
Sera Protein Signatures of Endometrial Cancer Lymph Node Metastases

 ,
,  ,
,
Abstract
:1. Introduction
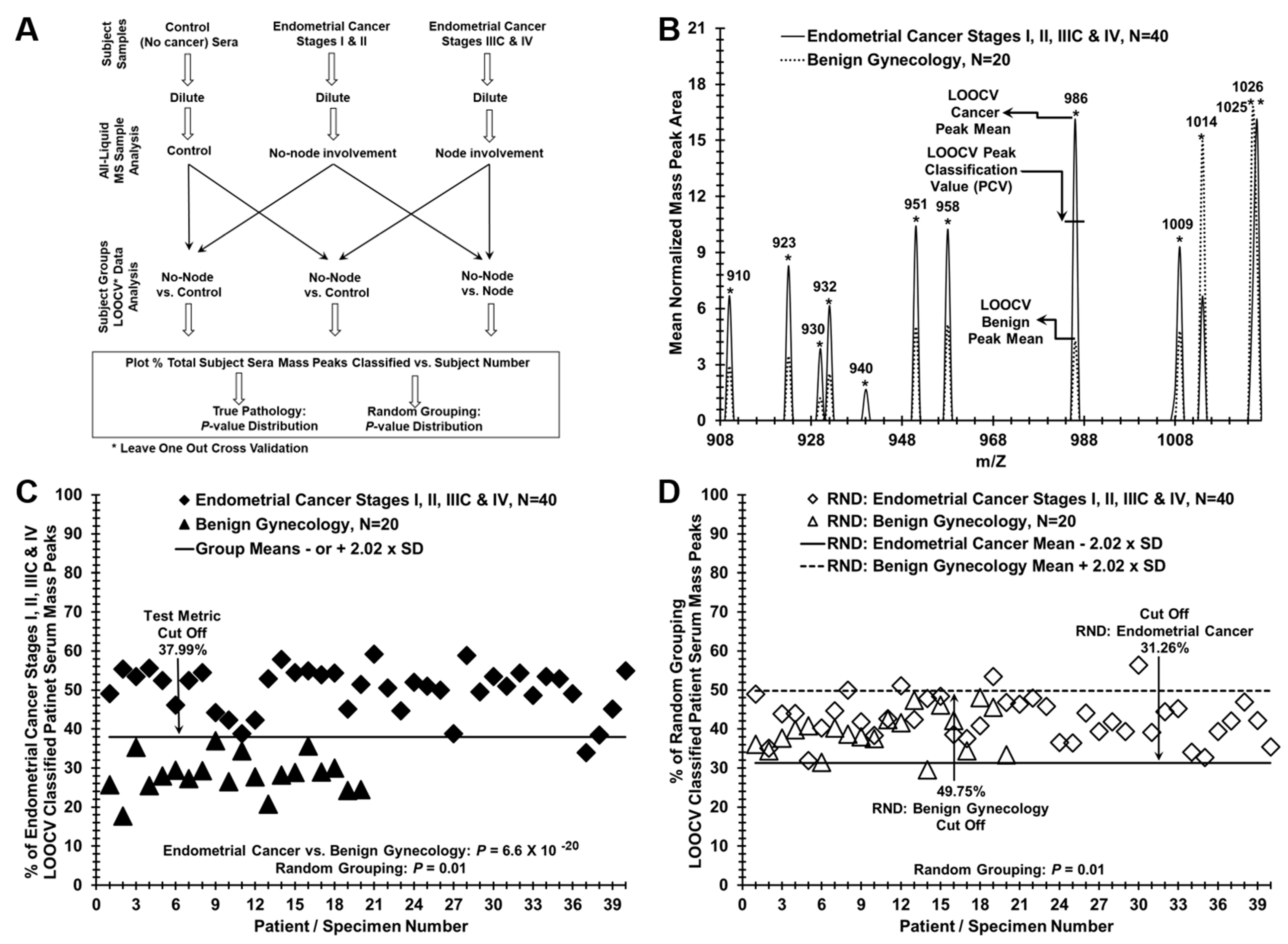
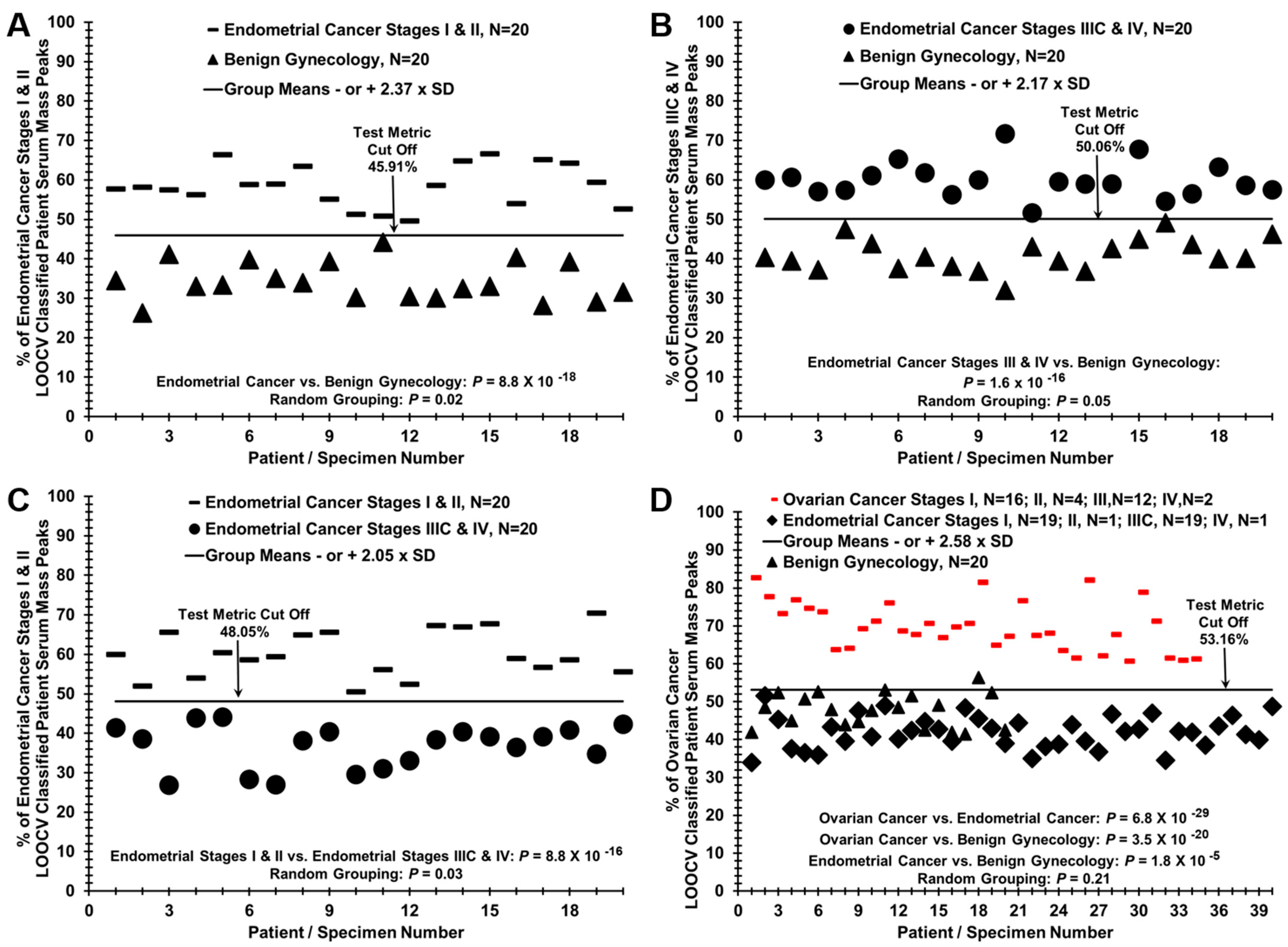
2. Results
3. Discussion
4. Materials and Methods
4.1. Clinical Specimens
4.2. ESI-MS
4.3. LOOCV Mass Peak Analysis and Statistics
4.4. Tandem MS/MS and Bioinformatic Analysis
4.5. Test Metrics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Forman, D.; Bray, F.B.; Brewster, D.H.; Gombe Mbalawa, C.; Kohler, B.; Pineros, M.; Steliarova-Foucher, E.; Swaminathan, R.; Ferlay, J. Cancer Incidence in Five Continents; IARC Scientific Publications No. 164; IARC Scientific Publications: Lyon, France, 2014; Volume X. [Google Scholar]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Buldukoglu, O.C.; Turker, A.; Usubutun, A.; Salman, M.C. Relationship of lymph node status with survival and recurrence among women with endometrial cancer. Int. J. Gynaecol. Obs. 2020, 151, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Press, J.Z.; Gotlieb, W.H. Controversies in the Treatment of Early Stage Endometrial Carcinoma. Obstet. Gynecol. Int. 2012, 2012, 578490. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, K.; Garcia-Sayre, J.; Medeiros, F.; Casabar, J.K.; Machida, H.; Moeini, A.; Roman, L.D. Impact of depth and extent of lymphovascular space invasion on lymph node metastasis and recurrence patterns in endometrial cancer. J. Surg. Oncol. 2015, 112, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Zhai, L.; Zhang, X.; Cui, M.; Wang, J. Sentinel Lymph Node Mapping in Endometrial Cancer: A Comprehensive Review. Front. Oncol. 2021, 11, 701758. [Google Scholar] [CrossRef] [PubMed]
- Åkesson, Å.; Wolmesjö, N.; Adok, C.; Milsom, I.; Dahm-Kähler, P. Lymphadenectomy, obesity and open surgery are associated with surgical complications in endometrial cancer. Eur. J. Surg. Oncol. 2021, 47, 2907–2914. [Google Scholar] [CrossRef]
- Matanes, E.; Eisenberg, N.; Amajoud, Z.; Gupta, V.; Yasmeen, A.; Ismail, S.; Racovitan, F.; Raban, O.; Lau, S.; Salvador, S.; et al. Sentinel Lymph Node Sampling as an Alternative to Lymphadenectomy in Patients with Endometrial Cancer and Obesity. J. Obs. Gynaecol. Can. 2021, 43, 1136–1144.e1131. [Google Scholar] [CrossRef] [PubMed]
- Colombo, N.; Creutzberg, C.; Amant, F.; Bosse, T.; González-Martín, A.; Ledermann, J.; Marth, C.; Nout, R.; Querleu, D.; Mirza, M.R.; et al. ESMO-ESGO-ESTRO Consensus Conference on Endometrial Cancer: Diagnosis, treatment and follow-up. Ann. Oncol. 2016, 27, 16–41. [Google Scholar] [CrossRef]
- Holloway, R.W.; Abu-Rustum, N.R.; Backes, F.J.; Boggess, J.F.; Gotlieb, W.H.; Jeffrey Lowery, W.; Rossi, E.C.; Tanner, E.J.; Wolsky, R.J. Sentinel lymph node mapping and staging in endometrial cancer: A Society of Gynecologic Oncology literature review with consensus recommendations. Gynecol. Oncol. 2017, 146, 405–415. [Google Scholar] [CrossRef]
- Marchocki, Z.; Cusimano, M.C.; Clarfield, L.; Kim, S.R.; Fazelzad, R.; Espin-Garcia, O.; Bouchard-Fortier, G.; Rossi, E.C.; Stewart, K.I.; Soliman, P.T.; et al. Sentinel lymph node biopsy in high-grade endometrial cancer: A systematic review and meta-analysis of performance characteristics. Am. J. Obstet. Gynecol. 2021, 225, 367.e361–367.e339. [Google Scholar] [CrossRef]
- Casarin, J.; Multinu, F.; Abu-Rustum, N.; Cibula, D.; Cliby, W.A.; Ghezzi, F.; Leitao, M.; Konishi, I.; Nam, J.-H.; Querleu, D.; et al. Factors influencing the adoption of the sentinel lymph node technique for endometrial cancer staging: An international survey of gynecologic oncologists. Int. J. Gynecol. Cancer 2019, 29, 60. [Google Scholar] [CrossRef] [PubMed]
- Bogani, G.; Papadia, A.; Buda, A.; Casarin, J.; Di Donato, V.; Gasparri, M.L.; Plotti, F.; Pinelli, C.; Paderno, M.C.; Lopez, S.; et al. Sentinel node mapping vs. sentinel node mapping plus back-up lymphadenectomy in high-risk endometrial cancer patients: Results from a multi-institutional study. Gynecol. Oncol. 2021, 161, 122–129. [Google Scholar] [CrossRef] [PubMed]
- How, J.; Gauthier, C.; Abitbol, J.; Lau, S.; Salvador, S.; Gotlieb, R.; Pelmus, M.; Ferenczy, A.; Probst, S.; Brin, S.; et al. Impact of sentinel lymph node mapping on recurrence patterns in endometrial cancer. Gynecol. Oncol. 2017, 144, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Hocker, J.R.; Bishop, E.A.; Lightfoot, S.A.; Lerner, M.R.; Peyton, M.D.; Brackett, D.J.; Hanas, R.J.; McMeekin, D.S.; Walker, J.L.; Hanas, J.S. Serum profiling to distinguish early- and late-stage ovarian cancer patients from disease-free individuals. Cancer Investig. 2012, 30, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Murali, R.; Soslow, R.A.; Weigelt, B. Classification of endometrial carcinoma: More than two types. Lancet Oncol. 2014, 15, e268–e278. [Google Scholar] [CrossRef]
- Kandoth, C.; Schultz, N.; Cherniack, A.D.; Akbani, R.; Liu, Y.; Shen, H.; Robertson, A.G.; Pashtan, I.; Shen, R.; Benz, C.C.; et al. Integrated genomic characterization of endometrial carcinoma. Nature 2013, 497, 67–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kent, C.N.; Guttilla Reed, I.K. Regulation of epithelial-mesenchymal transition in endometrial cancer: Connecting PI3K, estrogen signaling, and microRNAs. Clin. Transl. Oncol. 2016, 18, 1056–1061. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.P.; Goodwin, J.E. Cancer Biology and Prevention in Diabetes. Cells 2020, 9, 1380. [Google Scholar] [CrossRef] [PubMed]
- Viedma-Rodriguez, R.; Martinez-Hernandez, M.G.; Martinez-Torres, D.I.; Baiza-Gutman, L.A. Epithelial Mesenchymal Transition and Progression of Breast Cancer Promoted by Diabetes Mellitus in Mice Are Associated with Increased Expression of Glycolytic and Proteolytic Enzymes. Horm. Cancer 2020, 11, 170–181. [Google Scholar] [CrossRef] [PubMed]
- Parsons, R. Discovery of the PTEN Tumor Suppressor and Its Connection to the PI3K and AKT Oncogenes. Cold Spring Harb. Perspect. Med. 2020, 10, a036129. [Google Scholar] [CrossRef] [PubMed]
- Papa, A.; Pandolfi, P.P. The PTEN-PI3K Axis in Cancer. Biomolecules 2019, 9, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ijuin, T. Phosphoinositide phosphatases in cancer cell dynamics-Beyond PI3K and PTEN. Semin. Cancer Biol. 2019, 59, 50–65. [Google Scholar] [CrossRef] [PubMed]
- Westin, S.N.; Ju, Z.; Broaddus, R.R.; Krakstad, C.; Li, J.; Pal, N.; Lu, K.H.; Coleman, R.L.; Hennessy, B.T.; Klempner, S.J.; et al. PTEN loss is a context-dependent outcome determinant in obese and non-obese endometrioid endometrial cancer patients. Mol. Oncol. 2015, 9, 1694–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyo, S.; Nakayama, K. Endometrial Cancer as a Metabolic Disease with Dysregulated PI3K Signaling: Shedding Light on Novel Therapeutic Strategies. Int. J. Mol. Sci. 2020, 21, 6073. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-Y.; Xiao, R.; Li, C.-P.; Huangfu, J.; Mao, J.-F. Increased plasma levels of FABP4 and PTEN is associated with more severe insulin resistance in women with gestational diabetes mellitus. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2015, 21, 426–431. [Google Scholar] [CrossRef] [Green Version]
- Benchoula, K.; Parhar, I.S.; Wong, E.H. The crosstalk of hedgehog, PI3K and Wnt pathways in diabetes. Arch. Biochem. Biophys. 2021, 698, 108743. [Google Scholar] [CrossRef] [PubMed]
- Cerniglia, G.J.; Dey, S.; Gallagher-Colombo, S.M.; Daurio, N.A.; Tuttle, S.; Busch, T.M.; Lin, A.; Sun, R.; Esipova, T.V.; Vinogradov, S.A.; et al. The PI3K/Akt Pathway Regulates Oxygen Metabolism via Pyruvate Dehydrogenase (PDH)-E1α Phosphorylation. Mol. Cancer Ther. 2015, 14, 1928–1938. [Google Scholar] [CrossRef] [Green Version]
- Ghigo, A.; Laffargue, M.; Li, M.; Hirsch, E. PI3K and Calcium Signaling in Cardiovascular Disease. Circ. Res. 2017, 121, 282–292. [Google Scholar] [CrossRef]
- Xu, Y.; Li, N.; Xiang, R.; Sun, P. Emerging roles of the p38 MAPK and PI3K/AKT/mTOR pathways in oncogene-induced senescence. Trends Biochem. Sci. 2014, 39, 268–276. [Google Scholar] [CrossRef] [Green Version]
- Dabizzi, S.; Noci, I.; Borri, P.; Borrani, E.; Giachi, M.; Balzi, M.; Taddei, G.L.; Marchionni, M.; Scarselli, G.F.; Arcangeli, A. Luteinizing hormone increases human endometrial cancer cells invasiveness through activation of protein kinase A. Cancer Res. 2003, 63, 4281–4286. [Google Scholar]
- Law, N.C.; White, M.F.; Hunzicker-Dunn, M.E. G protein-coupled receptors (GPCRs) That Signal via Protein Kinase A (PKA) Cross-talk at Insulin Receptor Substrate 1 (IRS1) to Activate the phosphatidylinositol 3-kinase (PI3K)/AKT Pathway. J. Biol. Chem. 2016, 291, 27160–27169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Predescu, D.-V.; Crețoiu, S.M.; Crețoiu, D.; Pavelescu, L.A.; Suciu, N.; Radu, B.M.; Voinea, S.-C. G Protein-Coupled Receptors (GPCRs)-Mediated Calcium Signaling in Ovarian Cancer: Focus on GPCRs activated by Neurotransmitters and Inflammation-Associated Molecules. Int. J. Mol. Sci. 2019, 20, 5568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Hu, J.; Yu, T.; You, S.; Zhang, Y.; Hu, L. miR-27b-3p/MARCH7 regulates invasion and metastasis of endometrial cancer cells through Snail-mediated pathway. Acta Biochim. Biophys. Sin. 2019, 51, 492–500. [Google Scholar] [CrossRef] [PubMed]
- Cherubini, A.; Taddei, G.L.; Crociani, O.; Paglierani, M.; Buccoliero, A.M.; Fontana, L.; Noci, I.; Borri, P.; Borrani, E.; Giachi, M.; et al. HERG potassium channels are more frequently expressed in human endometrial cancer as compared to non-cancerous endometrium. Br. J. Cancer 2000, 83, 1722–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, A.J.; Susil, B.; Jobling, T.W.; Oehler, M.K. Endometrial cancer. Cell Tissue Res. 2005, 322, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Rolitsky, C.D.; Theil, K.S.; McGaughy, V.R.; Copeland, L.J.; Niemann, T.H. HER-2/neu amplification and overexpression in endometrial carcinoma. Int. J. Gynecol. Pathol. Off. J. Int. Soc. Gynecol. Pathol. 1999, 18, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Benevolo, M.; Vocaturo, A.; Novelli, F.; Mariani, L.; Vocaturo, G.; Cianciulli, A.M.; Marandino, F.; Perrone-Donnorso, R.; Giannarelli, D.; Natali, P.G.; et al. Prognostic value of HER2 and progesterone receptor expression in endometrial carcinoma with positive peritoneal washing. Anticancer Res. 2007, 27, 2839–2844. [Google Scholar]
- Srijaipracharoen, S.; Tangjitgamol, S.; Tanvanich, S.; Manusirivithaya, S.; Khunnarong, J.; Thavaramara, T.; Leelahakorn, S.; Pataradool, K. Expression of ER, PR, and Her-2/neu in endometrial cancer: A clinicopathological study. Asian Pac. J. Cancer Prev. APJCP 2010, 11, 215–220. [Google Scholar] [PubMed]
- Kalogiannidis, I.; Petousis, S.; Bobos, M.; Margioula-Siarkou, C.; Topalidou, M.; Papanikolaou, A.; Vergote, I.; Agorastos, T. HER-2/neu is an independent prognostic factor in type I endometrial adenocarcinoma. Arch. Gynecol. Obstet. 2014, 290, 1231–1237. [Google Scholar] [CrossRef]
- Jongen, V.H.; Briët, J.M.; de Jong, R.A.; Joppe, E.; ten Hoor, K.A.; Boezen, H.M.; Evans, D.B.; Hollema, H.; van der Zee, A.G.; Nijman, H.W. Aromatase, cyclooxygenase 2, HER-2/neu, and p53 as prognostic factors in endometrioid endometrial cancer. Int. J. Gynecol. Cancer 2009, 19, 670–676. [Google Scholar] [CrossRef]
- Gai, Q.Z.; Lv, Y.B.; Li, G.Y.; Zhang, D.Q.; Gao, Z.; Fang, X.H. Value of metabolic parameters of primary lesions examined by 18F-FDG PET/CT for endometrial cancer in preoperative evaluation. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 2493–2502. [Google Scholar] [CrossRef] [PubMed]
- Buchynska, L.G.; Brieieva, O.V.; Iurchenko, N.P. Assessment of HER-2/neu, c-MYC and CCNE1 gene copy number variations and protein expression in endometrial carcinomas. Exp. Oncol. 2019, 41, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Valdez, I.A.; Dirice, E.; Gupta, M.K.; Shirakawa, J.; Teo, A.K.K.; Kulkarni, R.N. Proinflammatory Cytokines Induce Endocrine Differentiation in Pancreatic Ductal Cells via STAT3-Dependent NGN3 Activation. Cell Rep. 2016, 15, 460–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wegehaupt, O.; Muckenhaupt, T.; Johnson, M.B.; Schwab, K.O.; Speckmann, C. Ruxolitinib Controls Lymphoproliferation and Diabetes in a STAT3-GOF Patient. J. Clin. Immunol. 2020, 40, 1207–1210. [Google Scholar] [CrossRef] [PubMed]
- Warshauer, J.T.; Belk, J.A.; Chan, A.Y.; Wang, J.; Gupta, A.R.; Shi, Q.; Skartsis, N.; Peng, Y.; Phipps, J.D.; Acenas, D.; et al. A human mutation in STAT3 promotes type 1 diabetes through a defect in CD8+ T cell tolerance. J. Exp. Med. 2021, 218, e20210759. [Google Scholar] [CrossRef] [PubMed]
- Wallbillich, J.J.; Josyula, S.; Saini, U.; Zingarelli, R.A.; Dorayappan, K.D.P.; Riley, M.K.; Wanner, R.A.; Cohn, D.E.; Selvendiran, K. High Glucose-Mediated STAT3 Activation in Endometrial Cancer Is Inhibited by Metformin: Therapeutic Implications for Endometrial Cancer. PLoS ONE 2017, 12, e0170318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Zee, M.; Sacchetti, A.; Cansoy, M.; Joosten, R.; Teeuwssen, M.; Heijmans-Antonissen, C.; Ewing-Graham, P.C.; Burger, C.W.; Blok, L.J.; Fodde, R. IL6/JAK1/STAT3 Signaling Blockade in Endometrial Cancer Affects the ALDHhi/CD126+ Stem-like Component and Reduces Tumor Burden. Cancer Res. 2015, 75, 3608–3622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieswandt, B.; Bergmeier, W.; Eckly, A.; Schulte, V.; Ohlmann, P.; Cazenave, J.-P.; Zirngibl, H.; Offermanns, S.; Gachet, C. Evidence for cross-talk between glycoprotein VI and Gi-coupled receptors during collagen-induced platelet aggregation. Blood 2001, 97, 3829–3835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muzykiewicz, K.P.; Iwanska, E.; Janeczek, M.; Glanowska, I.; Karolewski, K.; Blecharz, P. The analysis of the prognostic value of the neutrophil/ lymphocyte ratio and the platelet/lymphocyte ratio among advanced endometrial cancer patients. Ginekol. Pol. 2021, 92, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Cong, R.; Kong, F.; Ma, J.; Li, Q.; Wu, Q.; Ma, X. Combination of preoperative neutrophil-lymphocyte ratio, platelet-lymphocyte ratio and monocyte-lymphocyte ratio: A superior prognostic factor of endometrial cancer. BMC Cancer 2020, 20, 464. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wu, Q.; Zhang, Y.; Li, Q.; Ma, J.; Kong, F.; Ma, X. Nomograms based on the novel platelet index score predict postoperative prognosis in endometrial cancer. Gynecol. Oncol. 2020, 158, 689–697. [Google Scholar] [CrossRef]
- Selen, S.; Kilic, F.; Kimyon Comert, G.; Unsal, M.; Kilic, C.; Karalok, A.; Turkmen, O.; Turan, T. Can preoperative inflammatory markers differentiate endometrial cancer from complex atypical hyperplasia/endometrial intraepithelial neoplasia? J. Obstet. Gynaecol. Res. 2020, 46, 1148–1156. [Google Scholar] [CrossRef] [PubMed]
- Fidler, T.P.; Campbell, R.A.; Funari, T.; Dunne, N.; Balderas Angeles, E.; Middleton, E.A.; Chaudhuri, D.; Weyrich, A.S.; Abel, E.D. Deletion of GLUT1 and GLUT3 Reveals Multiple Roles for Glucose Metabolism in Platelet and Megakaryocyte Function. Cell Rep. 2017, 20, 881–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Androutsopoulos, G.; Adonakis, G.; Terzakis, E.; Geropoulou, E.; Decavalas, G. Endometrial cancer in a patient with rheumatoid arthritis. Eur. J. Gynaecol. Oncol. 2015, 36, 91–93. [Google Scholar] [PubMed]
- Abdalla, H.; Bagchi, A.; Bandagi, S. Rheumatoid Arthritis as a Therapeutic Challenge in a Patient with Lynch Syndrome. Am. J. Case Rep. 2015, 16, 390–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, A.E.; Alali, A.; Starr, L.J.; Shah, N.; Beavis, A.; Pereira, E.M.; Lindsay, M.E.; Klugman, S. Gain-of-function pathogenic variants in SMAD4 are associated with neoplasia in Myhre syndrome. Am. J. Med. Genet. Part A 2020, 182, 328–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.H.; Park, J.K.; Lee, Y.J.; Yang, J.A.; Lee, E.Y.; Song, Y.W.; Lee, E.B. Comparison of cancer incidence among patients with rheumatic disease: A retrospective cohort study. Arthritis Res. Ther. 2014, 16, 428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hocker, J.R.; Postier, R.G.; Li, M.; Lerner, M.R.; Lightfoot, S.A.; Peyton, M.D.; Deb, S.J.; Baker, C.M.; Williams, T.L.; Hanas, R.J.; et al. Discriminating patients with early-stage pancreatic cancer or chronic pancreatitis using serum electrospray mass profiling. Cancer Lett. 2015, 359, 314–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hocker, J.R.; Mohammed, A.; Aston, C.E.; Brewer, M.; Lightfoot, S.A.; Rao, C.V.; Hanas, J.S. Mass profiling of serum to distinguish mice with pancreatic cancer induced by a transgenic Kras mutation. Int. J. Cancer 2013, 133, 2662–2671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hocker, J.R.; Peyton, M.D.; Lerner, M.R.; Mitchell, S.L.; Lightfoot, S.A.; Lander, T.J.; Bates-Albers, L.M.; Vu, N.T.; Hanas, R.J.; Kupiec, T.C.; et al. Serum discrimination of early-stage lung cancer patients using electrospray-ionization mass spectrometry. Lung Cancer 2011, 74, 206–211. [Google Scholar] [CrossRef]
- Dittmar, G.A.; Wilkinson, C.R.; Jedrzejewski, P.T.; Finley, D. Role of a ubiquitin-like modification in polarized morphogenesis. Science 2002, 295, 2442–2446. [Google Scholar] [CrossRef] [PubMed]
- Altman, D.G.; Bland, J.M. Diagnostic tests. 1: Sensitivity and specificity. BMJ 1994, 308, 1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altman, D.G.; Bland, J.M. Statistics Notes: Diagnostic tests 2: Predictive values. BMJ 1994, 309, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Benign | Stage I or II | Stage IIIC or IVB | p-Value * |
|---|---|---|---|---|
| Age, y, median (range) | 58 (50–64) | 57 (52–63) | 62 (32–77) | 0.43 |
| BMI, mean (SD) | 35.4 (7.9) | 35.3 (7.5) | 34.7 (6.5) | 0.95 |
| Race: | 0.56 | |||
| White | 10 | 17 | 15 | |
| Black | 1 | 2 | 4 | |
| American Indian | 0 | 1 | 0 | |
| Asian | 0 | 0 | 1 | |
| Unknown/Other | 9 | 0 | 0 | |
| Endometrioid Histology: | ||||
| No | N/A | |||
| Yes | N/A | |||
| Tobacco Use: | ||||
| No | 15 | 13 | 13 | >0.99 |
| Yes | 1 | 1 | 2 | |
| Second-hand | 3 | 6 | 4 | |
| Unknown | 1 | 0 | 0 | |
| Alcohol Use: | 0.78 | |||
| No | 14 | 17 | 16 | |
| Yes | 5 | 3 | 3 | |
| Unknown | 1 | 0 | 0 | |
| NSAIDs Use: | >0.99 | |||
| No | 17 | 14 | 12 | |
| Yes | 3 | 6 | 7 | |
| Aspirin Use: | >0.99 | |||
| No | 18 | 16 | 16 | |
| Yes | 2 | 4 | 3 | |
| Metformin Use: | >0.99 | |||
| No | 20 | 19 | 18 | |
| Yes | 0 | 1 | 1 | |
| Insulin Use: | >0.99 | |||
| No | 20 | 20 | 18 | |
| Yes | 0 | 0 | 1 | |
| Type 2 Diabetes: | >0.99 | |||
| No | 19 | 17 | 16 | |
| Yes | 1 | 3 | 3 | |
| Hypertension: | >0.99 | |||
| No | 10 | 13 | 10 | |
| Yes | 10 | 7 | 9 | |
| Cardiovascular Disease: | >0.99 | |||
| No | 19 | 19 | 19 | |
| Yes | 1 | 1 | 0 | |
| Arthritis: | >0.99 | |||
| No | 19 | 18 | 14 | |
| Yes | 1 | 2 | 5 |
| Group Comparison | Pathway | z-Score | p-Value | Molecules * |
|---|---|---|---|---|
| Stages I and II vs. Benign | Hepatic Fibrosis Signaling | −2.646 | 0.019 | FGFR1, GLI1, ITGB2, NOX1, SOS2, TGFBR2, TTN, WNT6 |
| Regulation of EMT by Growth Factors | −2.236 | 0.018 | EGF, FGFR1, GSC, SOS2, TGFBR2 | |
| HER−2 Signaling in Breast Cancer | −1.342 | 0.019 | EGF, ITGB2, MT-CO1, NRG1, SOS2 | |
| Neuroinflammation Signaling | −1.342 | 0.040 | CASP8, ELP1, GRIN3B, NOX1, PPP3R1, TGFBR2 | |
| GP6 Signaling | −1 | 0.018 | COL4A3, LAMA2, LAMA5, NOX1 | |
| Regulation of EMT in Development | −1 | 0.005 | GLI1, GSC, JAG2, WNT6 | |
| STAT3 | −1 | 0.022 | EGF, FGFR1, INSR, TGFBR2 | |
| Th2 | 1.0 | 0.0005 | ITGB2, JAG2, NOTCH3, NOTCH4, TGFBR2 | |
| PTEN Signaling | 0.447 | 0.0002 | ELP1, FGFR1, INSR, ITGB2, MAGI2, SOS2, TGFBR2 | |
| PKA Signaling | 0.447 | 0.041 | AKAP13, PPP3R1, PTPN22, PTPRS, RYR2, TGFBR2, TTN | |
| Stages IIIC and IV vs. Benign | Cardiac Hypertrophy Signaling | 2.236 | 0.01 | CACNA1A, CACNA1E, KRAS, NFATC4, PLCG1 |
| Synaptic Long-term Depression | 2.236 | 0.003 | CACNA1A, CACNA1E, GRIA4, KRAS, PLCG1 | |
| Role of NFAT in Cardiac Hypertrophy | 2.236 | 0.006 | CACNA1A, CACNA1E, KRAS, NFATC4, PLCG1 | |
| ER Signaling | 2.236 | 0.005 | CACNA1A, CACNA1E, KRAS, MT-CYB, MT-ND1, MT-ND5, PLCG1 | |
| Senescence | 2.236 | 0.019 | CACNA1A, CACNA1E, EP400, KRAS, NFATC4 | |
| Oxidative Phosphorylation | 2 | 0.003 | MT-CO2, MT-CYB, MT-ND1, MT-ND5 | |
| G Beta Gamma Signaling | 2 | 0.005 | CACNA1A, CACNA1E, KRAS, PLCG1 | |
| Endocannabinoid Neuronal Synapse | 2 | 0.007 | CACNA1A, CACNA1E, GRIA4, PLCG1 | |
| Ca2+ Signaling | 2 | 0.026 | CACNA1A, CACNA1E, GRIA4, NFATC4 | |
| PKA Signaling | 1 | 0.018 | FLNB, NFATC4, PDE3B, PLCG1, PTPN5, TTN | |
| PI3K Signaling in B Lymphocytes | 1 | 0.007 | CBL, KRAS, NFATC4, PLCG1 | |
| Integrin Signaling | 1 | 0.025 | KRAS, PLCG1, TLN2, TTN | |
| Synaptogenesis Signaling | 0.447 | 0.023 | CDH23, GRIA4, KRAS, PLCG1, RELN | |
| Osteoarthritis | 0.447 | 0.0006 | FN1, ITGB6, LRP1, NOTCH1, PPARD, TNFRSF1B | |
| Stages IIIC and IV vs. Stages II and III | Osteoarthritis | 0.447 | 0.0006 | FN1, ITGB6, LRP1, NOTCH1, PPARD, TNFRSF1B |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benbrook, D.M.; Hocker, J.R.S.; Moxley, K.M.; Hanas, J.S. Sera Protein Signatures of Endometrial Cancer Lymph Node Metastases. Int. J. Mol. Sci. 2022, 23, 3277. https://doi.org/10.3390/ijms23063277
Benbrook DM, Hocker JRS, Moxley KM, Hanas JS. Sera Protein Signatures of Endometrial Cancer Lymph Node Metastases. International Journal of Molecular Sciences. 2022; 23(6):3277. https://doi.org/10.3390/ijms23063277
Chicago/Turabian StyleBenbrook, Doris Mangiaracina, James Randolph Sanders Hocker, Katherine Marie Moxley, and Jay S. Hanas. 2022. "Sera Protein Signatures of Endometrial Cancer Lymph Node Metastases" International Journal of Molecular Sciences 23, no. 6: 3277. https://doi.org/10.3390/ijms23063277
APA StyleBenbrook, D. M., Hocker, J. R. S., Moxley, K. M., & Hanas, J. S. (2022). Sera Protein Signatures of Endometrial Cancer Lymph Node Metastases. International Journal of Molecular Sciences, 23(6), 3277. https://doi.org/10.3390/ijms23063277