
Polarization of Microglia and Its Therapeutic Potential in Sepsis


 and
and
Abstract
:1. Introduction
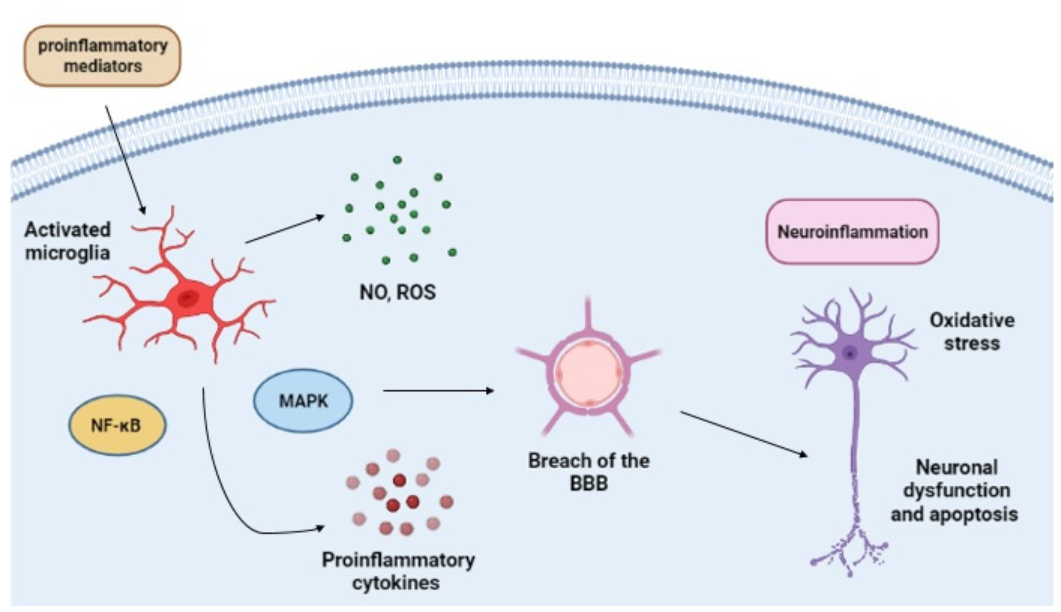
2. Microglia and Their Role in the CNS
3. PPARγ-Dependent Anti-Inflammatory and Pro-Resolutive Mechanisms
4. Critical View of the Actual Scenario
5. Final Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rhodes, A.; Evans, L.E.; Alhazzani, W.; Levy, M.M.; Antonelli, M.; Ferrer, R.; Kumar, A.; Sevransky, J.E.; Sprung, C.L.; Nunnally, M.E.; et al. Surviving Sepsis Campaign: International Guidelines for Management of Sepsis and Septic Shock: 2016. Intensive Care Med. 2017, 43, 304–377. [Google Scholar] [CrossRef] [PubMed]
- Tay, M.Z.; Poh, C.M.; Rénia, L.; Macary, P.A.; Ng, L.F.P. The trinity of COVID-19: Immunity, inflammation and intervention. Nat. Rev. Immunol. 2020, 20, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Gyawali, B.; Ramakrishna, K.; Dhamoon, A.S. Sepsis: The evolution in definition, pathophysiology, and management. SAGE Open Med. 2019, 7, 205031211983504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarczak, D.; Kluge, S.; Nierhaus, A. Sepsis—Pathophysiology and Therapeutic Concepts. Front. Med. 2021, 8, 8. [Google Scholar] [CrossRef]
- Calsavara, A.J.C.; Costa, P.A.; Nobre, V.; Teixeira, A.L. Factors Associated with Short and Long Term Cognitive Changes in Patients with Sepsis. Sci. Rep. 2018, 8, 4509. [Google Scholar] [CrossRef]
- Rudd, K.E.; Johnson, S.C.; Agesa, K.M.; Shackelford, K.A.; Tsoi, D.; Kievlan, D.R.; Colombara, D.V.; Ikuta, K.S.; Kissoon, N.; Finfer, S.; et al. Global, regional, and national sepsis incidence and mortality, 1990–2017: Analysis for the Global Burden of Disease Study. Lancet 2020, 395, 200–211. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.; Zhou, F.; Wang, Y.; Fan, G.; Cao, B. Respiratory viral sepsis: Epidemiology, pathophysiology, diagnosis and treatment. Eur. Respir. Rev. 2020, 29, 200038. [Google Scholar] [CrossRef]
- Meneses, G.; Cárdenas, G.; Espinosa, A.; Rassy, D.; Pérez-Osorio, I.N.; Bárcena, B.; Fleury, A.; Besedovsky, H.; Fragoso, G.; Sciutto, E. Sepsis: Developing new alternatives to reduce neuroinflammation and attenuate brain injury. Ann. N. Y. Acad. Sci. 2019, 1437, 43–56. [Google Scholar] [CrossRef]
- Ren, C.; Yao, R.-Q.; Zhang, H.; Feng, Y.-W.; Yao, Y.-M. Sepsis-associated encephalopathy: A vicious cycle of immunosuppression. J. Neuroinflamm. 2020, 17, 14. [Google Scholar] [CrossRef]
- Arcuri, C.; Mecca, C.; Bianchi, R.; Giambanco, I.; Donato, R. The Pathophysiological Role of Microglia in Dynamic Surveillance, Phagocytosis and Structural Remodeling of the Developing CNS. Front. Mol. Neurosci. 2017, 10, 191. [Google Scholar] [CrossRef] [Green Version]
- Czempik, P.F.; Pluta, M.P.; Krzych, Ł.J. Sepsis-Associated Brain Dysfunction: A Review of Current Literature. Int. J. Environ. Res. Public Health 2020, 17, 5852. [Google Scholar] [CrossRef] [PubMed]
- Adam, N.; Kandelman, S.; Mantz, J.; Chrétien, F.; Sharshar, T. Sepsis-induced brain dysfunction. Expert Rev. Anti-Infect. Ther. 2013, 11, 211–221. [Google Scholar] [CrossRef]
- Caraballo, C.; Jaimes, F. Organ Dysfunction in Sepsis: An Ominous Trajectory from Infection to Death. Yale J. Biol. Med. 2019, 92, 629–640. [Google Scholar]
- Mostel, Z.; Perl, A.; Marck, M.; Mehdi, S.F.; Lowell, B.; Bathija, S.; Santosh, R.; Pavlov, V.A.; Chavan, S.S.; Roth, J. Post-sepsis syndrome—An evolving entity that afflicts survivors of sepsis. Mol. Med. 2020, 26, 1–14. [Google Scholar] [CrossRef] [Green Version]
- van der Slikke, E.C.; An, A.Y.; Hancock, R.E.; Bouma, H.R. Exploring the pathophysiology of post-sepsis syndrome to identify therapeutic opportunities. EBioMedicine 2020, 61, 103044. [Google Scholar] [CrossRef]
- Prescott, H.; Angus, D.C. Enhancing Recovery from Sepsis: A Review. JAMA 2018, 319, 62–75. [Google Scholar] [CrossRef]
- Ginhoux, F.; Garel, S. The mysterious origins of microglia. Nat. Neurosci. 2018, 21, 897–899. [Google Scholar] [CrossRef]
- Bennett, M.L.; Bennett, F. The influence of environment and origin on brain resident macrophages and implications for therapy. Nat. Neurosci. 2020, 23, 157–166. [Google Scholar] [CrossRef]
- Peng, J.; Wang, K.; Xiang, W.; Li, Y.; Hao, Y.; Guan, Y. Rosiglitazone polarizes microglia and protects against pilocarpine-induced status epilepticus. CNS Neurosci. Ther. 2019, 25, 1363–1372. [Google Scholar] [CrossRef]
- Wen, L.; You, W.; Wang, H.; Meng, Y.; Feng, J.; Yang, X. Polarization of Microglia to the M2 Phenotype in a Peroxisome Proliferator-Activated Receptor Gamma–Dependent Manner Attenuates Axonal Injury Induced by Traumatic Brain Injury in Mice. J. Neurotrauma 2018, 35, 2330–2340. [Google Scholar] [CrossRef]
- Gogoleva, V.S.; Drutskaya, M.S.; Atretkhany, K.S.-N. The Role of Microglia in the Homeostasis of the Central Nervous System and Neuroinflammation. Mol. Biol. 2019, 53, 696–703. [Google Scholar] [CrossRef]
- Hong, S.; Dissing-Olesen, L.; Stevens, B. New insights on the role of microglia in synaptic pruning in health and disease. Curr. Opin. Neurobiol. 2016, 36, 128–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bar, E.; Barak, B. Microglia roles in synaptic plasticity and myelination in homeostatic conditions and neurodevelopmental disorders. Glia 2019, 67, 2125–2141. [Google Scholar] [CrossRef] [PubMed]
- Zrzavy, T.; Höftberger, R.; Berger, T.; Rauschka, H.; Butovsky, O.; Weiner, H.; Lassmann, H. Pro-inflammatory activation of microglia in the brain of patients with sepsis. Neuropathol. Appl. Neurobiol. 2019, 45, 278–290. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Le, W. Differential Roles of M1 and M2 Microglia in Neurodegenerative Diseases. Mol. Neurobiol. 2016, 53, 1181–1194. [Google Scholar] [CrossRef]
- Li, Y.; Yin, L.; Fan, Z.; Su, B.; Chen, Y.; Ma, Y.; Zhong, Y.; Hou, W.; Fang, Z.; Zhang, X. Microglia: A Potential Therapeutic Target for Sepsis-Associated Encephalopathy and Sepsis-Associated Chronic Pain. Front. Pharmacol. 2020, 11, 600421. [Google Scholar] [CrossRef]
- Wang, J.; Xing, H.; Wan, L.; Jiang, X.; Wang, C.; Wu, Y. Treatment targets for M2 microglia polarization in ischemic stroke. Biomed. Pharmacother. 2018, 105, 518–525. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, J.; You, Z. Switching of the Microglial Activation Phenotype Is a Possible Treatment for Depression Disorder. Front. Cell. Neurosci. 2018, 12, 306. [Google Scholar] [CrossRef]
- Haruwaka, K.; Ikegami, A.; Tachibana, Y.; Ohno, N.; Konishi, H.; Hashimoto, A.; Matsumoto, M.; Kato, D.; Ono, R.; Kiyama, H.; et al. Dual microglia effects on blood brain barrier permeability induced by systemic inflammation. Nat. Commun. 2019, 10, 5816. [Google Scholar] [CrossRef] [Green Version]
- Bachiller, S.; Jiménez-Ferrer, I.; Paulus, A.; Yang, Y.; Swanberg, M.; Deierborg, T.; Boza-Serrano, A. Microglia in Neurological Diseases: A Road Map to Brain-Disease Dependent-Inflammatory Response. Front. Cell. Neurosci. 2018, 12, 488. [Google Scholar] [CrossRef] [Green Version]
- Gomes-Leal, W. Why microglia kill neurons after neural disorders? The friendly fire hypothesis. Neural Regen. Res. 2019, 14, 1499–1502. [Google Scholar] [CrossRef]
- Honjoh, K.; Nakajima, H.; Hirai, T.; Watanabe, S.; Matsumine, A. Relationship of Inflammatory Cytokines from M1-Type Microglia/Macrophages at the Injured Site and Lumbar Enlargement with Neuropathic Pain After Spinal Cord Injury in the CCL21 Knockout (plt) Mouse. Front. Cell. Neurosci. 2019, 13, 525. [Google Scholar] [CrossRef] [Green Version]
- Lan, X.; Han, X.; Li, Q.; Yang, Q.-W.; Wang, J. Modulators of microglial activation and polarization after intracerebral haemorrhage. Nat. Rev. Neurol. 2017, 13, 420–433. [Google Scholar] [CrossRef] [Green Version]
- Brás, J.P.; Bravo, J.; Freitas, J.; Barbosa, M.A.; Santos, S.G.; Summavielle, T.; Almeida, M.I. TNF-alpha-induced microglia activation requires miR-342: Impact on NF-kB signaling and neurotoxicity. Cell Death Dis. 2020, 11, 415. [Google Scholar] [CrossRef]
- Belov Kirdajova, D.; Kriska, J.; Tureckova, J.; Anderova, M. Ischemia-Triggered Glutamate Excitotoxicity from the Perspective of Glial Cells. Front. Cell. Neurosci. 2020, 14, 51. [Google Scholar] [CrossRef] [Green Version]
- Mendiola, A.; Cardona, A.E. The IL-1β phenomena in neuroinflammatory diseases. J. Neural Transm. 2017, 125, 781–795. [Google Scholar] [CrossRef]
- Jiang, C.T.; Wu, W.F.; Deng, Y.H.; Ge, J.W. Modulators of microglia activation and polarization in ischemic stroke (Review). Mol. Med. Rep. 2020, 21, 2006–2018. [Google Scholar] [CrossRef] [Green Version]
- Moraes, C.A.; Santos, G.; Spohr, T.C.L.D.S.E.; D’Avila, J.; Lima, F.R.S.; Benjamim, C.; Bozza, F.A.; Gomes, F.C.A. Activated Microglia-Induced Deficits in Excitatory Synapses Through IL-1β: Implications for Cognitive Impairment in Sepsis. Mol. Neurobiol. 2015, 52, 653–663. [Google Scholar] [CrossRef]
- Moraes, C.; Zaverucha-Do-Valle, C.; Fleurance, R.; Sharshar, T.; Bozza, F.; D’Avila, J. Neuroinflammation in Sepsis: Molecular Pathways of Microglia Activation. Pharmaceuticals 2021, 14, 416. [Google Scholar] [CrossRef]
- Webber, R.J.; Sweet, R.M.; Webber, D.S. Circulating Microvesicle-Associated Inducible Nitric Oxide Synthase Is a Novel Therapeutic Target to Treat Sepsis: Current Status and Future Considerations. Int. J. Mol. Sci. 2021, 22, 13371. [Google Scholar] [CrossRef]
- Michels, M.; Steckert, A.V.; Quevedo, J.; Barichello, T.; Dal-Pizzol, F. Mechanisms of long-term cognitive dysfunction of sepsis: From blood-borne leukocytes to glial cells. Intensive Care Med. Exp. 2015, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Ye, B.; Tao, T.; Zhao, A.; Wen, L.; He, X.; Liu, Y.; Fu, Q.; Mi, W.; Lou, J. Blockade of IL-17A/IL-17R Pathway Protected Mice from Sepsis-Associated Encephalopathy by Inhibition of Microglia Activation. Mediat. Inflamm. 2019, 2019, 8461725. [Google Scholar] [CrossRef]
- Kobashi, S.; Terashima, T.; Katagi, M.; Nakae, Y.; Okano, J.; Suzuki, Y.; Urushitani, M.; Kojima, H. Transplantation of M2-Deviated Microglia Promotes Recovery of Motor Function after Spinal Cord Injury in Mice. Mol. Ther. 2020, 28, 254–265. [Google Scholar] [CrossRef]
- Kwon, H.S.; Koh, S.-H. Neuroinflammation in neurodegenerative disorders: The roles of microglia and astrocytes. Transl. Neurodegener. 2020, 9, 42. [Google Scholar] [CrossRef]
- Qin, X.; Wang, W.; Wu, H.; Liu, D.; Wang, R.; Xu, J.; Jiang, H.; Pan, F. PPARγ-mediated microglial activation phenotype is involved in depressive-like behaviors and neuroinflammation in stressed C57BL/6J and ob/ob mice. Psychoneuroendocrinology 2020, 117, 104674. [Google Scholar] [CrossRef]
- Subhramanyam, C.S.; Wang, C.; Hu, Q.; Dheen, S.T. Microglia-mediated neuroinflammation in neurodegenerative diseases. Semin. Cell Dev. Biol. 2019, 94, 112–120. [Google Scholar] [CrossRef]
- Ren, C.; Li, D.; Zhou, Q.; Hu, X. Mitochondria-targeted TPP-MoS2 with dual enzyme activity provides efficient neuroprotection through M1/M2 microglial polarization in an Alzheimer’s disease model. Biomaterials 2020, 232, 119752. [Google Scholar] [CrossRef]
- Xie, Z.; Zhao, J.; Wang, H.; Jiang, Y.; Yang, Q.; Fu, Y.; Zeng, H.; Hölscher, C.; Xu, J.; Zhang, Z. Magnolol alleviates Alzheimer’s disease-like pathology in transgenic C. elegans by promoting microglia phagocytosis and the degradation of beta-amyloid through activation of PPAR-γ. Biomed. Pharmacother. 2020, 124, 109886. [Google Scholar] [CrossRef]
- Chu, F.; Shi, M.; Zheng, C.; Shen, D.; Zhu, J.; Zheng, X.; Cui, L. The roles of macrophages and microglia in multiple sclerosis and experimental autoimmune encephalomyelitis. J. Neuroimmunol. 2018, 318, 1–7. [Google Scholar] [CrossRef]
- Zhang, Y.; Feng, S.; Nie, K.; Li, Y.; Gao, Y.; Gan, R.; Wang, L.; Li, B.; Sun, X.; Wang, L.; et al. TREM2 modulates microglia phenotypes in the neuroinflammation of Parkinson’s disease. Biochem. Biophys. Res. Commun. 2018, 499, 797–802. [Google Scholar] [CrossRef]
- Bok, E.; Chung, Y.C.; Kim, K.-S.; Baik, H.H.; Shin, W.-H.; Jin, B.K. Modulation of M1/M2 polarization by capsaicin contributes to the survival of dopaminergic neurons in the lipopolysaccharide-lesioned substantia nigra in vivo. Exp. Mol. Med. 2018, 50, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, H.; Li, R.; Han, C.; Lu, X.; Zhang, H. Minocycline promotes posthemorrhagic neurogenesis via M2 microglia polarization via upregulation of the TrkB/BDNF pathway in rats. J. Neurophysiol. 2018, 120, 1307–1317. [Google Scholar] [CrossRef] [PubMed]
- Tufano, M.; Pinna, G. Is There a Future for PPARs in the Treatment of Neuropsychiatric Disorders? Molecules 2020, 25, 1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holtman, I.R.; Skola, D.; Glass, C.K. Transcriptional control of microglia phenotypes in health and disease. J. Clin. Investig. 2017, 127, 3220–3229. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Zhu, B.; Cheon, I.S.; Goplen, N.P.; Jiang, L.; Zhang, R.; Peebles, R.S.; Mack, M.; Kaplan, M.H.; Limper, A.H.; et al. PPAR-γ in Macrophages Limits Pulmonary Inflammation and Promotes Host Recovery following Respiratory Viral Infection. J. Virol. 2019, 93, e00030-19. [Google Scholar] [CrossRef] [Green Version]
- Ciavarella, C.; Motta, I.; Valente, S.; Pasquinelli, G. Pharmacological (or Synthetic) and Nutritional Agonists of PPAR-γ as Candidates for Cytokine Storm Modulation in COVID-19 Disease. Molecules 2020, 25, 2076. [Google Scholar] [CrossRef]
- Souza, C.O.; Teixeira, A.A.; Biondo, L.A.; Silveira, L.S.; Breda, C.N.D.S.; Braga, T.T.; Camara, N.O.; Belchior, T.; Festuccia, W.; Diniz, T.A.; et al. Palmitoleic acid reduces high fat diet-induced liver inflammation by promoting PPAR-γ-independent M2a polarization of myeloid cells. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2020, 1865, 158776. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Abdalla, H.B.; Napimoga, M.H.; Lopes, A.H.; de Macedo Maganin, A.G.; Cunha, T.M.; Van Dyke, T.E.; Napimoga, J.T.C. Activation of PPAR-γ induces macrophage polarization and reduces neutrophil migration mediated by heme oxygenase. Int. Immunopharmacol. 2020, 84, 106565. [Google Scholar] [CrossRef]
- Zhou, D.; Ji, L.; Chen, Y. TSPO Modulates IL-4-Induced Microglia/Macrophage M2 Polarization via PPAR-γ Pathway. J. Mol. Neurosci. 2019, 70, 542–549. [Google Scholar] [CrossRef]
- Li, H.; Jiang, T.; Li, M.-Q.; Zheng, X.-L.; Zhao, G.-J. Transcriptional Regulation of Macrophages Polarization by MicroRNAs. Front. Immunol. 2018, 9, 1175. [Google Scholar] [CrossRef]
- Zhang, J.; Yi, S.; Xiao, C.; Li, Y.; Liu, C.; Jiang, W.; Yang, C.; Zhou, T. Asperosaponin VI inhibits LPS-induced inflammatory response by activating PPAR-γ pathway in primary microglia. Saudi J. Biol. Sci. 2020, 27, 3138–3144. [Google Scholar] [CrossRef]
- Baidoo, J.; Mukherjee, S.; Kashfi, K.; Banerjee, P. A New Perspective on Cancer Therapy: Changing the Treaded Path? Int. J. Mol. Sci. 2021, 22, 9836. [Google Scholar] [CrossRef]
- Mitchell, J.P.; Carmody, R.J. NF-κB and the Transcriptional Control of Inflammation. Int. Rev. Cell Mol. Biol. 2018, 335, 41–84. [Google Scholar] [CrossRef]
- Mu, P.-W.; Jiang, P.; Wang, M.-M.; Chen, Y.-M.; Zheng, S.-H.; Tan, Z.; Jiang, W.; Zeng, L.-Y.; Wang, T.-H. Oestrogen exerts anti-inflammation via p38 MAPK/NF-κB cascade in adipocytes. Obes. Res. Clin. Pract. 2016, 10, 633–641. [Google Scholar] [CrossRef]
- Giridharan, S.; Srinivasan, M. Mechanisms of NF-κB p65 and strategies for therapeutic manipulation. J. Inflamm. Res. 2018, 11, 407–419. [Google Scholar] [CrossRef] [Green Version]
- Giuliani, C.; Bucci, I.; Napolitano, G. The Role of the Transcription Factor Nuclear Factor-kappa B in Thyroid Autoimmunity and Cancer. Front. Endocrinol. 2018, 9, 471. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Chen, J.; Lin, L.; Pan, G.; Zhang, S.; Chen, H.; Zhang, M.; Xuan, Y.; Wang, Y.; You, Z. Quzhou Fructus Aurantii Extract suppresses inflammation via regulation of MAPK, NF-κB, and AMPK signaling pathway. Sci. Rep. 2020, 10, 1593. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Simion, V.; Pierce, J.B.; Haemmig, S.; Chen, A.F.; Feinberg, M.W. LncRNA-MAP3K4 regulates vascular inflammation through the p38 MAPK signaling pathway and cis-modulation of MAP3K. FASEB J. 2021, 35, e21133. [Google Scholar] [CrossRef]
- Dong, P.; Ji, X.; Han, W.; Han, H. Oxymatrine exhibits anti-neuroinflammatory effects on Aβ1–42-induced primary microglia cells by inhibiting NF-κB and MAPK signaling pathways. Int. Immunopharmacol. 2019, 74, 105686. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liu, B.; Yang, L.-E.; Zhang, C. Platycodigenin as Potential Drug Candidate for Alzheimer’s Disease via Modulating Microglial Polarization and Neurite Regeneration. Molecules 2019, 24, 3207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keledjian, K.; Tsymbalyuk, O.; Semick, S.; Moyer, M.; Negoita, S.; Kim, K.; Ivanova, S.; Gerzanich, V.; Simard, J.M. The peroxisome proliferator-activated receptor gamma (PPARγ) agonist, rosiglitazone, ameliorates neurofunctional and neuroinflammatory abnormalities in a rat model of Gulf War Illness. PLoS ONE 2020, 15, e0242427. [Google Scholar] [CrossRef] [PubMed]
- Zusso, M.; Lunardi, V.; Franceschini, D.; Pagetta, A.; Lo, R.; Stifani, S.; Frigo, A.C.; Giusti, P.; Moro, S. Ciprofloxacin and levofloxacin attenuate microglia inflammatory response via TLR4/NF-kB pathway. J. Neuroinflamm. 2019, 16, 148. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Huang, M.-W.; Xiao, C.-X.; Long, F.; Wang, Y.; Liu, S.-Y.; Jia, W.-W.; Wu, W.-J.; Yang, D.; Hu, J.-F.; et al. Matairesinol Suppresses Neuroinflammation and Migration Associated with Src and ERK1/2-NF-κB Pathway in Activating BV2 Microglia. Neurochem. Res. 2017, 42, 2850–2860. [Google Scholar] [CrossRef]
- Li, X.; Guo, Q.; Ye, Z.; Wang, E.; Zou, W.; Sun, Z.; He, Z.; Zhong, T.; Weng, Y.; Pan, Y. PPAR γ Prevents Neuropathic Pain by Down-Regulating CX3CR1 and Attenuating M1 Activation of Microglia in the Spinal Cord of Rats Using a Sciatic Chronic Constriction Injury Model. Front. Neurosci. 2021, 15, 307. [Google Scholar] [CrossRef]
- Tian, Y.; Yang, C.; Yao, Q.; Qian, L.; Liu, J.; Xie, X.; Ma, W.; Nie, X.; Lai, B.; Xiao, L.; et al. Procyanidin B2 Activates PPARγ to Induce M2 Polarization in Mouse Macrophages. Front. Immunol. 2019, 10, 1895. [Google Scholar] [CrossRef] [Green Version]
- Orekhov, A.N.; Orekhova, V.A.; Nikiforov, N.G.; Myasoedova, V.A.; Grechko, A.V.; Romanenko, E.B.; Zhang, N.; Chistiakov, D.A. Monocyte differentiation and macrophage polarization. Vessel Plus 2019, 3, 10. [Google Scholar] [CrossRef]
- Lu, H.; Wen, D.; Sun, J.; Zeng, L.; Du, J.; Du, D.; Zhang, L.; Deng, J.; Jiang, J.; Zhang, A. Enhancer polymorphism rs10865710 associated with traumatic sepsis is a regulator of PPARG gene expression. Crit. Care 2019, 23, 430. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.; Ge, Y.; Wang, F.; Ming, Y.; Wu, Z.; Wang, J.; Sun, S.; Huang, S.; Chen, M.; Xiao, W.; et al. Protectin DX ameliorates inflammation in sepsis-induced acute lung injury through mediating PPARγ/NF-κB pathway. Immunol. Res. 2020, 68, 280–288. [Google Scholar] [CrossRef]
- Li, Z.; Jia, Y.; Feng, Y.; Cui, R.; Wang, Z.; Qu, K.; Liu, C.; Zhang, J. Methane-Rich Saline Protects Against Sepsis-Induced Liver Damage by Regulating the PPAR-γ/NF-κB Signaling Pathway. Shock 2019, 52, e163–e172. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, N.; Shi, G.; Wang, H. Geniposide ameliorated sepsis-induced acute kidney injury by activating PPARγ. Aging 2020, 12, 22744–22758. [Google Scholar] [CrossRef]
- Parikh, S.M. Metabolic Stress Resistance in Acute Kidney Injury: Evidence for a PPAR-Gamma-Coactivator-1 Alpha-Nicotinamide Adenine Dinucleotide Pathway. Nephron 2019, 143, 184–187. [Google Scholar] [CrossRef]
- Peng, S.; Xu, J.; Ruan, W.; Li, S.; Xiao, F. PPAR-γ Activation Prevents Septic Cardiac Dysfunction via Inhibition of Apoptosis and Necroptosis. Oxidative Med. Cell. Longev. 2017, 2017, 8326749. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.; Chen, L.; Liu, H.; Sun, Z.; Yang, W.; Yang, Y.; Cui, S.; Li, S.; Wang, Y.; Song, L.; et al. Protectin DX increases survival in a mouse model of sepsis by ameliorating inflammation and modulating macrophage phenotype. Sci. Rep. 2017, 7, 99. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xu, Y.; Zhang, P.; Ruan, W.; Zhang, L.; Yuan, S.; Pang, T.; Jia, A.-Q. Smiglaside A ameliorates LPS-induced acute lung injury by modulating macrophage polarization via AMPK-PPARγ pathway. Biochem. Pharmacol. 2018, 156, 385–395. [Google Scholar] [CrossRef]
- Gong, W.; Zhu, H.; Lu, L.; Hou, Y.; Dou, H. A Benzenediamine Analog FC-99 Drives M2 Macrophage Polarization and Alleviates Lipopolysaccharide- (LPS-) Induced Liver Injury. Mediat. Inflamm. 2019, 2019, 7823069. [Google Scholar] [CrossRef] [Green Version]
- Sagheddu, C.; Melis, M.; Muntoni, A.L.; Pistis, M. Repurposing Peroxisome Proliferator-Activated Receptor Agonists in Neurological and Psychiatric Disorders. Pharmaceuticals 2021, 14, 1025. [Google Scholar] [CrossRef]
- Layrolle, P.; Payoux, P.; Chavanas, S. PPAR Gamma and Viral Infections of the Brain. Int. J. Mol. Sci. 2021, 22, 8876. [Google Scholar] [CrossRef]
- Carvalho, M.; Gonçalves-De-Albuquerque, C.; Silva, A. PPAR Gamma: From Definition to Molecular Targets and Therapy of Lung Diseases. Int. J. Mol. Sci. 2021, 22, 805. [Google Scholar] [CrossRef]
- Montaigne, D.; Butruille, L.; Staels, B. PPAR control of metabolism and cardiovascular functions. Nat. Rev. Cardiol. 2021, 18, 809–823. [Google Scholar] [CrossRef]
- Machado, M.M.F.; Bassani, T.B.; Cóppola-Segovia, V.; Moura, E.L.R.; Zanatta, S.M.; Andreatini, R.; Vital, M.A.B.F. PPAR-γ agonist pioglitazone reduces microglial proliferation and NF-κB activation in the substantia nigra in the 6-hydroxydopamine model of Parkinson’s disease. Pharmacol. Rep. 2018, 71, 556–564. [Google Scholar] [CrossRef]
- Assaf, N.; El-Shamarka, M.E.; Salem, N.A.; Khadrawy, Y.A.; El Sayed, N.S. Neuroprotective effect of PPAR alpha and gamma agonists in a mouse model of amyloidogenesis through modulation of the Wnt/beta catenin pathway via targeting alpha- and beta-secretases. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 97, 109793. [Google Scholar] [CrossRef]
- Zhao, Y.; Wei, X.; Song, J.; Zhang, M.; Huang, T.; Qin, J. Peroxisome Proliferator-Activated Receptor γ Agonist Rosiglitazone Protects Blood–Brain Barrier Integrity Following Diffuse Axonal Injury by Decreasing the Levels of Inflammatory Mediators Through a Caveolin-1-Dependent Pathway. Inflammation 2018, 42, 841–856. [Google Scholar] [CrossRef]
- Zhang, M.; Hu, M.; Montera, M.A.; Westlund, K.N. Sustained relief of trigeminal neuropathic pain by a blood–brain barrier penetrable PPAR gamma agonist. Mol. Pain 2019, 15, 1744806919884498. [Google Scholar] [CrossRef] [Green Version]
- Silva-Abreu, M.; Calpena, A.C.; Andrés-Benito, P.; Aso, E.; Romero, I.A.; Roig-Carles, D.; Gromnicova, R.; Espina, M.; Ferrer, I.; García, M.L.; et al. PPARγ agonist-loaded PLGA-PEG nanocarriers as a potential treatment for Alzheimer’s disease: In vitro and in vivo studies. Int. J. Nanomed. 2018, 13, 5577–5590. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, R.G.R.; Coutinho, A.J.; Pinheiro, M.; Neves, A.R. Nanoparticles for Targeted Brain Drug Delivery: What Do We Know? Int. J. Mol. Sci. 2021, 22, 11654. [Google Scholar] [CrossRef]
- Sarathlal, K.C.; Kakoty, V.; Marathe, S.; Chitkara, D.; Taliyan, R. Exploring the Neuroprotective Potential of Rosiglitazone Embedded Nanocarrier System on Streptozotocin Induced Mice Model of Alzheimer’s Disease. Neurotox. Res. 2021, 39, 240–255. [Google Scholar] [CrossRef]
- Rzemieniec, J.; Castiglioni, L.; Gelosa, P.; Muluhie, M.; Mercuriali, B.; Sironi, L. Nuclear Receptors in Myocardial and Cerebral Ischemia-Mechanisms of Action and Therapeutic Strategies. Int. J. Mol. Sci. 2021, 22, 12326. [Google Scholar] [CrossRef]
- Gunne, S.; Heinicke, U.; Parnham, M.J.; Laux, V.; Zacharowski, K.; Von Knethen, A. Nrf2—A Molecular Target for Sepsis Patients in Critical Care. Biomolecules 2020, 10, 1688. [Google Scholar] [CrossRef]
- Dovinova, I.; Kvandová, M.; Balis, P.; Gresova, L.; Majzunova, M.; Horakova, L.; Chan, J.Y.; Barancik, M. The role of Nrf2 and PPARgamma in the improvement of oxidative stress in hypertension and cardiovascular diseases. Physiol. Res. 2020, 69 (Suppl. 4), S541–S553. [Google Scholar] [CrossRef] [PubMed]
- Jayaram, S.; Krishnamurthy, P.T. Role of microgliosis, oxidative stress and associated neuroinflammation in the pathogenesis of Parkinson’s disease: The therapeutic role of Nrf2 activators. Neurochem. Int. 2021, 145, 105014. [Google Scholar] [CrossRef] [PubMed]
- Corona, J.C.; Duchen, M.R. PPARγ as a therapeutic target to rescue mitochondrial function in neurological disease. Free Radic. Biol. Med. 2016, 100, 153–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Orhan, I.E.; Banach, M.; Rollinger, J.M.; Barreca, D.; Weckwerth, W.; Bauer, R.; Bayer, E.A.; et al. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Yu, S.; Liu, M.; Hu, K. Natural products: Potential therapeutic agents in multiple sclerosis. Int. Immunopharmacol. 2019, 67, 87–97. [Google Scholar] [CrossRef]
- Garcia, C.; Gómez-Cañas, M.; Burgaz, S.; Palomares, B.; Gómez-Gálvez, Y.; Palomo-Garo, C.; Campo, S.; Ferrer-Hernández, J.; Pavicic, C.; Navarrete, C.; et al. Benefits of VCE-003.2, a cannabigerol quinone derivative, against inflammation-driven neuronal deterioration in experimental Parkinson’s disease: Possible involvement of different binding sites at the PPARγ receptor. J. Neuroinflamm. 2018, 15, 19. [Google Scholar] [CrossRef] [Green Version]
- Bernardo, A.; Plumitallo, C.; De Nuccio, C.; Visentin, S.; Minghetti, L. Curcumin promotes oligodendrocyte differentiation and their protection against TNF-α through the activation of the nuclear receptor PPAR-γ. Sci. Rep. 2021, 11, 4952. [Google Scholar] [CrossRef]
- Naeimi, R.; Safarpour, F.; Hashemian, M.; Tashakorian, H.; Ahmadian, S.R.; Ashrafpour, M.; Ghasemi-Kasman, M. Curcumin-loaded nanoparticles ameliorate glial activation and improve myelin repair in lyolecithin-induced focal demyelination model of rat corpus callosum. Neurosci. Lett. 2018, 674, 1–10. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, X.; Ciric, B.; Curtis, M.T.; Chen, W.-J.; Rostami, A.; Zhang, G.-X. A dual Effect of Ursolic Acid to the Treatment of Multiple Sclerosis through Both Immunomodulation and Direct Remyelination. Proc. Natl. Acad. Sci. USA 2020, 117, 9082–9093. [Google Scholar] [CrossRef]
- Silva, A.R.; Moraes, B.P.T.; Gonçalves-De-Albuquerque, C.F. Mediterranean Diet: Lipids, Inflammation, and Malaria Infection. Int. J. Mol. Sci. 2020, 21, 4489. [Google Scholar] [CrossRef]
- Gonçalves-De-Albuquerque, C.F.; Medeiros-De-Moraes, I.M.; Oliveira, F.M.D.J.; Burth, P.; Bozza, P.; Faria, M.V.C.; Silva, A.R.; De Castro-Faria-Neto, H.C. Omega-9 Oleic Acid Induces Fatty Acid Oxidation and Decreases Organ Dysfunction and Mortality in Experimental Sepsis. PLoS ONE 2016, 11, e0153607. [Google Scholar] [CrossRef] [Green Version]
- Medeiros-De-Moraes, I.M.; Gonçalves-De-Albuquerque, C.F.; Kurz, A.R.M.; de Jesus Oliveira, F.M.; De Abreu, V.H.P.; Torres, R.C.; Carvalho, V.F.; Estato, V.; Bozza, P.; Sperandio, M.; et al. Omega-9 Oleic Acid, the Main Compound of Olive Oil, Mitigates Inflammation during Experimental Sepsis. Oxidative Med. Cell. Longev. 2018, 2018, 6053492. [Google Scholar] [CrossRef] [Green Version]
- Romero-Molina, C.; Navarro, V.; Jimenez, S.; Muñoz-Castro, C.; Sanchez-Mico, M.V.; Gutierrez, A.; Vitorica, J.; Vizuete, M. Should We Open Fire on Microglia? Depletion Models as Tools to Elucidate Microglial Role in Health and Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 9734. [Google Scholar] [CrossRef]
- Li, Q.; Shen, C.; Liu, Z.; Ma, Y.; Wang, J.; Dong, H.; Zhang, X.; Wang, Z.; Yu, M.; Ci, L.; et al. Partial depletion and repopulation of microglia have different effects in the acute MPTP mouse model of Parkinson’s disease. Cell Prolif. 2021, 54, e13094. [Google Scholar] [CrossRef]
- Chen, Y.; Hong, T.; Chen, F.; Sun, Y.; Wang, Y.; Cui, L. Interplay Between Microglia and Alzheimer’s Disease—Focus on the Most Relevant Risks: APOE Genotype, Sex and Age. Front. Aging Neurosci. 2021, 13, 631827. [Google Scholar] [CrossRef]
- De Sousa, V.L.; Araújo, S.B.; Antonio, L.M.; Silva-Queiroz, M.; Colodeti, L.C.; Soares, C.; Barros-Aragão, F.; Mota-Araujo, H.P.; Alves, V.S.; Coutinho-Silva, R.; et al. Innate immune memory mediates increased susceptibility to Alzheimer’s disease-like pathology in sepsis surviving mice. Brain Behav. Immun. 2021, 95, 287–298. [Google Scholar] [CrossRef]
- Cudaback, E.; Graykowski, D. Don’t know what you got till it’s gone: Microglial depletion and neurodegeneration. Neural Regen. Res. 2021, 16, 1921–1927. [Google Scholar] [CrossRef]
- Shulyatnikova, T.; Verkhratsky, A. Astroglia in Sepsis Associated Encephalopathy. Neurochem. Res. 2020, 45, 83–99. [Google Scholar] [CrossRef]
- Araújo, C.; Estato, V.; Tibiriçá, E.; Bozza, P.; Castro-Faria-Neto, H.; Silva, A. PPAR gamma activation protects the brain against microvascular dysfunction in sepsis. Microvasc. Res. 2012, 84, 218–221. [Google Scholar] [CrossRef]
- Araújo, C.V.; Campbell, C.; Gonçalves-De-Albuquerque, C.F.; Molinaro, R.; Cody, M.J.; Yost, C.C.; Bozza, P.; Zimmerman, G.A.; Weyrich, A.; Castro-Faria-Neto, H.C.; et al. A PPARγ agonist enhances bacterial clearance through neutrophil extracellular trap formation and improves survival in sepsis. Shock 2016, 45, 393–403. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
| Study | Model | Main Results |
|---|---|---|
| Peng, Jing et al., 2019 [19] | Status epileptic male C57BL/6 mice aged 8–10 weeks | Polarization of microglia to M2 by PPARγ ligand rosiglitazone and protection against pilocarpine-induced status epilepticus with rescued neuron loss |
| Wen, Liang et al., 2018 [20] | Traumatic brain injury in C57BL/6J mice (10 to 12 weeks old) | M2 microglia attenuated axonal injury in the cerebral cortex and improved neurological function |
| Qin, Xiaqing et al., 2020 [45] | Chronic unpredictable mild stress in male wt mice (23–25 g, 8–10 weeks old) and male ob/ob mice (43–53 g, 8–10 weeks old) | The behavioral improvement due to the inhibition of microglia-mediated neuroinflammation and higher presence of the M2 neuroprotective phenotype in mice treated with the PPARγ agonist pioglitazone |
| Ren, Chaoxiu et al., 2020 [47] | Alzheimer disease in female APP/PS1 double transgenic mice (6 months old) | M2 microglia degraded Aβ deposits and efficiently promoted neuroprotection by inhibiting Aβ accumulation and neuroinflammation |
| Xie, Zhishen et al., 2020 [48] | Alzheimer-like disease in transgenic C. elegans | M2 microglia enhanced Aβ degradation reducing its deposition in the PPARγ-dependent mechanism |
| Zhang, Youwen et al., 2018 [50] | MPTP-intoxicated male C57BL/6 mice (5–6 weeks, weight 18–22 g) Parkinson disease model | Higher levels of M2 microglia alleviated neuroinflammation |
| Bok, Eugene et al., 2018 [51] | LPS-lesioned inflammatory model of Parkinson disease in female Sprague Dawley rats (230–280 g) | M2 microglia enhanced the survival of dopamine neurons |
| Miao, Hongsheng et al., 2018 [52] | Intracerebral hemorrhage in Sprague Dawley (SD) rats (250–350 g) | M2 microglia-derived BDNF promoted neurogenesis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro, L.V.G.; Gonçalves-de-Albuquerque, C.F.; Silva, A.R. Polarization of Microglia and Its Therapeutic Potential in Sepsis. Int. J. Mol. Sci. 2022, 23, 4925. https://doi.org/10.3390/ijms23094925
Castro LVG, Gonçalves-de-Albuquerque CF, Silva AR. Polarization of Microglia and Its Therapeutic Potential in Sepsis. International Journal of Molecular Sciences. 2022; 23(9):4925. https://doi.org/10.3390/ijms23094925
Chicago/Turabian StyleCastro, Léo Victor G., Cassiano F. Gonçalves-de-Albuquerque, and Adriana R. Silva. 2022. "Polarization of Microglia and Its Therapeutic Potential in Sepsis" International Journal of Molecular Sciences 23, no. 9: 4925. https://doi.org/10.3390/ijms23094925