; et al. Genome-Wide Meta-Analysis of Cerebral White Matter Hyperintensities in Patients with Stroke. Neurology 2016, 86, 146–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traylor, M.; Tozer, D.J.; Croall, I.D.; Lisiecka-Ford, D.M.; Olorunda, A.O.; Boncoraglio, G.; Dichgans, M.; Lemmens, R.; Rosand, J.; Rost, N.S.; et al. Genetic Variation in PLEKHG1 Is Associated with White Matter Hyperintensities (n = 11,226). Neurology 2019, 92, e749–e757. [Google Scholar] [CrossRef] [Green Version]
- Persyn, E.; Hanscombe, K.B.; Howson, J.M.M.; Lewis, C.M.; Traylor, M.; Markus, H.S. Genome-Wide Association Study of MRI Markers of Cerebral Small Vessel Disease in 42,310 Participants. Nat. Commun. 2020, 11, 2175. [Google Scholar] [CrossRef]
- Sargurupremraj, M.; Suzuki, H.; Jian, X.; Sarnowski, C.; Evans, T.E.; Bis, J.C.; Eiriksdottir, G.; Sakaue, S.; Terzikhan, N.; Habes, M.; et al. Cerebral Small Vessel Disease Genomics and Its Implications across the Lifespan. Nat. Commun. 2020, 11, 6285. [Google Scholar] [CrossRef]
- Rutten-Jacobs, L.C.A.; Tozer, D.J.; Duering, M.; Malik, R.; Dichgans, M.; Markus, H.S.; Traylor, M. Genetic Study of White Matter Integrity in UK Biobank (N = 8448) and the Overlap with Stroke, Depression, and Dementia. Stroke 2018, 49, 1340–1347. [Google Scholar] [CrossRef]
- Armstrong, N.J.; Mather, K.A.; Sargurupremraj, M.; Knol, M.J.; Malik, R.; Satizabal, C.L.; Yanek, L.R.; Wen, W.; Gudnason, V.G.; Dueker, N.D.; et al. Common Genetic Variation Indicates Separate Etiologies for Periventricular and Deep White Matter Hyperintensities. Stroke 2020, 51, 2111–2121. [Google Scholar] [CrossRef]
- Knol, M.J.; Lu, D.; Traylor, M.; Adams, H.H.H.; Romero, J.R.J.; Smith, A.V.; Fornage, M.; Hofer, E.; Liu, J.; Hostettler, I.C.; et al. Association of Common Genetic Variants with Brain Microbleeds: A Genome-Wide Association Study. Neurology 2020, 95, e3331–e3343. [Google Scholar] [CrossRef]
- Li, H.-Q.; Cai, W.-J.; Hou, X.-H.; Cui, M.; Tan, L.; Yu, J.-T.; Dong, Q. Alzheimer’s Disease Neuroimaging Initiative Genome-Wide Association Study of Cerebral Microbleeds on MRI. Neurotox. Res. 2020, 37, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Malik, R.; Chauhan, G.; Traylor, M.; Sargurupremraj, M.; Okada, Y.; Mishra, A.; Rutten-Jacobs, L.; Giese, A.-K.; van der Laan, S.W.; Gretarsdottir, S.; et al. Multiancestry Genome-Wide Association Study of 520,000 Subjects Identifies 32 Loci Associated with Stroke and Stroke Subtypes. Nat. Genet. 2018, 50, 524–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traylor, M.; Persyn, E.; Tomppo, L.; Klasson, S.; Abedi, V.; Bakker, M.K.; Torres, N.; Li, L.; Bell, S.; Rutten-Jacobs, L.; et al. Genetic Basis of Lacunar Stroke: A Pooled Analysis of Individual Patient Data and Genome-Wide Association Studies. Lancet Neurol. 2021, 20, 351–361. [Google Scholar] [CrossRef]
- Brandes, N.; Linial, N.; Linial, M. PWAS: Proteome-Wide Association Study—Linking Genes and Phenotypes by Functional Variation in Proteins. Genome Biol. 2020, 21, 173. [Google Scholar] [CrossRef]
- Ou, Y.-N.; Yang, Y.-X.; Deng, Y.-T.; Zhang, C.; Hu, H.; Wu, B.-S.; Liu, Y.; Wang, Y.-J.; Zhu, Y.; Suckling, J.; et al. Identification of Novel Drug Targets for Alzheimer’s Disease by Integrating Genetics and Proteomes from Brain and Blood. Mol. Psychiatry 2021, 1–9. [Google Scholar] [CrossRef]
- Wingo, T.S.; Liu, Y.; Gerasimov, E.S.; Gockley, J.; Logsdon, B.A.; Duong, D.M.; Dammer, E.B.; Lori, A.; Kim, P.J.; Ressler, K.J.; et al. Brain Proteome-Wide Association Study Implicates Novel Proteins in Depression Pathogenesis. Nat. Neurosci. 2021, 24, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Malik, R.; Dichgans, M. Challenges and Opportunities in Stroke Genetics. Cardiovasc. Res. 2018, 114, 1226–1240. [Google Scholar] [CrossRef]
- Wingo, A.P.; Liu, Y.; Gerasimov, E.S.; Gockley, J.; Logsdon, B.A.; Duong, D.M.; Dammer, E.B.; Robins, C.; Beach, T.G.; Reiman, E.M.; et al. Integrating Human Brain Proteomes with Genome-Wide Association Data Implicates New Proteins in Alzheimer’s Disease Pathogenesis. Nat. Genet. 2021, 53, 143–146. [Google Scholar] [CrossRef]
- Liu, D.-Z.; Sharp, F.R. Excitatory and Mitogenic Signaling in Cell Death, Blood-Brain Barrier Breakdown, and BBB Repair after Intracerebral Hemorrhage. Transl. Stroke Res. 2012, 3, 62–69. [Google Scholar] [CrossRef]
- Lai, T.W.; Zhang, S.; Wang, Y.T. Excitotoxicity and Stroke: Identifying Novel Targets for Neuroprotection. Prog. Neurobiol. 2014, 115, 157–188. [Google Scholar] [CrossRef] [Green Version]
- Chagnot, A.; Barnes, S.R.; Montagne, A. Magnetic Resonance Imaging of Blood-Brain Barrier Permeability in Dementia. Neuroscience 2021, 474, 14–29. [Google Scholar] [CrossRef] [PubMed]
- Kalaria, R.N.; Sepulveda-Falla, D. Cerebral Small Vessel Disease in Sporadic and Familial Alzheimer Disease. Am. J. Pathol. 2021, 191, 1888–1905. [Google Scholar] [CrossRef] [PubMed]
- Bordes, C.; Sargurupremraj, M.; Mishra, A.; Debette, S. Genetics of Common Cerebral Small Vessel Disease. Nat. Rev. Neurol. 2022, 1–18. [Google Scholar] [CrossRef]
- Vuckovic, D.; Bao, E.L.; Akbari, P.; Lareau, C.A.; Mousas, A.; Jiang, T.; Chen, M.-H.; Raffield, L.M.; Tardaguila, M.; Huffman, J.E.; et al. The Polygenic and Monogenic Basis of Blood Traits and Diseases. Cell 2020, 182, 1214–1231.e11. [Google Scholar] [CrossRef] [PubMed]
- Van der Meer, D.; Frei, O.; Kaufmann, T.; Shadrin, A.A.; Devor, A.; Smeland, O.B.; Thompson, W.K.; Fan, C.C.; Holland, D.; Westlye, L.T.; et al. Understanding the Genetic Determinants of the Brain with MOSTest. Nat. Commun. 2020, 11, 3512. [Google Scholar] [CrossRef]
- Gusev, A.; Ko, A.; Shi, H.; Bhatia, G.; Chung, W.; Penninx, B.W.J.H.; Jansen, R.; de Geus, E.J.C.; Boomsma, D.I.; Wright, F.A.; et al. Integrative Approaches for Large-Scale Transcriptome-Wide Association Studies. Nat. Genet. 2016, 48, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Bennett, D.A.; Buchman, A.S.; Boyle, P.A.; Barnes, L.L.; Wilson, R.S.; Schneider, J.A. Religious Orders Study and Rush Memory and Aging Project. J. Alzheimer’s Dis. 2018, 64, S161–S189. [Google Scholar] [CrossRef]
- Beach, T.G.; Adler, C.H.; Sue, L.I.; Serrano, G.; Shill, H.A.; Walker, D.G.; Lue, L.; Roher, A.E.; Dugger, B.N.; Maarouf, C.; et al. Arizona Study of Aging and Neurodegenerative Disorders and Brain and Body Donation Program. Neuropathology 2015, 35, 354–389. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhang, F.; Hu, H.; Bakshi, A.; Robinson, M.R.; Powell, J.E.; Montgomery, G.W.; Goddard, M.E.; Wray, N.R.; Visscher, P.M.; et al. Integration of Summary Data from GWAS and EQTL Studies Predicts Complex Trait Gene Targets. Nat. Genet. 2016, 48, 481–487. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Discovery | Replication | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Trait | Protein | Chr | P0–P1 | pQTL ID | pQTL z-Score | pQTL GWAS z-Score | NSNP | PWAS z-Score | PWAS p-Value | PP4 | q-Value | PWAS z-Score | PWAS p-Value | PP4 | q-Value |
| SVS | ICA1L | 2 | 203640690 - 203736708 | rs7582720 | 7.21 | −4.426 | 81 | −4.43 | 9.60 × 10−6 | 0.90 | 1.42 × 10−2 | −3.55 | 3.91 × 10−4 | 0.99 | 5.08 × 10−3 |
| Non-lobar ICH | −4.801 | −4.80 | 1.58 × 10−6 | 0.93 | 2.33 × 10−3 | −4.07 | 4.76 × 10−5 | 0.99 | 6.19 × 10−4 | ||||||
| Trait | Protein | b_SMR | p_SMR |
|---|---|---|---|
| SVS | ICA1L | −2.78 | 3.66 × 10−5 |
| SVS | SPATA20 | −0.24 | 3.69 × 10−1 |
| ICH | ICA1L | −1.55 | 1.81 × 10−5 |
| Non-lobar ICH | ICA1L | −1.55 | 1.81 × 10−5 |
| Lobar ICH | SPATA20 | −0.05 | 7.98 × 10−1 |
| Discovery | Replication | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Trait | Protein | Chr | pQTL ID | pQTL z-Score | pQTL GWAS z-Score | NSNP | PWAS z-Score | PWAS p-Value | PP4 | q-Value | PWAS Z-Score | PWAS p-Value | PP4 | q-Value |
| SVS | SPATA20 | 17 | rs878619 | −10.89 | −2.917 | 102 | 3.71 | 2.10 × 10−4 | 0.17 | 3.10 × 10−1 | 3.79 | 1.50 × 10−4 | 0.97 * | 1.95 × 10−3 |
| SVS | ALDH2 | 12 | rs4648328 | −7.25 | −3.437 | 61 | 3.58 | 3.38 × 10−4 | 0.29 | 4.99 × 10−1 | 3.38 | 7.28 × 10−4 | 0.27 | 9.46 × 10−3 |
| SVS | EXOC6 | 10 | rs980204 | 4.98 | −2.469 | 210 | −3.58 | 3.41 × 10−4 | 0.19 | 5.03 × 10−1 | 3.37 | 7.36 × 10−1 | - | 1 |
| SVS | OSBPL11 | 3 | rs2922170 | 4.11 | 3577 | 111 | 3.58 | 3.48 × 10−4 | 0.35 | 5.14 × 10−1 | - | - | - | |
| ICH | ICA1L | 2 | rs7582720 | 7.21 | −3.935 | 81 | −3.94 | 8.32 × 10−5 | 0.68 | 1.23 × 10−1 | −3.30 | 9.66 × 10−4 | 0.99 * | 1.25 × 10−2 |
| ICH | NRBF2 | 10 | rs4379723 | −4.19 | −3.96 | 77 | 3.85 | 1.19 × 10−4 | 0.18 | 1.76 × 10−1 | - | - | - | |
| ICH | DLGAP2 | 8 | rs7842425 | 4.32 | −3.742 | 225 | −3.74 | 1.83 × 10−4 | 0.12 | 2.70 × 10−1 | 1.14 | 2.54 × 10−1 | - | 1 |
| ICH | SPATA20 | 17 | rs878619 | −10.89 | −2.89 | 102 | 3.58 | 3.47 × 10−4 | 0.12 | 5.12 × 10−1 | 3.27 | 1.09 × 10−3 | 0.33 | 1.41 × 10−2 |
| ICH | MADD | 11 | rs11570115 | −9.09 | −2.565 | 99 | 3.54 | 4.06 × 10−4 | 0.03 | 5.99 × 10−1 | 1.88 | 6.04 × 10−2 | - | 0.78 |
| Non-lobar ICH | SPATA20 | 17 | rs878619 | −10.89 | −2.672 | 102 | 3.69 | 2.19 × 10−4 | 0.42 | 3.23 × 10−1 | 3.51 | 4.44 × 10−4 | 0.27 | 5.77 × 10−3 |
| Non-lobar ICH | NRBF2 | 10 | rs4379723 | −4.19 | −3.569 | 77 | 3.43 | 6.00× 10−4 | 0.29 | 8.90 × 10−1 | - | - | - | |
| Lobar ICH | MRVI1 | 11 | rs753002 | −5.36 | 2.452 | 211 | −3.69 | 2.20 × 10−4 | 0.02 | 3.25 × 10−1 | −1.87 | 6.22× 10−2 | - | 0.80 |
| Lobar ICH | NRBF2 | 10 | rs4379723 | −4.19 | −3.804 | 77 | 3.68 | 2.33 × 10−4 | 0.11 | 3.44 × 10−1 | - | - | ||
| Lobar ICH | SPATA20 | 17 | rs878619 | −10.89 | −3.186 | 102 | 3.51 | 4.44× 10−4 | 0.28 | 6.55 × 10−1 | 3.40 | 6.79× 10−4 | 0.55 * | 8.82 × 10−3 |
| Lobar ICH | ICA1L | 2 | rs7582720 | 7.21 | 81 | −3.30 | 9.6× 10−4 | 0.2 | 1 | −2.5692 | 1.02 × 10−2 | 0.9 | 0.13 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cullell, N.; Gallego-Fábrega, C.; Cárcel-Márquez, J.; Muiño, E.; Llucià-Carol, L.; Lledós, M.; Martín-Campos, J.M.; Molina, J.; Casas, L.; Almeria, M.; et al. ICA1L Is Associated with Small Vessel Disease: A Proteome-Wide Association Study in Small Vessel Stroke and Intracerebral Haemorrhage. Int. J. Mol. Sci. 2022, 23, 3161. https://doi.org/10.3390/ijms23063161
Cullell N, Gallego-Fábrega C, Cárcel-Márquez J, Muiño E, Llucià-Carol L, Lledós M, Martín-Campos JM, Molina J, Casas L, Almeria M, et al. ICA1L Is Associated with Small Vessel Disease: A Proteome-Wide Association Study in Small Vessel Stroke and Intracerebral Haemorrhage. International Journal of Molecular Sciences. 2022; 23(6):3161. https://doi.org/10.3390/ijms23063161
Chicago/Turabian StyleCullell, Natalia, Cristina Gallego-Fábrega, Jara Cárcel-Márquez, Elena Muiño, Laia Llucià-Carol, Miquel Lledós, Jesús M. Martín-Campos, Jessica Molina, Laura Casas, Marta Almeria, and et al. 2022. "ICA1L Is Associated with Small Vessel Disease: A Proteome-Wide Association Study in Small Vessel Stroke and Intracerebral Haemorrhage" International Journal of Molecular Sciences 23, no. 6: 3161. https://doi.org/10.3390/ijms23063161