
The Potent Phytoestrogen 8-Prenylnaringenin: A Friend or a Foe?


Abstract
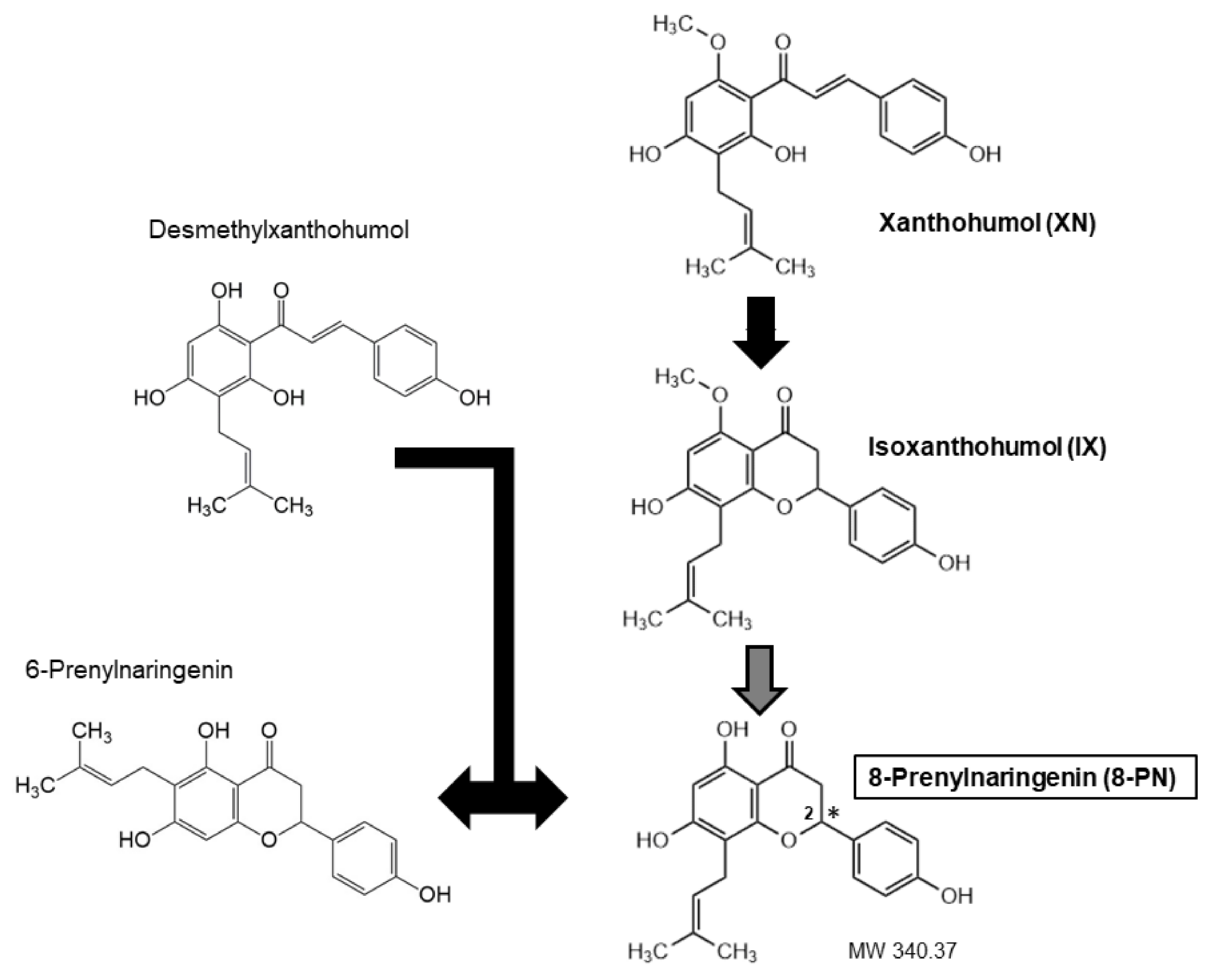
:1. Introduction
2. Occurrence, Sources of, and Exposure to 8-PN
3. Methods Applied in 8-PN Identification and Quantification

{kind=link}
{kind=link}
| Sensitivity | Precision | Accuracy | ||||||
|---|---|---|---|---|---|---|---|---|
| Method 1 | Matrix | LLOQ 2 (ng/mL) | LOD 3 (ng/mL) | CV 4-Intra (%) | CV-Inter (%) | RE 5 (%) | Related Analytes Measured | Reference |
| GC/MS-SIM | Beer, hop pellets | NP | 5 (beer) | NP | NP | ≤65 (beer) | – | [26] |
| LC–ESI–MS | Beer | 2.4 | 0.8 | 2.0 | 8.0 | 8.8 | – | [27] |
| LC–ESI–MS | Serum, urine 6 | S: 50; U: 10 | NP | S: ≤10.9; U: ≤14.9 | S: ≤13.7; U: ≤14.1 | S: 2.6; U: 2.2 | – | [20] |
| LC−ESI-MS/MS | Urine, beer | 5 (urine) | 0.03 (urine) | ≤13.9 (urine) | ≤12.6 (urine) | ≤14.6 (urine) | X, IX | [29] |
| HPLC–MS/MS | Beer, hop extracts, herb teas | NP | NP | 8.8 (beer) | 8.2 (beer) | ≤10 (beer) | XN, IX, 6-PN | [15] |
| UHPLC–MS/MS | Serum | 1 | NP | ≤10.5 | ≤12.1 | ≤4 | XN, IX, 6-PN | [28] |
| UHPLC–MS/MS | Beer, hop pellets | NP | NP | ≤6 | ≤5 | ≤13 | XN, IX | [19] |
| HPLC–APCI–MS | Dietary supplement for breast enhancement | NP | NP | 6 | 2 | 3 | XN, IX, 6-PN, 6,8-diPN | [24] |
| HPLC–APCI–MS | Serum, urine | S: 4.8; U: 1.2 | S: 1.5; U: 0.4 | S: 3.9; U: 6.0 | S: 9.7; U: 14.4 | S: 7.4; U: 10.1 | XN, IX | [30] |
| HPLC–APCI–MS/MS | Beer, hop extracts, herb teas | NP | NP | ≤7.9 | ≤8.2 (beer) | ≤10 | XN, IX, 6-PN | [15] |
| HPLC–APCI–MS | Beer, hop extracts | 20 | 6 | NP | NP | NP | XN, IX | [18] |
| HPLC–UV | Beer, hop extracts | 100 | 30 | NP | NP | NP | XN, IX | [18] |
| HPLC-UV | Beer, hops, hop pellets | 30 | 10 | <0.3 | ≤ 2.0 | ≤4.8 | XN, IX | [32] |
| HPLC-UV/DAD | Hop cultivars and genotypes | 3800 | 1000 | NP | NP | 3.2 | XN, IX, 6-PN | [31] |
| HPLC–UV/DAD (+SS) | Hop extract and capsules | NP | 860 | <4.0 | ≤ 5.0 | ≤8.1 | XN, IX, 6-PN | [25] |
| UHPSFC–UV | Hop dietary supplements, herbal products | 100 | 60 | 0.0 7 | 0.1 7 | ≤7.7 | XN, IX, 6-PN | [23] |
| UHPSFC–MS | Hop dietary supplements, herbal products | 50 | 20 | 0.02 7 | 0.1 7 | NP | XN, IX, 6-PN | [23] |
| RIA (polyclonal) 8 | Beer, urine | NP | 0.3 (urine) | <9 (urine) | <27 (urine) | ≤36 (urine) | – | [33] |
| ELISA (monoclonal) 9 | Serum, urine | 17.1 | 4.4 | S: 2.4; U: 0.7 | S: 6.1; U: 7.2 | S: 5.6; U: 5.1 | X, IX | [34] |
| SIDA-LC-MS/MS | Beer, hop pellets, and tea | 1.3 (beer) | 0.32 (beer) | 2.09 (beer) | 8.2 (beer) | ≤13 (beer) | XN, IX, 6-PN | [16] |
| 1 Method abbreviations: | APCI, atmospheric pressure chemical ionization; DAD, diode-array detection; ELISA, enzyme-linked immunosorbent assay; ESI, electrospray ionization; GC, gas chromatography; | |||||||
| HPLC, high-performance liquid chromatography; LC, liquid chromatography; MS, mass spectrometry; RIA, radioimmunoassay; SIDA, stable isotope dilution analysis; | ||||||||
| UHPSFC, ultrahigh-performance supercritical fluid chromatography; UV, ultraviolet light | ||||||||
| 2 Lower limit of quantification | ||||||||
| 3 Limit of detection | ||||||||
| 4 Coefficient of variation | ||||||||
| 5 Relative error | ||||||||
| 6 For both enantiomers | ||||||||
| 7 Based on peak area | ||||||||
| 8 Both enantiomers detected; cross-reactivity with XN, IX and 6-PN ≤ 0.15% | ||||||||
| 9 Both enantiomers detected; cross-reactivity with XN and IX < 0.01%, with 6-PN < 0.03% | ||||||||
4. Pharmacokinetics of 8-PN
5. Estrogenic Activity of 8-PN
5.1. In Vitro Studies
5.2. In Vivo Studies
| 1. In Vitro | ||||||||
|---|---|---|---|---|---|---|---|---|
| Test Assay | Type of 8-PN | Variable Measured | 8-PN | E2 | Coumestrol | Genistein | Daidzein | Reference |
| Binding to ER in rat uterine cytosol | Racemic | Relative affinity | 0.023 | 1 | 0.008 | 0.003 | [7] | |
| Stimulation of alkaline phosphatase | Racemic | EC50 (nM) | 4 | 0.8 | 30 | 200 | 1500 | [7] |
| activity in Ishikawa cells | ||||||||
| ERE-reporter gene expression in yeast | Racemic | EC50 (nM) | 40 | 0.3 | 70 | 1200 | 2200 | [7] |
| cells transfected with the human ERα | ||||||||
| Human ERα binding in vitro | (R)-8-PN | Relative affinity | ~0.01 | 1 | [5] | |||
| Human ERα binding in vitro | (S)-8-PN | Relative affinity | ~0.01 | 1 | [5] | |||
| Human ERα binding in vitro | (R)-8-PN | Relative affinity | ~0.01 | 1 | [5] | |||
| Human ERα binding in vitro | (S)-8-PN | Relative affinity | ~0.01 | 1 | [5] | |||
| Inhibition of E2 binding to ER in MCF-7 cells | Racemic | Molar excess over E2 | 45 | 35 | 1000 | >104 | [62] | |
| ERE-CAT induction in MCF-7 cells | Racemic | Relative potency | 100 | 1 | 330 | 250 | 3300 | [62] |
| Proliferation of MCF-7 cells after 7 days | Racemic | Relative potency | 3.3 | 1 | 500 | 500 | 5000 | [62] |
| ERE-reporter gene expression in yeast | Racemic | EC50 (nM) | 0.02 | 0.005 | 40 | [85] | ||
| cells transfected with the human ERα | ||||||||
| Proliferation of MCF-7 cells after 24 h | Racemic | EC50 (nM) | 5 | 0.2 | 830 | [85] | ||
| ERE-reporter gene expression in yeast | Racemic | EC50 (nM) | 100 | 0.8 | 140 | 2000 | No resp. | [67] |
| cells transfected with the human ERα | ||||||||
| ERE-reporter gene expression in yeast | Racemic | EC50 (nM) | 33 | 0.2 | 3 | 8 | 800 | [67] |
| cells transfected with the human ERβ | ||||||||
| Human ERα binding in vitro | Racemic | IC50 (nM) | 510 | 20 | 300 | 17,000 | [61] | |
| Human ERβ binding in vitro | Racemic | IC50 (nM) | 1700 | 15 | 20 | 1200 | [61] | |
| Human ERα binding in vitro | Racemic | IC50 (nM) | 59 | 11 | [66] | |||
| Human ERβ binding in vitro | Racemic | IC50 (nM) | 65 | 4.2 | [66] | |||
| ERE-reporter gene expression in yeast | Racemic | EC50 (nM) | 130 | 0.8 | 9300 | [64] | ||
| cells transfected with the human ERα | ||||||||
| Binding to the ligand-binding domain | Racemic | IC50 (nM) | 57 | 1.2 | 1145 | [73] | ||
| of human ERα in vitro | ||||||||
| Binding to the ligand-binding domain | Racemic | IC50 (nM) | 68 | 1.4 | 25 | [73] | ||
| of human ERβ in vitro | ||||||||
| 2. In vivo | ||||||||
| Assay used | Type of 8-PN | Variable measured | 8-PN | E2 | Coumestrol | Genistein | Daidzein | Reference |
| Vascular permeability in mouse uterus | Racemic | Relative potency | <0.01 | 1 | <0.01 | <0.001 | No effect | [5] |
| Vaginal epithelial mitosis | Racemic | Relative potency | <0.001 | 1 | [5] | |||
| in OVX mice |
6. Endocrine Roles of Estrogen and 8-PN
6.1. ERα Tissue Expression
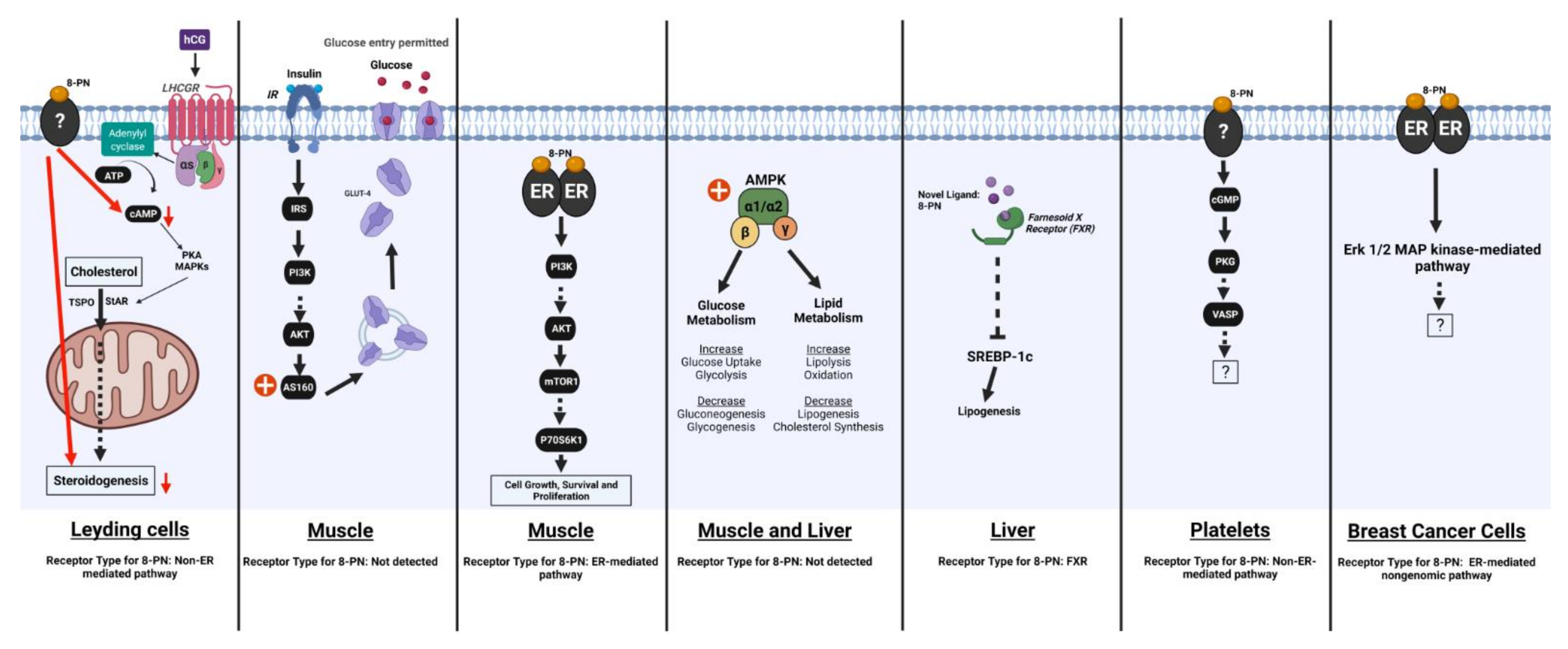
6.2. Energy Metabolism
6.2.1. Effects of 8-PN and Related Compounds on Energy Balance
6.2.2. Effects of 8-PN and Related Compounds on Lipid Metabolism
6.2.3. Effects of 8-PN and Related Compounds on Glucose Homeostasis and Insulin Sensitivity
6.3. Pituitary Function
6.3.1. Effects on LH and FSH Secretion
6.3.2. Effects on Other Pituitary Hormones
7. Regulation of Bone Homeostasis
8. Effects on Tumor Cells In Vitro
9. Other Beneficial and Adverse Effects of 8-PN
9.1. Cytotoxicity to Somatic Primary Cells
9.2. Effects on Gonadal Cells
9.3. Effects on Aromatase in Other Cell Types
9.4. Effects on Other Enzymes of Clinical Relevance
9.5. Effects on Blood Vessels
9.6. Effects on Inflammation and Immune Reactions
9.7. Effects on Platelets and Blood Coagulation
9.8. Effects on Muscle
9.9. Effects on Neurons and Behavior
9.10. Effects on Barrier Tissue Integrity
9.11. Antimicrobial Effects
9.12. Effects on Oxidative Stress
9.13. Modulation of Other Compound Toxicities
10. Conclusions and Future Research Needs
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tronina, T.; Popłoński, J.; Bartmańska, A. Flavonoids as phytoestrogenic components of hops and beer. Molecules 2020, 25, 4201. [Google Scholar] [CrossRef] [PubMed]
- Verzele, M. 100 years of hop chemistry and its relevance to brewing. J. Inst. Brew. 1986, 92, 32–48. [Google Scholar] [CrossRef]
- Goetz, P. Traitement des bouffées de chaleur par insuffisance ovarienne par l’extrait de houblon (Humulus lupulus). Rev. Phytothérapie Prat. 1990, 4, 13–15. [Google Scholar]
- Hesse, R.; Hoffmann, B.; Karg, H.; Vogt, K. Untersuchungen über den Nachweis von Phytoöstrogenen in Futterpflanzen und Hopfen mit Hilfe eines Rezeptortests 1. Zent. Für Veterinärmedizin Reihe A 1981, 28, 442–454. [Google Scholar] [CrossRef]
- Milligan, S.; Kalita, J.; Pocock, V.; Heyerick, A.; De Cooman, L.; Rong, H.; De Keukeleire, D. Oestrogenic activity of the hop phyto-oestrogen, 8-prenylnaringenin. Reprod. -Camb. 2002, 123, 235–242. [Google Scholar] [CrossRef]
- Fenselau, C.; Talalay, P. Is oestrogenic activity present in hops? Food Cosmet. Toxicol. 1973, 11, 597–603. [Google Scholar] [CrossRef]
- Milligan, S.; Kalita, J.; Heyerick, A.; Rong, H.; De Cooman, L.; De Keukeleire, D. Identification of a potent phytoestrogen in hops (Humulus lupulus L.) and beer. J. Clin. Endocrinol. Metab. 1999, 84, 2249. [Google Scholar] [CrossRef]
- Mizobuchi, S.; Sato, Y. A new flavanone with antifungal activity isolated from hops. Agric. Biol. Chem. 1984, 48, 2771–2775. [Google Scholar]
- Stevens, J.F.; Page, J.E. Xanthohumol and related prenylflavonoids from hops and beer: To your good health! Phytochemistry 2004, 65, 1317–1330. [Google Scholar] [CrossRef]
- Bolton, J.L.; Dunlap, T.L.; Hajirahimkhan, A.; Mbachu, O.; Chen, S.-N.; Chadwick, L.; Nikolic, D.; van Breemen, R.B.; Pauli, G.F.; Dietz, B.M.; et al. The multiple biological targets of hops and bioactive compounds. Chem. Res. Toxicol. 2019, 32, 222–233. [Google Scholar] [CrossRef] [Green Version]
- Karabin, M.; Hudcova, T.; Jelinek, L.; Dostalek, P. Biotransformations and biological activities of hop flavonoids. Biotechnol. Adv. 2015, 33, 1063–1090. [Google Scholar] [CrossRef] [PubMed]
- Kodama, S.; Yamamoto, A.; Sato, A.; Suzuki, K.; Yamashita, T.; Kemmei, T.; Taga, A.; Hayakawa, K. Enantioseparation of isoxanthohumol in beer by hydroxypropyl-γ-cyclodextrin-modified micellar electrokinetic chromatography. J. Agric. Food Chem. 2007, 55, 6547–6552. [Google Scholar] [CrossRef] [PubMed]
- Nikolic, D.; B Van Breemen, R. Analytical methods for quantitation of prenylated flavonoids from hops. Curr. Anal. Chem. 2013, 9, 71–85. [Google Scholar] [CrossRef]
- Kirin Beer University. Global Beer Consumption by Country in 2018. Available online: https://www.kirinholdings.com/en/newsroom/release/2019/1224_01.html (accessed on 30 January 2022).
- Stevens, J.F.; Taylor, A.W.; Deinzer, M.L. Quantitative analysis of xanthohumol and related prenylflavonoids in hops and beer by liquid chromatography–tandem mass spectrometry. J. Chromatogr. A 1999, 832, 97–107. [Google Scholar] [CrossRef]
- Buckett, L.; Schinko, S.; Urmann, C.; Riepl, H.; Rychlik, M. Stable Isotope Dilution Analysis of the Major Prenylated Flavonoids Found in Beer, Hop Tea, and Hops. Front. Nutr. 2020, 7, 619921. [Google Scholar] [CrossRef] [PubMed]
- Possemiers, S.; Bolca, S.; Grootaert, C.; Heyerick, A.; Decroos, K.; Dhooge, W.; De Keukeleire, D.; Rabot, S.; Verstraete, W.; Van de Wiele, T.; et al. The prenylflavonoid isoxanthohumol from hops (Humulus lupulus L.) is activated into the potent phytoestrogen 8-prenylnaringenin in vitro and in the human intestine. J. Nutr. 2006, 136, 1862–1867. [Google Scholar] [CrossRef] [Green Version]
- Česlová, L.; Holčapek, M.; Fidler, M.; Drštičková, J.; Lísa, M. Characterization of prenylflavonoids and hop bitter acids in various classes of Czech beers and hop extracts using high-performance liquid chromatography–mass spectrometry. J. Chromatogr. A 2009, 1216, 7249–7257. [Google Scholar] [CrossRef]
- Golubović, J.B.; Heath, E.; Košir, I.J.; Ogrinc, N.; Potočnik, D.; Strojnik, L.; Heath, D. Differences in the levels of the selected phytoestrogens and stable isotopes in organic vs. conventional hops and beer. Foods 2021, 10, 1839. [Google Scholar] [CrossRef]
- Martinez, S.E.; Lakowski, T.M.; Davies, N.M. Enantiospecific analysis of 8-prenylnaringenin in biological fluids by liquid-chromatography-electrospray ionization mass spectrometry: Application to preclinical pharmacokinetic investigations. Chirality 2014, 26, 419–426. [Google Scholar] [CrossRef]
- Ito, C.; Mizuno, T.; Matsuoka, M.; Kimura, Y.; Sato, K.; Kajiura, I.; Omura, M.; JU-Ichi, M.; Furukawa, H. A new flavonoid and other new components from Citrus plants. Chem. Pharm. Bull. 1988, 36, 3292–3295. [Google Scholar] [CrossRef] [Green Version]
- Clarke, D.B.; Bailey, V.; Lloyd, A. Determination of phytoestrogens in dietary supplements by LC-MS/MS. Food Addit. Contam. 2008, 25, 534–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schretter, C.; Langeder, J.; Freisinger, V.; Rollinger, J.M.; Grienke, U. Quantitative Analysis of Prenylated Constituents in Commercial Hops Samples Using Ultrahigh-Performance Supercritical Fluid Chromatography. Planta Med. 2020, 86, 1140–1147. [Google Scholar] [CrossRef]
- Coldham, N.; Sauer, M. Identification, quantitation and biological activity of phytoestrogens in a dietary supplement for breast enhancement. Food Chem. Toxicol. 2001, 39, 1211–1224. [Google Scholar] [CrossRef]
- Dhooghe, L.; Naessens, T.; Heyerick, A.; De Keukeleire, D.; Vlietinck, A.J.; Pieters, L.; Apers, S. Quantification of xanthohumol, isoxanthohumol, 8-prenylnaringenin, and 6-prenylnaringenin in hop extracts and derived capsules using secondary standards. Talanta 2010, 83, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Tekel’, J.; De Keukeleire, D.; Rong, H.; Daeseleire, E.; Van Peteghem, C. Determination of the hop-derived phytoestrogen, 8-prenylnaringenin, in beer by gas chromatography/mass spectrometry. J. Agric. Food Chem. 1999, 47, 5059–5063. [Google Scholar] [CrossRef]
- Maragou, N.C.; Rosenberg, E.; Thomaidis, N.S.; Koupparis, M.A. Direct determination of the estrogenic compounds 8-prenylnaringenin, zearalenone, α-and β-zearalenol in beer by liquid chromatography–mass spectrometry. J. Chromatogr. A 2008, 1202, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Qiu, X.; Nikolic, D.; Dahl, J.H.; van Breemen, R.B. Method development and validation for ultra-high-pressure LC/MS/MS determination of hop prenylflavonoids in human serum. J. AOAC Int. 2012, 95, 1744–1749. [Google Scholar] [CrossRef]
- Quifer-Rada, P.; Martínez-Huélamo, M.; Jáuregui, O.; Chiva-Blanch, G.; Estruch, R.N.; Lamuela-Raventós, R.M. Analytical condition setting a crucial step in the quantification of unstable polyphenols in acidic conditions: Analyzing prenylflavanoids in biological samples by liquid chromatography–electrospray ionization triple quadruple mass spectrometry. Anal. Chem. 2013, 85, 5547–5554. [Google Scholar] [CrossRef]
- Wyns, C.; Bolca, S.; De Keukeleire, D.; Heyerick, A. Development of a high-throughput LC/APCI-MS method for the determination of thirteen phytoestrogens including gut microbial metabolites in human urine and serum. J. Chromatogr. B 2010, 878, 949–956. [Google Scholar] [CrossRef]
- Prencipe, F.P.; Brighenti, V.; Rodolfi, M.; Mongelli, A.; dall’Asta, C.; Ganino, T.; Bruni, R.; Pellati, F. Development of a new high-performance liquid chromatography method with diode array and electrospray ionization-mass spectrometry detection for the metabolite fingerprinting of bioactive compounds in Humulus lupulus L. J. Chromatogr. A 2014, 1349, 50–59. [Google Scholar] [CrossRef]
- Moriya, H.; Tanaka, S.; Iida, Y.; Kitagawa, S.; Aizawa, S.I.; Taga, A.; Terashima, H.; Yamamoto, A.; Kodama, S. Chiral separation of isoxanthohumol and 8-prenylnaringenin in beer, hop pellets and hops by HPLC with chiral columns. Biomed. Chromatogr. 2018, 32, e4289. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, O.; Bohlmann, R.; Schleuning, W.-D.; Schulze-Forster, K.; Hümpel, M. Development of a radioimmunoassay for the quantitative determination of 8-prenylnaringenin in biological matrices. J. Agric. Food Chem. 2005, 53, 2881–2889. [Google Scholar] [CrossRef] [PubMed]
- Wyns, C.; Derycke, L.; Soenen, B.; Bolca, S.; Deforce, D.; Bracke, M.; Heyerick, A. Production of monoclonal antibodies against hop-derived (Humulus lupulus L.) prenylflavonoids and the development of immunoassays. Talanta 2011, 85, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Philbin, C.S.; Schwartz, S.J. Resolution of diastereomeric flavonoid (1S)-(−)-camphanic acid esters via reversed-phase HPLC. Phytochemistry 2007, 68, 1206–1211. [Google Scholar] [CrossRef]
- Calvo-Castro, L.A.; Burkard, M.; Sus, N.; Scheubeck, G.; Leischner, C.; Lauer, U.M.; Bosy-Westphal, A.; Hund, V.; Busch, C.; Venturelli, S.; et al. The Oral Bioavailability of 8-Prenylnaringenin from Hops (Humulus Lupulus L.) in Healthy Women and Men is Significantly Higher than that of its Positional Isomer 6-Prenylnaringenin in a Randomized Crossover Trial. Mol. Nutr. Food Res. 2018, 62, 1700838. [Google Scholar] [CrossRef]
- Rad, M.; Hümpel, M.; Schaefer, O.; Schoemaker, R.; Schleuning, W.D.; Cohen, A.; Burggraaf, J. Pharmacokinetics and systemic endocrine effects of the phyto-oestrogen 8-prenylnaringenin after single oral doses to postmenopausal women. Br. J. Clin. Pharmacol. 2006, 62, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Nikolic, D.; Li, Y.; Chadwick, L.R.; van Breemen, R.B. In vitro studies of intestinal permeability and hepatic and intestinal metabolism of 8-prenylnaringenin, a potent phytoestrogen from hops (Humulus lupulus L.). Pharm. Res. 2006, 23, 864–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolic, D.; Li, Y.; Chadwick, L.R.; Grubjesic, S.; Schwab, P.; Metz, P.; van Breemen, R.B. Metabolism of 8-prenylnaringenin, a potent phytoestrogen from hops (Humulus lupulus), by human liver microsomes. Drug Metab. Dispos. 2004, 32, 272–279. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.-B.; Nikolić, D.; Lankin, D.C.; Simmler, C.; Chen, S.-N.; Ramos Alvarenga, R.F.; Liu, Y.; Pauli, G.F.; van Breemen, R.B. Formation of (2 R)-and (2 S)-8-Prenylnaringenin Glucuronides by Human UDP-Glucuronosyltransferases. J. Agric. Food Chem. 2019, 67, 11650–11656. [Google Scholar] [CrossRef] [PubMed]
- Paraiso, I.L.; Plagmann, L.S.; Yang, L.; Zielke, R.; Gombart, A.F.; Maier, C.S.; Sikora, A.E.; Blakemore, P.R.; Stevens, J.F. Reductive Metabolism of Xanthohumol and 8-Prenylnaringenin by the Intestinal Bacterium Eubacterium ramulus. Mol. Nutr. Food Res. 2019, 63, 1800923. [Google Scholar] [CrossRef] [PubMed]
- Bartmańska, A.; Tronina, T.; Huszcza, E. Biotransformation of the phytoestrogen 8-prenylnaringenin. Z. Für Nat. C 2010, 65, 603–606. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, S.-H.; Kang, B.Y.; Lee, I.-S. Microbial metabolites of 8-prenylnaringenin, an estrogenic prenylflavanone. Arch. Pharmacal Res. 2008, 31, 1241–1246. [Google Scholar] [CrossRef] [PubMed]
- Bartmańska, A.; Tronina, T.; Huszcza, E. Transformation of 8-prenylnaringenin by Absidia coerulea and Beauveria bassiana. Bioorganic Med. Chem. Lett. 2012, 22, 6451–6453. [Google Scholar] [CrossRef]
- Tronina, T.; Huszcza, E. Microbial sulfation of 8-prenylnaringenin. Z. Für Nat. C 2013, 68, 231–235. [Google Scholar] [CrossRef]
- Coldham, N.; Horton, R.; Byford, M.; Sauer, M. A binary screening assay for pro-oestrogens in food: Metabolic activation using hepatic microsomes and detection with oestrogen sensitive recombinant yeast cells. Food Addit. Contam. 2002, 19, 1138–1147. [Google Scholar] [CrossRef]
- Nikolic, D.; Li, Y.; Chadwick, L.R.; Pauli, G.F.; van Breemen, R.B. Metabolism of xanthohumol and isoxanthohumol, prenylated flavonoids from hops (Humulus lupulus L.), by human liver microsomes. J. Mass Spectrom. 2005, 40, 289–299. [Google Scholar] [CrossRef]
- Guo, J.; Nikolic, D.; Chadwick, L.R.; Pauli, G.F.; van Breemen, R.B. Identification of human hepatic cytochrome P450 enzymes involved in the metabolism of 8-prenylnaringenin and isoxanthohumol from hops (Humulus lupulus L.). Drug Metab. Dispos. 2006, 34, 1152–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolca, S.; Possemiers, S.; Maervoet, V.; Huybrechts, I.; Heyerick, A.; Vervarcke, S.; Depypere, H.; De Keukeleire, D.; Bracke, M.; De Henauw, S.; et al. Microbial and dietary factors associated with the 8-prenylnaringenin producer phenotype: A dietary intervention trial with fifty healthy post-menopausal Caucasian women. Br. J. Nutr. 2007, 98, 950–959. [Google Scholar] [CrossRef] [Green Version]
- Bolca, S.; Wyns, C.; Possemiers, S.; Depypere, H.; De Keukeleire, D.; Bracke, M.; Verstraete, W.; Heyerick, A. Cosupplementation of isoflavones, prenylflavonoids, and lignans alters human exposure to phytoestrogen-derived 17 β-estradiol equivalents. J. Nutr. 2009, 139, 2293–2300. [Google Scholar] [CrossRef] [Green Version]
- Possemiers, S.; Heyerick, A.; Robbens, V.; De Keukeleire, D.; Verstraete, W. Activation of proestrogens from hops (Humulus lupulus L.) by intestinal microbiota; conversion of isoxanthohumol into 8-prenylnaringenin. J. Agric. Food Chem. 2005, 53, 6281–6288. [Google Scholar] [CrossRef] [Green Version]
- Possemiers, S.; Rabot, S.; Espín, J.C.; Bruneau, A.; Philippe, C.; González-Sarrías, A.; Heyerick, A.; Tomás-Barberán, F.A.; De Keukeleire, D.; Verstraete, W.; et al. Eubacterium limosum activates isoxanthohumol from hops (Humulus lupulus L.) into the potent phytoestrogen 8-prenylnaringenin in vitro and in rat intestine. J. Nutr. 2008, 138, 1310–1316. [Google Scholar] [CrossRef]
- Hanske, L.; Loh, G.; Sczesny, S.; Blaut, M.; Braune, A. Recovery and metabolism of xanthohumol in germ-free and human microbiota-associated rats. Mol. Nutr. Food Res. 2010, 54, 1405–1413. [Google Scholar] [CrossRef] [PubMed]
- Terao, J.; Mukai, R. Prenylation modulates the bioavailability and bioaccumulation of dietary flavonoids. Arch. Biochem. Biophys. 2014, 559, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.W.; Cooney, J.; Jensen, D.; Li, Y.; Paxton, J.W.; Birch, N.P.; Scheepens, A. Hop-derived prenylflavonoids are substrates and inhibitors of the efflux transporter breast cancer resistance protein (BCRP/ABCG 2). Mol. Nutr. Food Res. 2014, 58, 2099–2110. [Google Scholar] [CrossRef]
- Wesołowska, O.; Wiśniewski, J.; Środa, K.; Krawczenko, A.; Bielawska-Pohl, A.; Paprocka, M.; Duś, D.; Michalak, K. 8-Prenylnaringenin is an inhibitor of multidrug resistance-associated transporters, P-glycoprotein and MRP1. Eur. J. Pharmacol. 2010, 644, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Caccamese, S.; Manna, L.; Scivoli, G. Chiral HPLC separation and CD spectra of the C-2 diastereomers of naringin in grapefruit during maturation. Chirality Pharmacol. Biol. Chem. Conseq. Mol. Asymmetry 2003, 15, 661–667. [Google Scholar] [CrossRef]
- Zhang, J.-Q.; Cai, W.-Q.; Zhou, D.-S.; Su, B.-Y. Distribution and differences of estrogen receptor beta immunoreactivity in the brain of adult male and female rats. Brain Res. 2002, 935, 73–80. [Google Scholar] [CrossRef]
- Takamura-Enya, T.; Ishihara, J.; Tahara, S.; Goto, S.; Totsuka, Y.; Sugimura, T.; Wakabayashi, K. Analysis of estrogenic activity of foodstuffs and cigarette smoke condensates using a yeast estrogen screening method. Food Chem. Toxicol. 2003, 41, 543–550. [Google Scholar] [CrossRef]
- Zierau, O.; Gester, S.; Schwab, P.; Metz, P.; Kolba, S.; Wulf, M.; Vollmer, G. Estrogenic activity of the phytoestrogens naringenin, 6-(1, 1-dimethylallyl) naringenin and 8-prenylnaringenin. Planta Med. 2002, 68, 449–451. [Google Scholar] [CrossRef]
- Overk, C.R.; Yao, P.; Chadwick, L.R.; Nikolic, D.; Sun, Y.; Cuendet, M.A.; Deng, Y.; Hedayat, A.; Pauli, G.F.; Farnsworth, N.R.; et al. Comparison of the in vitro estrogenic activities of compounds from hops (Humulus lupulus) and red clover (Trifolium pratense). J. Agric. Food Chem. 2005, 53, 6246–6253. [Google Scholar] [CrossRef] [Green Version]
- Matsumura, A.; Ghosh, A.; Pope, G.; Darbre, P. Comparative study of oestrogenic properties of eight phytoestrogens in MCF7 human breast cancer cells. J. Steroid Biochem. Mol. Biol. 2005, 94, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, O.; Hümpel, M.; Fritzemeier, K.-H.; Bohlmann, R.; Schleuning, W.-D. 8-Prenyl naringenin is a potent ERα selective phytoestrogen present in hops and beer. J. Steroid Biochem. Mol. Biol. 2003, 84, 359–360. [Google Scholar] [CrossRef]
- Zierau, O.; Hamann, J.; Tischer, S.; Schwab, P.; Metz, P.; Vollmer, G.; Gutzeit, H.O.; Scholz, S. Naringenin-type flavonoids show different estrogenic effects in mammalian and teleost test systems. Biochem. Biophys. Res. Commun. 2005, 326, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Kretzschmar, G.; Zierau, O.; Wober, J.; Tischer, S.; Metz, P.; Vollmer, G. Prenylation has a compound specific effect on the estrogenicity of naringenin and genistein. J. Steroid Biochem. Mol. Biol. 2010, 118, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Helle, J.; Kräker, K.; Bader, M.I.; Keiler, A.M.; Zierau, O.; Vollmer, G.; Welsh, J.; Kretzschmar, G. Assessment of the proliferative capacity of the flavanones 8-prenylnaringenin, 6-(1.1-dimethylallyl) naringenin and naringenin in MCF-7 cells and the rat mammary gland. Mol. Cell. Endocrinol. 2014, 392, 125–135. [Google Scholar] [CrossRef]
- Bovee, T.F.; Helsdingen, R.J.; Rietjens, I.M.; Keijer, J.; Hoogenboom, R.L. Rapid yeast estrogen bioassays stably expressing human estrogen receptors α and β, and green fluorescent protein: A comparison of different compounds with both receptor types. J. Steroid Biochem. Mol. Biol. 2004, 91, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Amer, D.A.; Kretzschmar, G.; Müller, N.; Stanke, N.; Lindemann, D.; Vollmer, G. Activation of transgenic estrogen receptor-beta by selected phytoestrogens in a stably transduced rat serotonergic cell line. J. Steroid Biochem. Mol. Biol. 2010, 120, 208–217. [Google Scholar] [CrossRef]
- van de Schans, M.G.; Ritschel, T.; Bovee, T.F.; Sanders, M.G.; de Waard, P.; Gruppen, H.; Vincken, J.P. Involvement of a hydrophobic pocket and helix 11 in determining the modes of action of prenylated flavonoids and isoflavonoids in the human estrogen receptor. ChemBioChem 2015, 16, 2668–2677. [Google Scholar] [CrossRef]
- Amer, D.A.; Jähne, M.; Weigt, C.; Kretzschmar, G.; Vollmer, G. Effect of 17β-estradiol and flavonoids on the regulation of expression of newly identified oestrogen responsive genes in a rat raphe nuclei-derived cell line. J. Cell. Physiol. 2012, 227, 3434–3445. [Google Scholar] [CrossRef]
- Bovee, T.F.; Schoonen, W.G.; Hamers, A.R.; Bento, M.J.; Peijnenburg, A.A. Screening of synthetic and plant-derived compounds for (anti) estrogenic and (anti) androgenic activities. Anal. Bioanal. Chem. 2008, 390, 1111–1119. [Google Scholar] [CrossRef]
- Zierau, O.; Morrissey, C.; Watson, R.W.G.; Schwab, P.; Kolba, S.; Metz, P.; Vollmer, G. Antiandrogenic activity of the phytoestrogens naringenin, 6-(1, 1-dimethylallyl) naringenin and 8-prenylnaringenin. Planta Med. 2003, 69, 856–858. [Google Scholar] [PubMed]
- Roelens, F.; Heldring, N.; Dhooge, W.; Bengtsson, M.; Comhaire, F.; Gustafsson, J.-Å.; Treuter, E.; De Keukeleire, D. Subtle side-chain modifications of the hop phytoestrogen 8-prenylnaringenin result in distinct agonist/antagonist activity profiles for estrogen receptors α and β. J. Med. Chem. 2006, 49, 7357–7365. [Google Scholar] [CrossRef]
- Bjornstrom, L.; Sjoberg, M. Mechanisms of estrogen receptor signaling: Convergence of genomic and nongenomic actions on target genes. Mol. Endocrinol. 2005, 19, 833–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunelli, E.; Minassi, A.; Appendino, G.; Moro, L. 8-Prenylnaringenin, inhibits estrogen receptor-α mediated cell growth and induces apoptosis in MCF-7 breast cancer cells. J. Steroid Biochem. Mol. Biol. 2007, 107, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Aichinger, G.; Bliem, G.; Marko, D. Systemically Achievable Doses of Beer Flavonoids Induce Estrogenicity in Human Endometrial Cells and Cause Synergistic Effects With Selected Pesticides. Front. Nutr. 2021, 8, 691872. [Google Scholar] [CrossRef] [PubMed]
- Aichinger, G.; Beisl, J.; Marko, D. The hop polyphenols xanthohumol and 8-prenyl-naringenin antagonize the estrogenic effects of fusarium mycotoxins in human endometrial cancer cells. Front. Nutr. 2018, 5, 85. [Google Scholar] [CrossRef] [Green Version]
- Diel, P.; Thomae, R.B.; Caldarelli, A.; Zierau, O.; Kolba, S.; Schmidt, S.; Schwab, P.; Metz, P.; Vollmer, G. Regulation of gene expression by 8-prenylnaringenin in uterus and liver of Wistar rats. Planta Med. 2004, 70, 39–44. [Google Scholar] [PubMed]
- Keiler, A.M.; Dörfelt, P.; Chatterjee, N.; Helle, J.; Bader, M.I.; Vollmer, G.; Kretzschmar, G.; Kuhlee, F.; Thieme, D.; Zierau, O.; et al. Assessment of the effects of naringenin-type flavanones in uterus and vagina. J. Steroid Biochem. Mol. Biol. 2015, 145, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Zierau, O.; Kretzschmar, G.; Möller, F.; Weigt, C.; Vollmer, G. Time dependency of uterine effects of naringenin type phytoestrogens in vivo. Mol. Cell. Endocrinol. 2008, 294, 92–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowe, J.; Li, X.F.; Kinsey-Jones, J.; Heyerick, A.; Brain, S.; Milligan, S.; O’Byrne, K. The hop phytoestrogen, 8-prenylnaringenin, reverses the ovariectomy-induced rise in skin temperature in an animal model of menopausal hot flushes. J. Endocrinol. 2006, 191, 399–405. [Google Scholar] [CrossRef]
- Miyamoto, M.; Matsushita, Y.; Kiyokawa, A.; Fukuda, C.; Sugano, M.; Akiyama, T. Prenylflavonoids: A new class of non-steroidal phytoestrogen (part 2). estrogenic effects of 18-isopentenylnaringenin on bone metabolism. Planta Med. 1998, 64, 516–519. [Google Scholar] [CrossRef] [PubMed]
- Overk, C.R.; Guo, J.; Chadwick, L.R.; Lantvit, D.D.; Minassi, A.; Appendino, G.; Chen, S.-N.; Lankin, D.C.; Farnsworth, N.R.; Pauli, G.F.; et al. In vivo estrogenic comparisons of Trifolium pratense (red clover) Humulus lupulus (hops), and the pure compounds isoxanthohumol and 8-prenylnaringenin. Chem. Biol. Interact. 2008, 176, 30–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rimoldi, G.; Christoffel, J.; Wuttke, W. Morphologic changes induced by oral long-term treatment with 8-prenylnaringenin in the uterus, vagina, and mammary gland of castrated rats. Menopause 2006, 13, 669–677. [Google Scholar] [CrossRef]
- Nasri, A.; Pohjanvirta, R. In vitro estrogenic, cytotoxic, and genotoxic profiles of the xenoestrogens 8-prenylnaringenine, genistein and tartrazine. Environ. Sci. Pollut. Res. 2021, 28, 27988–27997. [Google Scholar] [CrossRef] [PubMed]
- Jensen, E.V. Basic guides to the mechanism of estrogen action. Recent Progr. Horm. Res. 1962, 18, 387–414. [Google Scholar]
- Kuiper, G.; Enmark, E.; Pelto-Huikko, M.; Nilsson, S.; Gustafsson, J.-A. Cloning of a novel receptor expressed in rat prostate and ovary. Proc. Natl. Acad. Sci. USA 1996, 93, 5925–5930. [Google Scholar] [CrossRef] [Green Version]
- Gruber, C.J.; Tschugguel, W.; Schneeberger, C.; Huber, J.C. Production and actions of estrogens. N. Engl. J. Med. 2002, 346, 340–352. [Google Scholar] [CrossRef]
- Lim, Y.C.; Li, L.; Desta, Z.; Zhao, Q.; Rae, J.M.; Flockhart, D.A.; Skaar, T.C. Endoxifen, a secondary metabolite of tamoxifen, and 4-OH-tamoxifen induce similar changes in global gene expression patterns in MCF-7 breast cancer cells. J. Pharmacol. Exp. Ther. 2006, 318, 503–512. [Google Scholar] [CrossRef] [Green Version]
- Kuiper, G.G.; Lemmen, J.G.; Carlsson, B.; Corton, J.C.; Safe, S.H.; Van Der Saag, P.T.; Van Der Burg, B.; Gustafsson, J.-A.K. Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor β. Endocrinology 1998, 139, 4252–4263. [Google Scholar] [CrossRef]
- Ososki, A.L.; Kennelly, E.J. Phytoestrogens: A review of the present state of research. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2003, 17, 845–869. [Google Scholar] [CrossRef]
- Simons, R.; Gruppen, H.; Bovee, T.F.; Verbruggen, M.A.; Vincken, J.-P. Prenylated isoflavonoids from plants as selective estrogen receptor modulators (phytoSERMs). Food Funct. 2012, 3, 810–827. [Google Scholar] [CrossRef]
- Koehler, K.F.; Helguero, L.A.; Haldosén, L.-A.; Warner, M.; Gustafsson, J.-A. Reflections on the discovery and significance of estrogen receptor β. Endocr. Rev. 2005, 26, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Jordan, V.C. Chemoprevention of breast cancer with selective oestrogen-receptor modulators. Nat. Rev. Cancer 2007, 7, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.; Lange, I.; Daxenberger, A.; Meyer, H. Tissue-specific expression pattern of estrogen receptors (ER): Quantification of ERα and ERβ mRNA with real-time RT-PCR. Apmis 2001, 109, S540–S550. [Google Scholar] [CrossRef]
- Mauvais-Jarvis, F.; Clegg, D.J.; Hevener, A.L. The role of estrogens in control of energy balance and glucose homeostasis. Endocr. Rev. 2013, 34, 309–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutson, D.D.; Gurrala, R.; Ogola, B.O.; Zimmerman, M.A.; Mostany, R.; Satou, R.; Lindsey, S.H. Estrogen receptor profiles across tissues from male and female Rattus norvegicus. Biol. Sex Differ. 2019, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Simerly, R.; Swanson, L.; Chang, C.; Muramatsu, M. Distribution of androgen and estrogen receptor mRNA-containing cells in the rat brain: An in situ hybridization study. J. Comp. Neurol. 1990, 294, 76–95. [Google Scholar] [CrossRef] [PubMed]
- Simonian, S.X.; Herbison, A.E. Differential expression of estrogen receptor alpha and beta immunoreactivity by oxytocin neurons of rat paraventricular nucleus. J. Neuroendocrinol. 1997, 9, 803–806. [Google Scholar] [CrossRef]
- Voisin, D.; Simonian, S.; Herbison, A. Identification of estrogen receptor-containing neurons projecting to the rat supraoptic nucleus. Neuroscience 1997, 78, 215–228. [Google Scholar] [CrossRef]
- Wilkinson, H.A.; Dahllund, J.; Liu, H.; Yudkovitz, J.; Cai, S.-J.; Nilsson, S.; Schaeffer, J.M.; Mitra, S.W. Identification and characterization of a functionally distinct form of human estrogen receptor β. Endocrinology 2002, 143, 1558–1561. [Google Scholar] [CrossRef]
- Shima, N.; Yamaguchi, Y.; Yuri, K. Distribution of estrogen receptor β mRNA-containing cells in ovariectomized and estrogen-treated female rat brain. Anat. Sci. Int. 2003, 78, 85–97. [Google Scholar] [CrossRef]
- Merchenthaler, I.; Lane, M.V.; Numan, S.; Dellovade, T.L. Distribution of estrogen receptor α and β in the mouse central nervous system: In vivo autoradiographic and immunocytochemical analyses. J. Comp. Neurol. 2004, 473, 270–291. [Google Scholar] [CrossRef]
- Manoranjan, B.; Salehi, F.; Scheithauer, B.; Rotondo, F.; Kovacs, K.; Cusimano, M. Estrogen receptors α and β immunohistochemical expression: Clinicopathological correlations in pituitary adenomas. Anticancer. Res. 2010, 30, 2897–2904. [Google Scholar] [PubMed]
- Christoffel, J. Endocrine potential of Genistein, Resveratrol and 8-Prenylnaringenin in Gonadal and Thyroid Axes and Related Organs. Ph.D. Thesis, Faculty of Mathematics and Natural Sciences, Christian-Albrecht University of Kiel, Kiel, Germany, 2004. [Google Scholar]
- Costa, R.; Rodrigues, I.; Guardão, L.; Rocha-Rodrigues, S.; Silva, C.; Magalhães, J.; Ferreira-de-Almeida, M.; Negrão, R.; Soares, R. Xanthohumol and 8-prenylnaringenin ameliorate diabetic-related metabolic dysfunctions in mice. J. Nutr. Biochem. 2017, 45, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, E.E.; Allister, E.M.; Sutherland, B.G.; Telford, D.E.; Sawyez, C.G.; Edwards, J.Y.; Markle, J.M.; Hegele, R.A.; Huff, M.W. Naringenin prevents dyslipidemia, apolipoprotein B overproduction, and hyperinsulinemia in LDL receptor–null mice with diet-induced insulin resistance. Diabetes 2009, 58, 2198–2210. [Google Scholar] [CrossRef] [Green Version]
- Assini, J.M.; Mulvihill, E.E.; Sutherland, B.G.; Telford, D.E.; Sawyez, C.G.; Felder, S.L.; Chhoker, S.; Edwards, J.Y.; Gros, R.; Huff, M.W.; et al. Naringenin prevents cholesterol-induced systemic inflammation, metabolic dysregulation, and atherosclerosis in Ldlr−/−mice [S]. J. Lipid Res. 2013, 54, 711–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assini, J.M.; Mulvihill, E.E.; Burke, A.C.; Sutherland, B.G.; Telford, D.E.; Chhoker, S.S.; Sawyez, C.G.; Drangova, M.; Adams, A.C.; Kharitonenkov, A.; et al. Naringenin prevents obesity, hepatic steatosis, and glucose intolerance in male mice independent of fibroblast growth factor 21. Endocrinology 2015, 156, 2087–2102. [Google Scholar] [CrossRef] [Green Version]
- Murugesan, N.; Woodard, K.; Ramaraju, R.; Greenway, F.L.; Coulter, A.A.; Rebello, C.J. Naringenin increases insulin sensitivity and metabolic rate: A case study. J. Med. Food 2020, 23, 343–348. [Google Scholar] [CrossRef]
- Park, M.; Kim, K.; Lee, Y.M.; Rhyu, M.R.; Kim, H.Y. Naringenin stimulates cholecystokinin secretion in STC-1 cells. Nutr. Res. Pract. 2014, 8, 146–150. [Google Scholar] [CrossRef] [Green Version]
- Miquilena-Colina, M.E.; Lima-Cabello, E.; Sánchez-Campos, S.; García-Mediavilla, M.V.; Fernández-Bermejo, M.; Lozano-Rodríguez, T.; Vargas-Castrillón, J.; Buqué, X.; Ochoa, B.; Aspichueta, P.; et al. Hepatic fatty acid translocase CD36 upregulation is associated with insulin resistance, hyperinsulinaemia and increased steatosis in non-alcoholic steatohepatitis and chronic hepatitis C. Gut 2011, 60, 1394–1402. [Google Scholar] [CrossRef]
- Hagberg, C.E.; Falkevall, A.; Wang, X.; Larsson, E.; Huusko, J.; Nilsson, I.; van Meeteren, L.A.; Samen, E.; Lu, L.; Vanwildemeersch, M.; et al. Vascular endothelial growth factor B controls endothelial fatty acid uptake. Nature 2010, 464, 917–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Broderick, D.; Campbell, Y.; Gombart, A.F.; Stevens, J.F.; Jiang, Y.; Hsu, V.L.; Bisson, W.H.; Maier, C.S. Conformational modulation of the farnesoid X receptor by prenylflavonoids: Insights from hydrogen deuterium exchange mass spectrometry (HDX-MS), fluorescence titration and molecular docking studies. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2016, 1864, 1667–1677. [Google Scholar] [CrossRef] [Green Version]
- Klip, A. The many ways to regulate glucose transporter 4. Appl. Physiol. Nutr. Metab. 2009, 34, 481–487. [Google Scholar] [CrossRef]
- Christoffel, J.; Rimoldi, G.; Wuttke, W. Effects of 8-prenylnaringenin on the hypothalamo-pituitary-uterine axis in rats after 3-month treatment. J. Endocrinol. 2006, 188, 397–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarry, H.; Sprenger, M.; Wuttke, W. Rates of release of GABA and catecholamines in the mediobasal hypothalamus of ovariectomized and ovariectomized estrogen-treated rats: Correlation with blood prolactin levels. Neuroendocrinology 1986, 44, 422–428. [Google Scholar] [CrossRef]
- Tataryn, I.V.; Meldrum, D.R.; Lu, K.; Fruraar, A.M.; Judd, H.L. LH, FSH and skin temperature during the menopausal hot flas. J. Clin. Endocrinol. Metab. 1979, 49, 152–154. [Google Scholar] [CrossRef] [PubMed]
- Gambone, J.; Meldrum, D.; Laufer, L.; Chang, R.; Lu, J.; Judd, H. Further delineation of hypothalamic dysfunction responsible for menopausal hot flashes. J. Clin. Endocrinol. Metab. 1984, 59, 1097–1102. [Google Scholar] [CrossRef]
- Heyerick, A.; Vervarcke, S.; Depypere, H.; Bracke, M.; De Keukeleire, D. A first prospective, randomized, double-blind, placebo-controlled study on the use of a standardized hop extract to alleviate menopausal discomforts. Maturitas 2006, 54, 164–175. [Google Scholar] [CrossRef]
- Stefaneanu, L.; Kovacs, K.; Horvath, E.; Lloyd, R.; Buchfelder, M.; Fahlbusch, R.; Smyth, H. In situ hybridization study of estrogen receptor messenger ribonucleic acid in human adenohypophysial cells and pituitary adenomas. J. Clin. Endocrinol. Metab. 1994, 78, 83–88. [Google Scholar]
- Szczepankiewicz, D.; Skrzypski, M.; Pruszynska-Oszmalek, E.; Zimmermann, D.; Andralojc, K.; Kaczmarek, P.; Wojciechowicz, T.; Sassek, M.; Nowak, K. Importance of ghrelin in hypothalamus—Pituitary axis on growth hormone release during normal pregnancy in the rat. J. Physiol. Pharmacol. 2010, 61, 443. [Google Scholar]
- Ho, K.K.; O’Sullivan, A.J.; Weissberger, A.J.; Kelly, J.J. Sex steroid regulation of growth hormone secretion and action. Horm. Res. Paediatr. 1996, 45, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Rupa, R.; Voellger, B.; Kirches, E.; Mawrin, C.; Firsching, R. 8-Prenylnaringenin decreases hormone expression in GH3 pituitary adenoma cells of the rat. In Proceedings of the 15th European Congress of Endocrinology, Copenhagen, Denmark, 27 April–1 May 2013; Volume 32, p. 839. [Google Scholar]
- Bottner, M.; Christoffel, J.; Wuttke, W. Effects of long-term treatment with 8-prenylnaringenin and oral estradiol on the GH-IGF-1 axis and lipid metabolism in rats. J. Endocrinol. 2008, 198, 395–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ming, L.; Ge, B.; Wang, M.; Chen, K. Comparison between 8-prenylnarigenin and narigenin concerning their activities on promotion of rat bone marrow stromal cells’ osteogenic differentiation in vitro. Cell Prolif. 2012, 45, 508–515. [Google Scholar] [CrossRef]
- Ming, L.-G.; Lv, X.; Ma, X.-N.; Ge, B.-F.; Zhen, P.; Song, P.; Zhou, J.; Ma, H.-P.; Xian, C.J.; Chen, K.-M.; et al. The prenyl group contributes to activities of phytoestrogen 8-prenynaringenin in enhancing bone formation and inhibiting bone resorption in vitro. Endocrinology 2013, 154, 1202–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, D.; Kang, L.; Ma, Y.; Chen, H.; Kuang, H.; Huang, Q.; He, M.; Peng, W. Effects and mechanisms of 8-prenylnaringenin on osteoblast MC 3T3-E1 and osteoclast-like cells RAW 264.7. Food Sci. Nutr. 2014, 2, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Hümpel, M.; Isaksson, P.; Schaefer, O.; Kaufmann, U.; Ciana, P.; Maggi, A.; Schleuning, W.-D. Tissue specificity of 8-prenylnaringenin: Protection from ovariectomy induced bone loss with minimal trophic effects on the uterus. J. Steroid Biochem. Mol. Biol. 2005, 97, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Sehmisch, S.; Hammer, F.; Christoffel, J.; Seidlova-Wuttke, D.; Tezval, M.; Wuttke, W.; Stuermer, K.M.; Stuermer, E.K. Comparison of the phytohormones genistein, resveratrol and 8-prenylnaringenin as agents for preventing osteoporosis. Planta Med. 2008, 74, 794–801. [Google Scholar] [CrossRef]
- Hoffmann, D.B.; Griesel, M.H.; Brockhusen, B.; Tezval, M.; Komrakova, M.; Menger, B.; Wassmann, M.; Stuermer, K.M.; Sehmisch, S. Effects of 8-prenylnaringenin and whole-body vibration therapy on a rat model of osteopenia. J. Nutr. Metab. 2016, 2016, 6893137. [Google Scholar] [CrossRef] [Green Version]
- Pedrera-Zamorano, J.D.; Lavado-Garcia, J.M.; Roncero-Martin, R.; Calderon-Garcia, J.F.; Rodriguez-Dominguez, T.; Canal-Macias, M.L. Effect of beer drinking on ultrasound bone mass in women. Nutrition 2009, 25, 1057–1063. [Google Scholar] [CrossRef]
- Delmulle, L.; Bellahcene, A.; Dhooge, W.; Comhaire, F.; Roelens, F.; Huvaere, K.; Heyerick, A.; Castronovo, V.; De Keukeleire, D. Anti-proliferative properties of prenylated flavonoids from hops (Humulus lupulus L.) in human prostate cancer cell lines. Phytomedicine 2006, 13, 732–734. [Google Scholar] [CrossRef]
- Delmulle, L.; Berghe, T.V.; Keukeleire, D.D.; Vandenabeele, P. Treatment of PC-3 and DU145 prostate cancer cells by prenylflavonoids from hop (Humulus lupulus L.) induces a caspase-independent form of cell death. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2008, 22, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Busch, C.; Noor, S.; Leischner, C.; Burkard, M.; Lauer, U.M.; Venturelli, S. Anti-proliferative activity of hop-derived prenylflavonoids against human cancer cell lines. Wien. Med. Wochenschr. 2015, 165, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Gąsiorowska, J.; Teisseyre, A.; Uryga, A.; Michalak, K. The influence of 8-prenylnaringenin on the activity of voltage-gated Kv1. 3 potassium channels in human Jurkat T cells. Cell. Mol. Biol. Lett. 2012, 17, 559–570. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, Y.; Wang, J.; Chen, J.; Aggarwal, B.; Pang, X.; Liu, M. Xanthohumol, a prenylated chalcone derived from hops, suppresses cancer cell invasion through inhibiting the expression of CXCR4 chemokine receptor. Curr. Mol. Med. 2012, 12, 153–162. [Google Scholar] [CrossRef]
- Monteiro, R.; Faria, A.; Azevedo, I.; Calhau, C. Modulation of breast cancer cell survival by aromatase inhibiting hop (Humulus lupulus L.) flavonoids. J. Steroid Biochem. Mol. Biol. 2007, 105, 124–130. [Google Scholar] [CrossRef]
- Diller, R.A.; Riepl, H.M.; Rose, O.; Frias, C.; Henze, G.; Prokop, A. Ability of prenylflavanones present in hops to induce apoptosis in a human Burkitt lymphoma cell line. Planta Med. 2007, 73, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Venturelli, S.; Niessner, H.; Sinnberg, T.; Berger, A.; Burkard, M.; Urmann, C.; Donaubauer, K.; Böcker, A.; Leischner, C.; Riepl, H.; et al. 6-and 8-Prenylnaringenin, novel natural histone deacetylase inhibitors found in hops, exert antitumor activity on melanoma cells. Cell. Physiol. Biochem. 2018, 51, 543–556. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, D.; Liu, W.; Li, C.; Hao, L.; Zhang, G.; Deng, S.; Yang, R.; Qin, J.; Li, J.; et al. Cytotoxic Activity and Related Mechanisms of Prenylflavonoids Isolated from Mallotus conspurcatus Croizat. Chem. Biodivers. 2019, 16, e1800465. [Google Scholar] [CrossRef]
- Koosha, S.; Mohamed, Z.; Sinniah, A.; Ibrahim, Z.A.; Seyedan, A.; Alshawsh, M.A. Antiproliferative and apoptotic activities of 8-prenylnaringenin against human colon cancer cells. Life Sci. 2019, 232, 116633. [Google Scholar] [CrossRef]
- Stompor, M.; Uram, Ł.; Podgórski, R. In vitro effect of 8-prenylnaringenin and naringenin on fibroblasts and glioblastoma cells-cellular accumulation and cytotoxicity. Molecules 2017, 22, 1092. [Google Scholar] [CrossRef] [Green Version]
- Rong, H.; Boterberg, T.; Maubach, J.; Stove, C.; Depypere, H.; Van Slambrouck, S.; Serreyn, R.; De Keukeleire, D.; Mareel, M.; Bracke, M.; et al. 8-Prenylnaringenin, the phytoestrogen in hops and beer, upregulates the function of the E-cadherin/catenin complex in human mammary carcinoma cells. Eur. J. Cell Biol. 2001, 80, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Singhai, R.; Patil, V.W.; Jaiswal, S.R.; Patil, S.D.; Tayade, M.B.; Patil, A.V. E-Cadherin as a diagnostic biomarker in breast cancer. N. Am. J. Med. Sci. 2011, 3, 227. [Google Scholar] [CrossRef]
- Effenberger, K.E.; Johnsen, S.A.; Monroe, D.G.; Spelsberg, T.C.; Westendorf, J.J. Regulation of osteoblastic phenotype and gene expression by hop-derived phytoestrogens. J. Steroid Biochem. Mol. Biol. 2005, 96, 387–399. [Google Scholar] [CrossRef] [PubMed]
- van Duursen, M.B.; Smeets, E.E.; Rijk, J.C.; Nijmeijer, S.M.; van den Berg, M. Phytoestrogens in menopausal supplements induce ER-dependent cell proliferation and overcome breast cancer treatment in an in vitro breast cancer model. Toxicol. Appl. Pharmacol. 2013, 269, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, T.; Fallarini, S.; Gugliesi, F.; Minassi, A.; Appendino, G.; Lombardi, G. Anti-inflammatory and vascularprotective properties of 8-prenylapigenin. Eur. J. Pharmacol. 2009, 620, 120–130. [Google Scholar] [CrossRef]
- Tokalov, S.V.; Henker, Y.; Schwab, P.; Metz, P.; Gutzeit, H.O. Toxicity and cell cycle effects of synthetic 8-prenylnaringenin and derivatives in human cells. Pharmacology 2004, 71, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Negrao, R.; Costa, R.; Duarte, D.; Taveira Gomes, T.; Mendanha, M.; Moura, L.; Vasques, L.; Azevedo, I.; Soares, R. Angiogenesis and inflammation signaling are targets of beer polyphenols on vascular cells. J. Cell. Biochem. 2010, 111, 1270–1279. [Google Scholar] [CrossRef]
- Adeoya-Osiguwa, S.; Markoulaki, S.; Pocock, V.; Milligan, S.; Fraser, L. 17β-Estradiol and environmental estrogens significantly affect mammalian sperm function. Hum. Reprod. 2003, 18, 100–107. [Google Scholar] [CrossRef] [Green Version]
- Fraser, L.R.; Beyret, E.; Milligan, S.R.; Adeoya-Osiguwa, S.A. Effects of estrogenic xenobiotics on human and mouse spermatozoa. Hum. Reprod. 2006, 21, 1184–1193. [Google Scholar] [CrossRef] [Green Version]
- Izzo, G.; Söder, O.; Svechnikov, K. The prenylflavonoid phytoestrogens 8-prenylnaringenin and isoxanthohumol diferentially suppress steroidogenesis in rat Leydig cells in ontogenesis. J. Appl. Toxicol. 2011, 31, 589–594. [Google Scholar] [CrossRef]
- Solak, K.A.; Santos, R.R.; van den Berg, M.; Blaauboer, B.J.; Roelen, B.A.; van Duursen, M.B. Naringenin (NAR) and 8-prenylnaringenin (8-PN) reduce the developmental competence of porcine oocytes in vitro. Reprod. Toxicol. 2014, 49, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Solak, K.A.; Wijnolts, F.M.; Nijmeijer, S.M.; Blaauboer, B.J.; van den Berg, M.; van Duursen, M.B. Excessive levels of diverse phytoestrogens can modulate steroidogenesis and cell migration of KGN human granulosa-derived tumor cells. Toxicol. Rep. 2014, 1, 360–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, E.R.; Clyne, C.; Rubin, G.; Boon, W.C.; Robertson, K.; Britt, K.; Speed, C.; Jones, M. Aromatase—A brief overview. Annu. Rev. Physiol. 2002, 64, 93–127. [Google Scholar] [CrossRef]
- Monteiro, R.; Becker, H.; Azevedo, I.; Calhau, C. Effect of hop (Humulus lupulus L.) flavonoids on aromatase (estrogen synthase) activity. J. Agric. Food Chem. 2006, 54, 2938–2943. [Google Scholar] [CrossRef] [PubMed]
- Corso, A.; Cappiello, M.; Mura, U. From a dull enzyme to something else: Facts and perspectives regarding aldose reductase. Curr. Med. Chem. 2008, 15, 1452–1461. [Google Scholar] [CrossRef]
- Endo, S.; Matsunaga, T.; Nishinaka, T. The Role of AKR1B10 in Physiology and Pathophysiology. Metabolites 2021, 11, 332. [Google Scholar] [CrossRef]
- Seliger, J.M.; Misuri, L.; Maser, E.; Hintzpeter, J. The hop-derived compounds xanthohumol, isoxanthohumol and 8-prenylnaringenin are tight-binding inhibitors of human aldo-keto reductases 1B1 and 1B10. J. Enzym. Inhib. Med. Chem. 2018, 33, 607–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seliger, J.M.; Martin, H.-J.; Maser, E.; Hintzpeter, J. Potent inhibition of human carbonyl reductase 1 (CBR1) by the prenylated chalconoid xanthohumol and its related prenylflavonoids isoxanthohumol and 8-prenylnaringenin. Chem. Biol. Interact. 2019, 305, 156–162. [Google Scholar] [CrossRef]
- Orhan, I.E.; Jedrejek, D.; Senol, F.S.; Salmas, R.E.; Durdagi, S.; Kowalska, I.; Pecio, L.; Oleszek, W. Molecular modeling and in vitro approaches towards cholinesterase inhibitory effect of some natural xanthohumol, naringenin, and acyl phloroglucinol derivatives. Phytomedicine 2018, 42, 25–33. [Google Scholar] [CrossRef]
- Pepper, M.S.; Hazel, S.J.; Hümpel, M.; Schleuning, W.D. 8-prenylnaringenin, a novel phytoestrogen, inhibits angiogenesis in vitro and in vivo. J. Cell. Physiol. 2004, 199, 98–107. [Google Scholar] [CrossRef]
- Costa, R.; Rodrigues, I.; Guardao, L.; Lima, J.Q.; Sousa, E.; Soares, R.; Negrao, R. Modulation of VEGF signaling in a mouse model of diabetes by xanthohumol and 8-prenylnaringenin: Unveiling the angiogenic paradox and metabolism interplay. Mol. Nutr. Food Res. 2017, 61, 1600488. [Google Scholar] [CrossRef] [PubMed]
- Fetalvero, K.M.; Martin, K.A.; Hwa, J. Cardioprotective prostacyclin signaling in vascular smooth muscle. Prostaglandins Other Lipid Mediat. 2007, 82, 109–118. [Google Scholar] [CrossRef]
- Cho, Y.-C.; You, S.-K.; Kim, H.J.; Cho, C.-W.; Lee, I.-S.; Kang, B.Y. Xanthohumol inhibits IL-12 production and reduces chronic allergic contact dermatitis. Int. Immunopharmacol. 2010, 10, 556–561. [Google Scholar] [CrossRef] [PubMed]
- Deiters, B.; Prehm, P. Inhibition of hyaluronan export reduces collagen degradation in interleukin-1 treated cartilage. Arthritis Res. Ther. 2008, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Stracke, D.; Schulz, T.; Prehm, P. Inhibitors of hyaluronan export from hops prevent osteoarthritic reactions. Mol. Nutr. Food Res. 2011, 55, 485–494. [Google Scholar] [CrossRef]
- Dixon, R.A. Phytoestrogens. Annu. Rev. Plant Biol. 2004, 55, 225–261. [Google Scholar] [CrossRef] [Green Version]
- Di Vito, C.; Bertoni, A.; Nalin, M.; Sampietro, S.; Zanfa, M.; Sinigaglia, F. The phytoestrogen 8-prenylnaringenin inhibits agonist-dependent activation of human platelets. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2012, 1820, 1724–1733. [Google Scholar] [CrossRef]
- van Breemen, R.B.; Yuan, Y.; Banuvar, S.; Shulman, L.P.; Qiu, X.; Alvarenga, R.F.R.; Chen, S.N.; Dietz, B.M.; Bolton, J.L.; Pauli, G.F.; et al. Pharmacokinetics of prenylated hop phenols in women following oral administration of a standardized extract of hops. Mol. Nutr. Food Res. 2014, 58, 1962–1969. [Google Scholar] [CrossRef] [Green Version]
- Mukai, R.; Horikawa, H.; Lin, P.-Y.; Tsukumo, N.; Nikawa, T.; Kawamura, T.; Nemoto, H.; Terao, J. 8-Prenylnaringenin promotes recovery from immobilization-induced disuse muscle atrophy through activation of the Akt phosphorylation pathway in mice. Am. J. Physiol. -Regul. Integr. Comp. Physiol. 2016, 311, R1022–R1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukai, R.; Horikawa, H.; Fujikura, Y.; Kawamura, T.; Nemoto, H.; Nikawa, T.; Terao, J. Prevention of disuse muscle atrophy by dietary ingestion of 8-prenylnaringenin in denervated mice. PLoS ONE 2012, 7, e45048. [Google Scholar] [CrossRef] [Green Version]
- Oberbauer, E.; Urmann, C.; Steffenhagen, C.; Bieler, L.; Brunner, D.; Furtner, T.; Humpel, C.; Bäumer, B.; Bandtlow, C.; Couillard-Despres, S.; et al. Chroman-like cyclic prenylflavonoids promote neuronal differentiation and neurite outgrowth and are neuroprotective. J. Nutr. Biochem. 2013, 24, 1953–1962. [Google Scholar] [CrossRef]
- Urmann, C.; Oberbauer, E.; Couillard-Després, S.; Aigner, L.; Riepl, H. Neurodifferentiating potential of 8-prenylnaringenin and related compounds in neural precursor cells and correlation with estrogen-like activity. Planta Med. 2015, 81, 305–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteiro, A.F.M.; Viana, J.D.O.; Nayarisseri, A.; Zondegoumba, E.N.; Mendonça Junior, F.J.B.; Scotti, M.T.; Scotti, L. Computational studies applied to flavonoids against alzheimer’s and parkinson’s diseases. Oxidative Med. Cell. Longev. 2018, 2018, 912765. [Google Scholar] [CrossRef]
- Kim, J.H.; Cho, I.S.; So, Y.K.; Kim, H.-H.; Kim, Y.H. Kushenol A and 8-prenylkaempferol, tyrosinase inhibitors, derived from Sophora flavescens. J. Enzym. Inhib. Med. Chem. 2018, 33, 1048–1054. [Google Scholar] [CrossRef] [Green Version]
- Benkherouf, A.Y.; Soini, S.L.; Stompor, M.; Uusi-Oukari, M. Positive allosteric modulation of native and recombinant GABAA receptors by hops prenylflavonoids. Eur. J. Pharmacol. 2019, 852, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Bagatin, M.C.; Tozatti, C.S.S.; Abiko, L.A.; dos Santos Yamazaki, D.A.; Silva, P.R.A.; Perego, L.M.; Audi, E.A.; Seixas, F.A.V.; Basso, E.A.; de Freitas Gauze, G.; et al. Molecular Docking and Panicolytic Effect of 8-Prenylnaringenin in the Elevated T-Maze. Chem. Pharm. Bull. 2014, 62, 1231–1237. [Google Scholar] [CrossRef] [Green Version]
- Luescher, S.; Urmann, C.; Butterweck, V. Effect of hops derived prenylated phenols on TNF-α induced barrier dysfunction in intestinal epithelial cells. J. Nat. Prod. 2017, 80, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Baum, B.; Georgiou, M. Dynamics of adherens junctions in epithelial establishment, maintenance, and remodeling. J. Cell Biol. 2011, 192, 907–917. [Google Scholar] [CrossRef] [PubMed]
- De Ryck, T.; Van Impe, A.; Vanhoecke, B.W.; Heyerick, A.; Vakaet, L.; De Neve, W.; Müller, D.; Schmidt, M.; Dörr, W.; Bracke, M.E.; et al. 8-prenylnaringenin and tamoxifen inhibit the shedding of irradiated epithelial cells and increase the latency period of radiation-induced oral mucositis. Strahlenther. Und Onkol. 2015, 191, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Bartmańska, A.; Wałecka-Zacharska, E.; Tronina, T.; Popłoński, J.; Sordon, S.; Brzezowska, E.; Bania, J.; Huszcza, E. Antimicrobial properties of spent hops extracts, flavonoids isolated therefrom, and their derivatives. Molecules 2018, 23, 2059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, K.R.; Lyu, X.; Mark, R.; Chen, W.N. Antimicrobial and antioxidant activities of phenolic metabolites from flavonoid-producing yeast: Potential as natural food preservatives. Food Chem. 2019, 270, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Omar, R.M.; Igoli, J.; Gray, A.I.; Ebiloma, G.U.; Clements, C.; Fearnley, J.; Edrada Ebel, R.A.; Zhang, T.; De Koning, H.P.; Watson, D.G.; et al. Chemical characterisation of Nigerian red propolis and its biological activity against Trypanosoma brucei. Phytochem. Anal. 2016, 27, 107–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda, C.L.; Stevens, J.F.; Ivanov, V.; McCall, M.; Frei, B.; Deinzer, M.L.; Buhler, D.R. Antioxidant and prooxidant actions of prenylated and nonprenylated chalcones and flavanones in vitro. J. Agric. Food Chem. 2000, 48, 3876–3884. [Google Scholar] [CrossRef] [PubMed]
- Luís, C.; Costa, R.; Rodrigues, I.; Castela, Â.; Coelho, P.; Guerreiro, S.; Gomes, J.; Reis, C.; Soares, R. Xanthohumol and 8-prenylnaringenin reduce type 2 diabetes–associated oxidative stress by downregulating galectin-3. Porto Biomed. J. 2019, 4, e23. [Google Scholar] [CrossRef] [PubMed]
- Nowotny, K.; Jung, T.; Höhn, A.; Weber, D.; Grune, T. Advanced glycation end products and oxidative stress in type 2 diabetes mellitus. Biomolecules 2015, 5, 194–222. [Google Scholar] [CrossRef] [Green Version]
- Koch, K.; Schulz, G.; Döring, W.; Büchter, C.; Havermann, S.; Mutiso, P.C.; Passreiter, C.; Wätjen, W. Abyssinone V, a prenylated flavonoid isolated from the stem bark of Erythrina melanacantha increases oxidative stress and decreases stress resistance in Caenorhabditis elegans. J. Pharm. Pharmacol. 2019, 71, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Gross-Steinmeyer, K.; Stapleton, P.L.; Tracy, J.H.; Bammler, T.K.; Strom, S.C.; Buhler, D.R.; Eaton, D.L. Modulation of aflatoxin B1–mediated genotoxicity in primary cultures of human hepatocytes by diindolylmethane, curcumin, and xanthohumols. Toxicol. Sci. 2009, 112, 303–310. [Google Scholar] [CrossRef] [Green Version]

| Beneficial | Adverse |
|---|---|
| - Antidiabetic | - Affects sperm with extraordinary potency |
| - Counteracts the metabolic syndrome | - Impairs spindle formation in oocytes and inhibits cumulus expansion |
| - Relieves hot flashes | - Disrupts androgen production in Leydig cells |
| - Inhibits the growth of many cancer cell lines | - Promotes the growth of some cancer cell lines |
| - Induces osteogenesis | - Aggravates inflammatory response topically in skin |
| - Inhibits AKR1B1 and AKR1B10 | - Inhibits transporter proteins (ABCG2, ABCB1/P-gp, ABCC1/MRP1) |
| - Inhibits CBR1 activity | - Alters TSH, T3, GH and IGF-1 serum levels |
| - Induces PGI2 production | - Inhibits aromatase in various cell types |
| - Mitigates LPS-induced effects | - Enhances DNA adduct formation by aflatoxin |
| - Inhibits MRP5 | |
| - Inhibits platelet aggregation | |
| - Accelerates muscle recovery from disuse atrophy | |
| - Maintains barrier tissue integrity | |
| - Has antimicrobial properties | |
| - Promotes neuronal differentiation | |
| - Has high potential for neuroprotective activity |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pohjanvirta, R.; Nasri, A. The Potent Phytoestrogen 8-Prenylnaringenin: A Friend or a Foe? Int. J. Mol. Sci. 2022, 23, 3168. https://doi.org/10.3390/ijms23063168
Pohjanvirta R, Nasri A. The Potent Phytoestrogen 8-Prenylnaringenin: A Friend or a Foe? International Journal of Molecular Sciences. 2022; 23(6):3168. https://doi.org/10.3390/ijms23063168
Chicago/Turabian StylePohjanvirta, Raimo, and Atefeh Nasri. 2022. "The Potent Phytoestrogen 8-Prenylnaringenin: A Friend or a Foe?" International Journal of Molecular Sciences 23, no. 6: 3168. https://doi.org/10.3390/ijms23063168
APA StylePohjanvirta, R., & Nasri, A. (2022). The Potent Phytoestrogen 8-Prenylnaringenin: A Friend or a Foe? International Journal of Molecular Sciences, 23(6), 3168. https://doi.org/10.3390/ijms23063168