Abstract
Hydrogen sulfide (H2S) has been shown to participate in various stress responses in plants, including drought, salinity, extreme temperatures, osmotic stress, and heavy metal stress. Manganese (Mn), as a necessary nutrient for plant growth, plays an important role in photosynthesis, growth, development, and enzymatic activation of plants. However, excessive Mn2+ in the soil can critically affect plant growth, particularly in acidic soil. In this study, the model plant Arabidopsis thaliana was used to explore the mechanism of H2S participation and alleviation of Mn stress. First, using wild-type Arabidopsis with excessive Mn2+ treatment, the following factors were increased: H2S content, the main H2S synthetase L-cysteine desulfhydrase enzyme (AtLCD) activity, and the expression level of the AtLCD gene. Further, using the wild-type, AtLCD deletion mutant (lcd) and overexpression lines (OE5 and OE32) as materials, the phenotype of Arabidopsis seedlings was observed by exogenous application of hydrogen sulfide donor sodium hydrosulfide (NaHS) and scavenger hypotaurine (HT) under excessive Mn2+ treatment. The results showed that NaHS can significantly alleviate the stress caused by Mn2+, whereas HT aggravates this stress. The lcd mutant is more sensitive to Mn stress than the wild type, and the overexpression lines are more resistant. Moreover, the mechanism of H2S alleviating Mn stress was determined. The Mn2+ content and the expression of the Mn transporter gene in the mutant were significantly higher than those of the wild-type and overexpression lines. The accumulation of reactive oxygen species was significantly reduced in NaHS-treated Arabidopsis seedlings and AtLCD overexpression lines, and the activities of various antioxidant enzymes (SOD, POD, CAT, APX) also significantly increased. In summary, H2S is involved in the response of Arabidopsis to Mn stress and may alleviate the inhibition of Mn stress on Arabidopsis seedling growth by reducing Mn2+ content, reducing reactive oxygen species content, and enhancing antioxidant enzyme activity. This study provides an important basis for further study of plant resistance to heavy metal stress.
1. Introduction
As a necessary trace element for plant growth, manganese (Mn) is mainly absorbed by plants in the form of divalent Mn ion (Mn2+), which plays an important role in plant growth, development, and metabolism [1]. However, when the concentration of Mn2+ in the soil exceeds a certain threshold, it will be toxic to the plant, thus affecting normal growth. Mn stress usually occurs in acidic soil. When the pH value of the soil is lower than 5.5, a large amount of soluble Mn2+ is released. In these conditions, the concentration of Mn2+ in the soil increases sharply, which leads to Mn2+ accumulation in the plant [2]. With rapid industrialization and changes in tillage methods, the area of acidic soils in the world has expanded. Mn stress has become the second-largest plant-growth-limiting factor, after aluminum toxicity [3]. Therefore, it is of great significance to explore the mechanisms of Mn stress affecting plant growth and how to alleviate it.
When subjected to Mn stress, plants will behave differently at different growth stages [4]. Overall, Mn stress has a greater effect on the above-ground part than on the root system, and the leaf is the main target of Mn. Nearly 90% of the Mn absorbed by the plant is transferred to the above-ground tissue. Excess Mn2+ can inhibit leaf photosynthesis [5]. Mn stress decreases the activity of many important enzymes in plants [6], in addition to affecting the absorption, transport, and distribution of other nutrients that destroy the root structure [7]. In plants, Mn2+ is absorbed and transported through Mn2+ transporters. Most of these transporters are transmembrane proteins, which can transport and store Mn2+ in the inner membrane organelles [8,9]. However, the mechanism of how this process is initiated by plants has not fully been revealed.
H2S is an important gas signal molecule. Studies on endogenous H2S in plants date back decades. Wilson et al. (1978) observed that the leaves of cucumber, corn, and soybeans can release H2S [10]. Rennenberg et al. (1987) found that in Arabidopsis, L-cysteine desulfhydrase and D-cysteine desulfhydrase used L/D-cysteine as a substrate to produce H2S. L/D-cysteine desulfhydrase is a key enzyme for H2S synthesis in plants, and it is also the most studied H2S synthetase [11]. In recent years, it has been found that H2S can participate in plant growth, development, and metabolism, such as enhancing plant photosynthesis, delaying flowering and senescence, and promoting seed germination [12,13,14]. Additionally, H2S can increase plant resistance to a variety of environmental stresses, including drought, high salt, extreme temperatures, and various heavy metal stresses such as chromium [15,16], cadmium [17], and aluminum [18]. It has been reported that H2S can alleviate aluminum toxicity by reducing the absorption of Al3+ and increasing the antioxidant capacity in barley [19]. H2S can regulate the AsA–GSH cycle and alleviate As toxicity to peas [20]. It can also alleviate the inhibition of Cr6+ on the roots of Arabidopsis by upregulating the heavy metal (HM) chelator synthase-encoding genes, such as PCS1, PCS2, and MT2A. Increased content of metallothionein and phytochelatins increases Arabidopsis tolerance to Cr stress [21]. However, no reports have been made on H2S response to Mn stress.
Using wild-type Arabidopsis, the AtLCD defective mutant lcd, and the AtLCD over-expression lines OELCD (OE5 and OE32) as materials, we performed physiological and biochemical methods to explore the function and mechanism of H2S in response to Mn stress in Arabidopsis.
2. Results
2.1. Effects of Mn Stress on H2S Content, AtLCD Enzyme Activity, and Gene Expression in Arabidopsis Seedlings
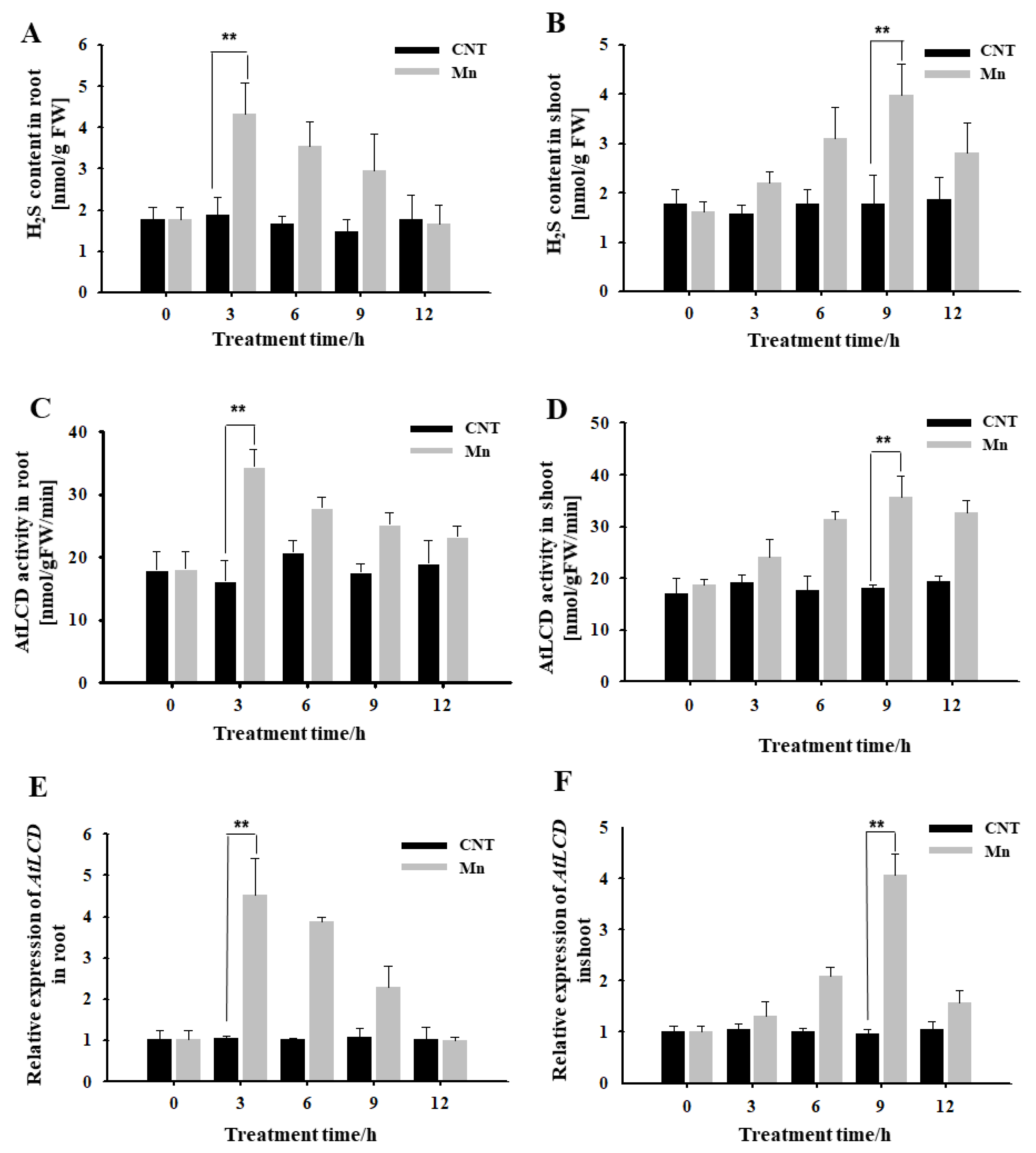
It can be seen from Figure 1 that, under hydroponic conditions, H2S content in roots treated using 4 mM Mn2+ for 3 h increased significantly, reached a high level, and then gradually decreased (Figure 1A). Both the activity of the H2S synthase AtLCD and the expression of AtLCD reached maximum levels at 3 h and then decreased (Figure 1C,E). The H2S-related indicators of the shoots were tested. It was also found that Mn2+ caused the H2S content to increase, reaching the highest level at 9 h (Figure 1B). The main synthase activity and AtLCD expression had the same trend (Figure 1D,F). Thus, it is speculated that H2S may participate in the response to Mn stress.
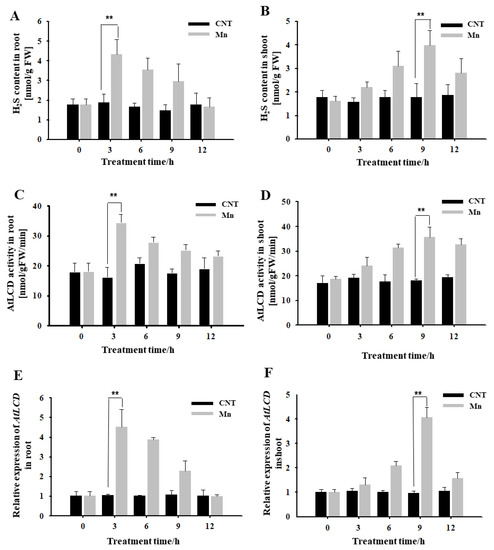
Figure 1.
Effects of Mn stress on H2S content, AtLCD enzyme activity, and gene expression in Arabidopsis seedlings. Effects of Mn stress on H2S content in Arabidopsis roots (A) and shoot (B); AtLCD enzyme activity in Arabidopsis roots (C) and shoot (D); the relative expression of AtLCD in Arabidopsis roots (E) and shoot (F). Three independent experimental replications were conducted. Values are the means ± SE of three independent experiments (** p < 0.01).
2.2. Effects of H2S on Phenotype and Growth of Arabidopsis under Mn Stress
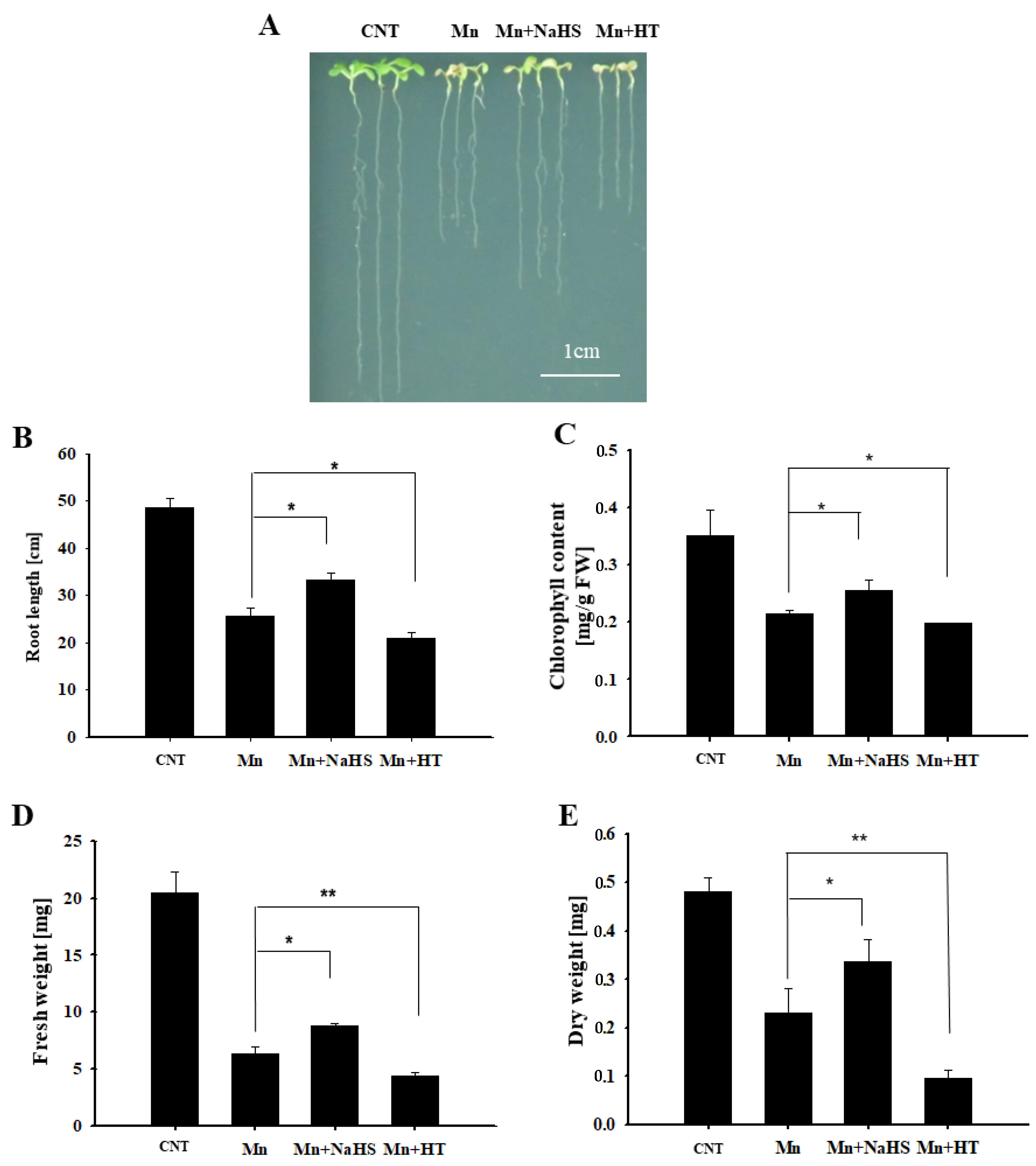
To investigate the role of H2S in Mn stress, using wild-type Arabidopsis as the material, the seedling phenotype and growth indicators were observed by exogenous application of H2S donor NaHS and scavenger HT under 4 mM Mn2+ treatment after 5 d. Compared with the control, it can be seen from Figure 2A that the growth of Arabidopsis seedlings was inhibited by Mn2+ treatment, the phenotype of Mn stress was significantly alleviated after NaHS treatment, and the phenotype of Mn stress was aggravated after HT treatment. Furthermore, the main root length, chlorophyll content, fresh weight, and dry weight were calculated, and the results were consistent with the phenotype results (Figure 2B,E). We also measured whether NaHS and HT affected H2S content under Mn stress. The results showed that H2S content increased after 5 days of Mn stress; moreover, NaHS could further enhance the content of H2S, while HT decreased H2S concentration (Figure S2). This further suggests the participation of H2S in response to Mn stress, i.e., alleviating the stress caused by Mn2 + to Arabidopsis seedlings.
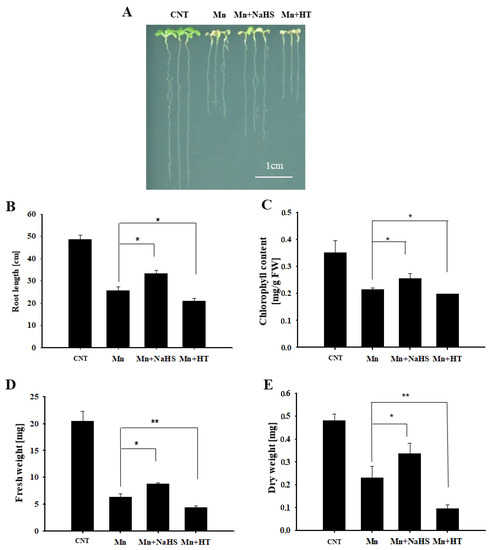
Figure 2.
Effects of NaHS and HT on the growth of wild-type Arabidopsis seedlings under Mn stress. Effects of NaHS and HT on the phenotype (A), root length (B), chlorophyll content (C), fresh weight (D), and dry weight (E) of wild-type Arabidopsis under Mn stress. Three independent experimental replications were conducted. Values are the means ± SE of three independent experiments (* p < 0.05, ** p < 0.01). Scale bar = 1 cm.
2.3. Effects on the Phenotype of lcd and OELCD Lines under Mn Stress
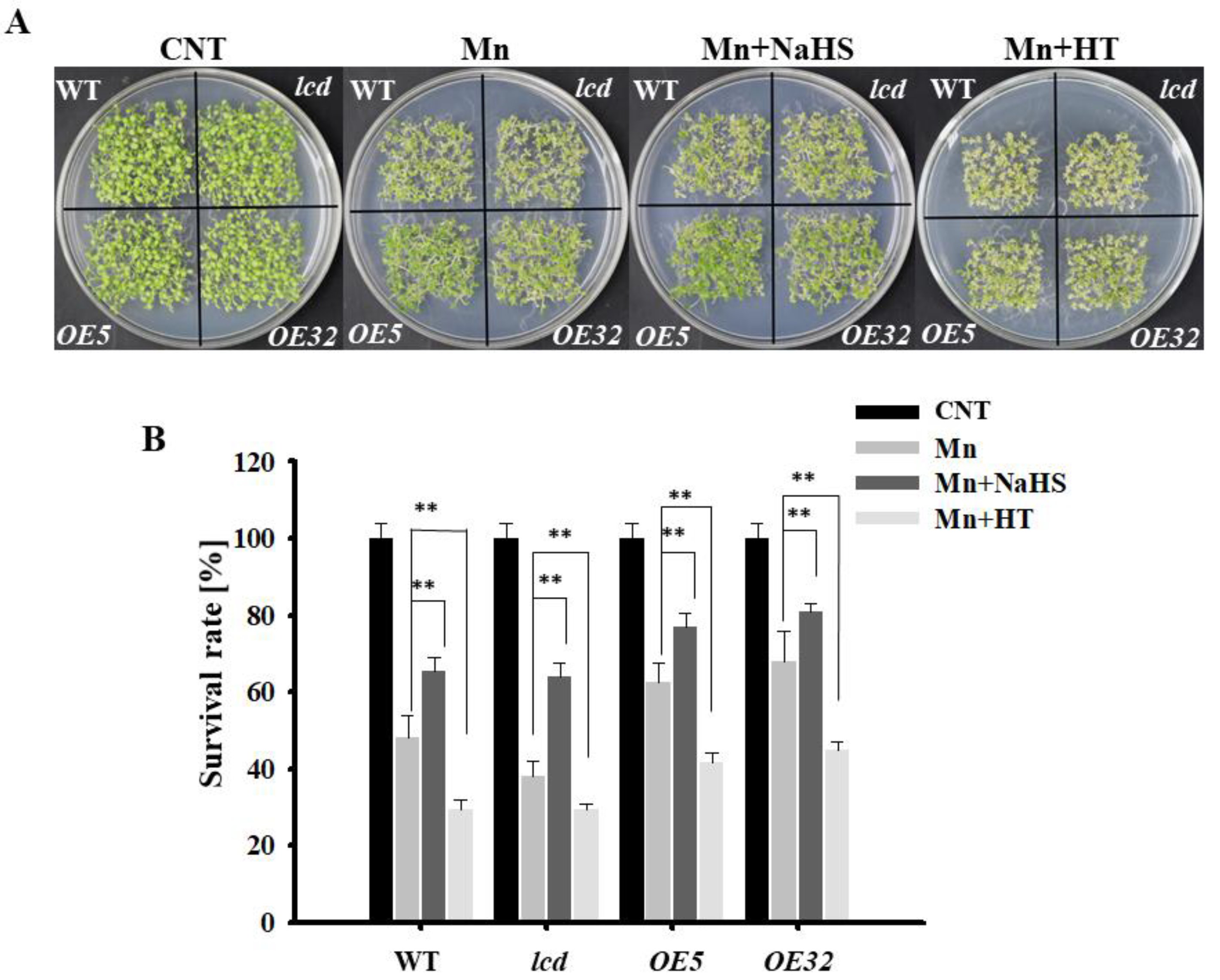
To provide genetic evidence of H2S participation in Mn stress, lcd mutant and two lines overexpressing AtLCD (OE5, OE32) were obtained and identified, and the results are shown in Supplementary Data (Figure S1). The phenotype and survival of each line after Mn stress were observed. The results are shown in Figure 3A. Overall, compared with Arabidopsis seedlings after 2 mM Mn2+ application, the phenotypes of all lines of Mn stress were alleviated after NaHS treatment, whereas the growth state of Arabidopsis seedlings was poor, and the leaves turned yellow after HT treatment. From a single treatment, after the Mn2+, Mn2+ and NaHS treatment, and Mn2+ and HT treatment, the phenotypes of both overexpression lines were significantly better than those of the wild type, whereas mutants showed poor growth, and the leaves turned yellow. The survival statistics were consistent with those of the phenotype (Figure 3B).
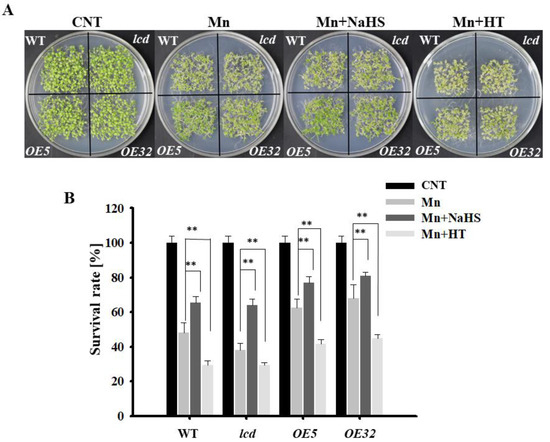
Figure 3.
Effects of NaHS and HT on the growth of lcd and OELCD under Mn stress. Effects of NaHS and HT on the phenotype (A) and survival rates (B) of wild type, lcd, and two independent overexpression Arabidopsis seedlings after Mn stress. Three independent experimental replications were conducted. Values are the means ± SE of three independent experiments (** p < 0.01).
2.4. Effects on Mn Transporter-Related Gene Expression in Roots of lcd and OELCD under Mn Stress
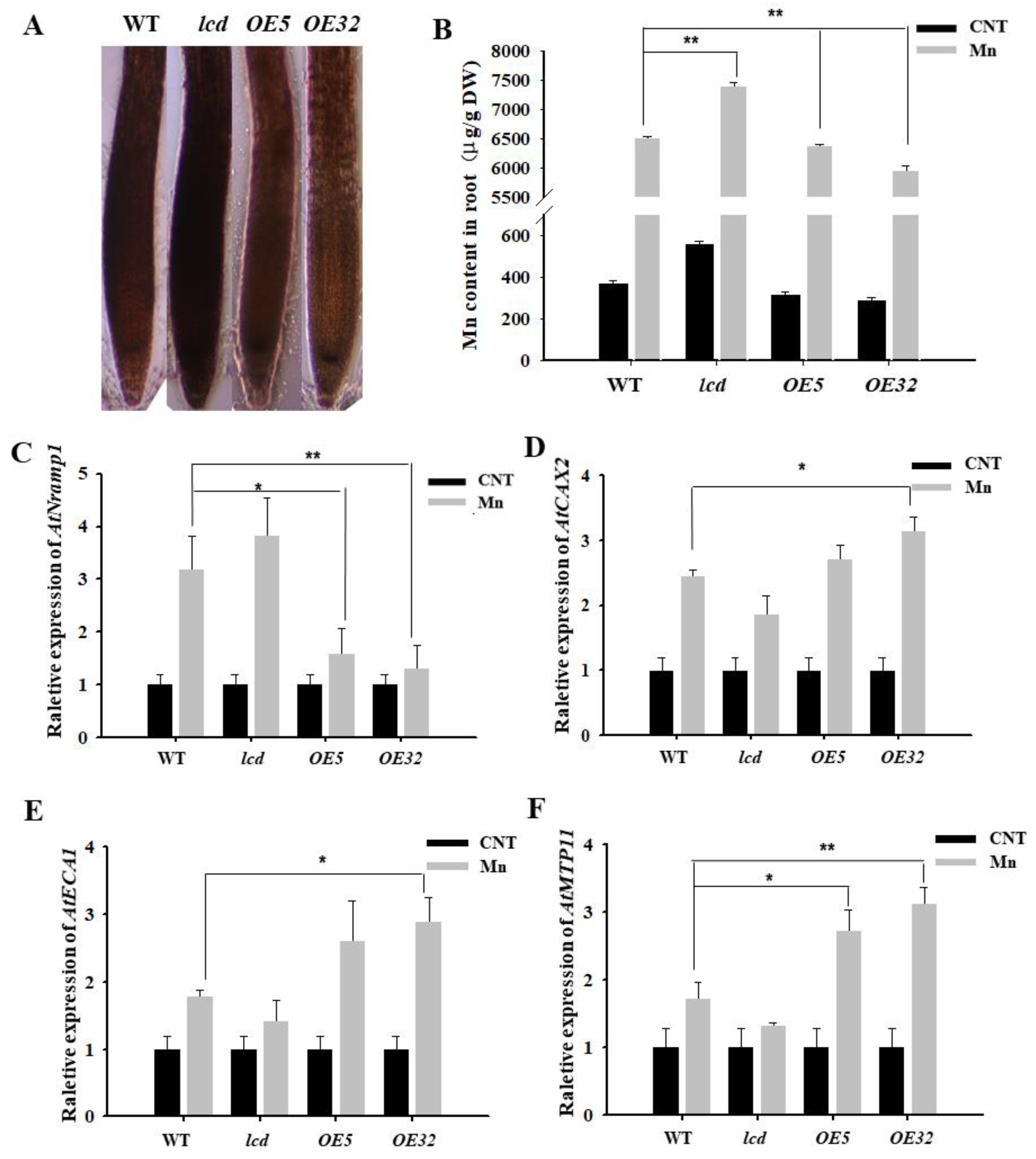
Hematoxylin is often used to observe the distribution of metal ions in plant roots [22]. Under Mn stress, the combination of hematoxylin with Mn2+ in root cells shows a purple color; the darker the purple color, the more Mn2+ in root cells. After 4 mM Mn2+ treatment of WT, lcd, and OELCD, hematoxylin staining was performed, and the results are shown in Figure 4A. The roots of lcd were stained deeper than those of the wild type, and the roots of OE5 and OE32 were stained shallowly. As hematoxylin staining is not specific to Mn, Mn content in roots was analyzed further by inductively coupled plasma atomic emission spectrophotometry (ICP–MS). The differences between wild type and other lines were compared. The content of Mn in OE5 and OE32 lines was significantly lower than that in the wild type under Mn stress, whereas, in lcd mutant, Mn content was significantly higher than that of the wild type (Figure 4B). It is, therefore, speculated that H2S may reduce the root tissue Mn content under Mn stress.
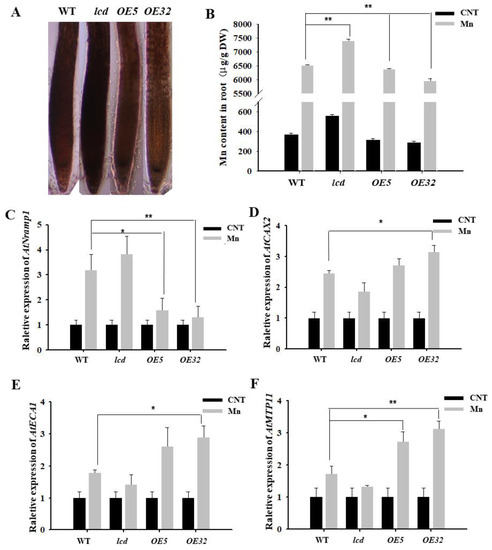
Figure 4.
Effects on Mn transporter-related gene expression in roots of lcd and OELCD under Mn stress. Hematoxylin staining (A), Mn content (B), and the expression of AtNramp1 (C), AtCAX2 (D), AtECA1 (E), and AtMTP11 (F) in the roots of wild type, lcd, and two independent overexpression lines under Mn stress. Three independent experimental replications were conducted. Values are the means ± SE of three independent experiments (* p < 0.05; ** p < 0.01).
Furthermore, we analyzed whether a decrease in Mn content in root tissue by H2S was related to the Mn transporter. There are numerous Mn transport-related proteins in plant roots, in which AtNramp1 is located in the cell membrane and is responsible for absorbing Mn2+ from the external environment; AtCAX2, AtMTP11, AtECA1 are located in the endomembrane system, such as in vacuole membranes, endoplasmic omentum, and Golgi membranes, and they are responsible for transporting excess Mn2+ to the cell organelles when the concentration of cytoplasmic Mn2+ is too high, thereby alleviating Mn stress [23]. Does H2S reduce Mn2+ content in roots by regulating the expression of Mn transporter genes? The expression of Mn transporter-related genes in root tissues of WT, lcd, and OELCD was detected. The results reveal that 4 mM Mn treatments strongly induced the expression levels of four genes in all lines. The differences between wild type and other lines were further compared. The expression of the Mn2+-uptake-related gene AtNramp1 in OE5 and OE32 lines was significantly lower than that in the wild type under Mn stress, whereas, in lcd mutant, the gene expression level increased but did not reach a significant level (Figure 4C). At the same time, compared with wild type, the expression levels of AtCAX2, AtMTP11, and AtECA1 in lcd lines exhibited reduction but not significantly, while those of OE5 and OE32 lines were significantly higher than that of wild type (Figure 4D,F). The expression levels of four manganese-transport-related genes changed slightly in the mutant. Therefore, it is speculated that other H2S synthesis genes and AtLCD have functional redundancy. Furthermore, it is inferred that H2S may have partially prevented the root intake of Mn2+ by reducing the gene expression of Mn2+-uptake-related proteins, thereby promoting the transport of Mn2+ to organelles by increasing partial transporter gene expression.
2.5. Effects on Reactive Oxygen Species Content of Arabidopsis Seedlings under Mn Stress
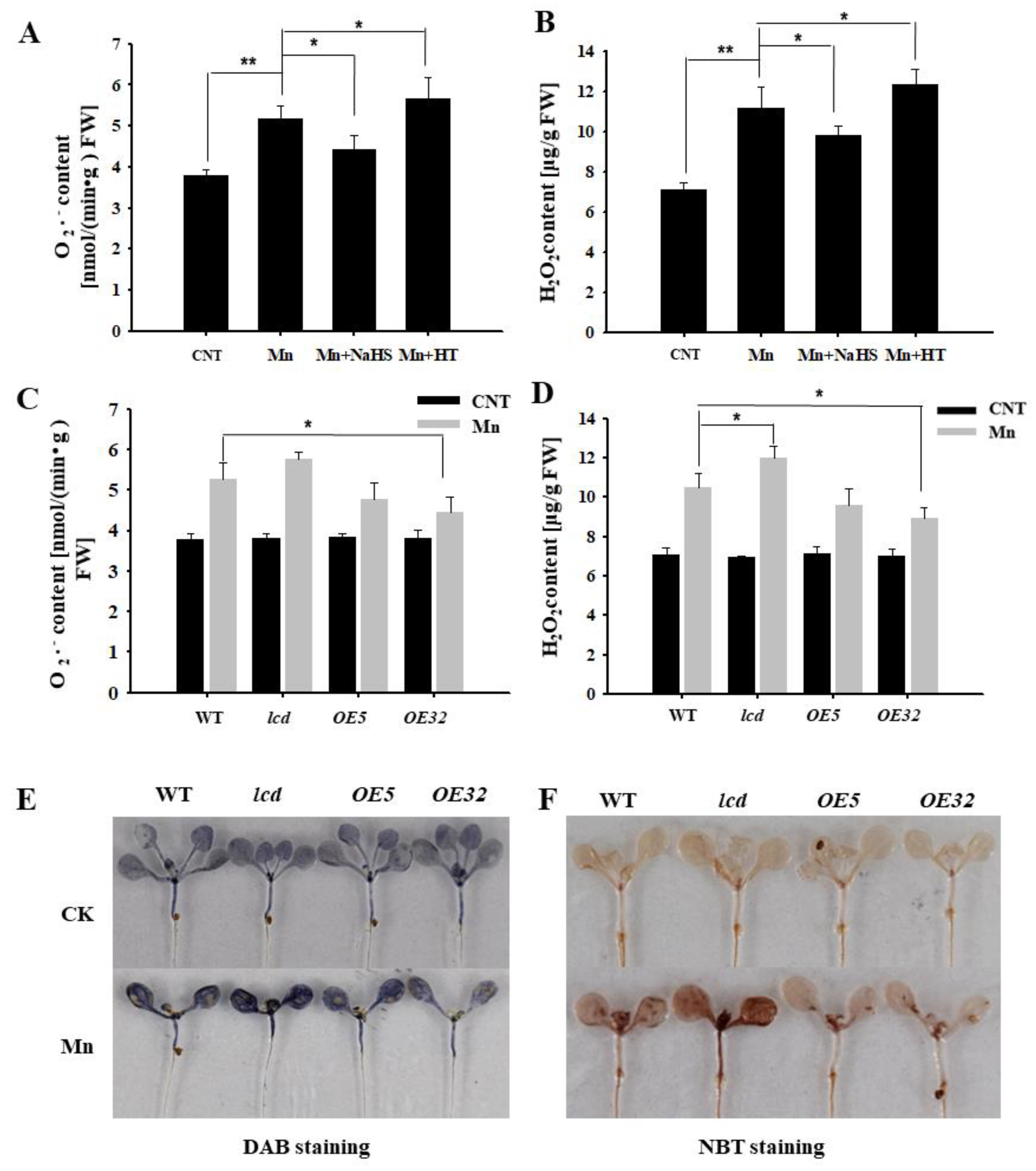
The content of reactive oxygen species in plants increases under abiotic stresses, such as heavy metal stress. Using wild-type Arabidopsis as material, the content of superoxide anion and hydrogen peroxide was detected by exogenous application of NaHS and HT in 4 mM Mn2+ treatments. Figure 5A,B show that the concentrations of O2− and H2O2 in Arabidopsis seedlings after Mn2+ treatment were significantly higher than that of control. The content of reactive oxygen speeies (ROS) was reduced with NaHS, compared with the Mn treatment, whereas the exogenous application of HT increased its content. Thus, it is speculated that exogenous H2S can reduce the reactive oxygen species in Arabidopsis seedlings under Mn stress.
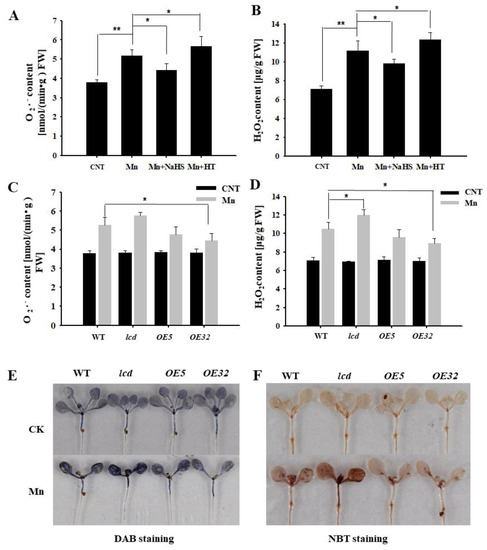
Figure 5.
Effects of H2S on H2O2 and O2− contents in Arabidopsis seedlings under Mn stress. Effects of NaHS and HT on the quantitative measurement of O2− (A) and H2O2 (B) concentrations in wild-type Arabidopsis seedlings under Mn stress. Quantitative measurement of O2− (C) and H2O2 (D) concentrations in wild type, lcd, and two independent overexpression lines seedlings treated with and without manganese. In situ accumulations of H2O2 (E) and O2− (F) before and after Mn treatment were revealed by DAB and NBT staining, respectively. Three independent experimental replications were conducted. Values are the means ± SE of three independent experiments (* p < 0.05; ** p < 0.01).
Further evidence from genetics was provided. The concentrations of O2− and H2O2 in AtLCD deficient mutants and overexpression lines were quantified under 4 mM Mn stress. The results showed that the concentrations of O2− and H2O2 in OELCD lines were significantly lower than that in wild type, whereas lcd lines were higher than that in wild type (Figure 5C,D), and the results of DAB and NBT were consistent with the above-described results (Figure 5E,F). Thus, it is speculated that H2S may alleviate Mn stress in Arabidopsis seedlings by reducing reactive oxygen species content.
2.6. Effects on Antioxidant Enzyme Activity of lcd and OELCD under Mn Stress
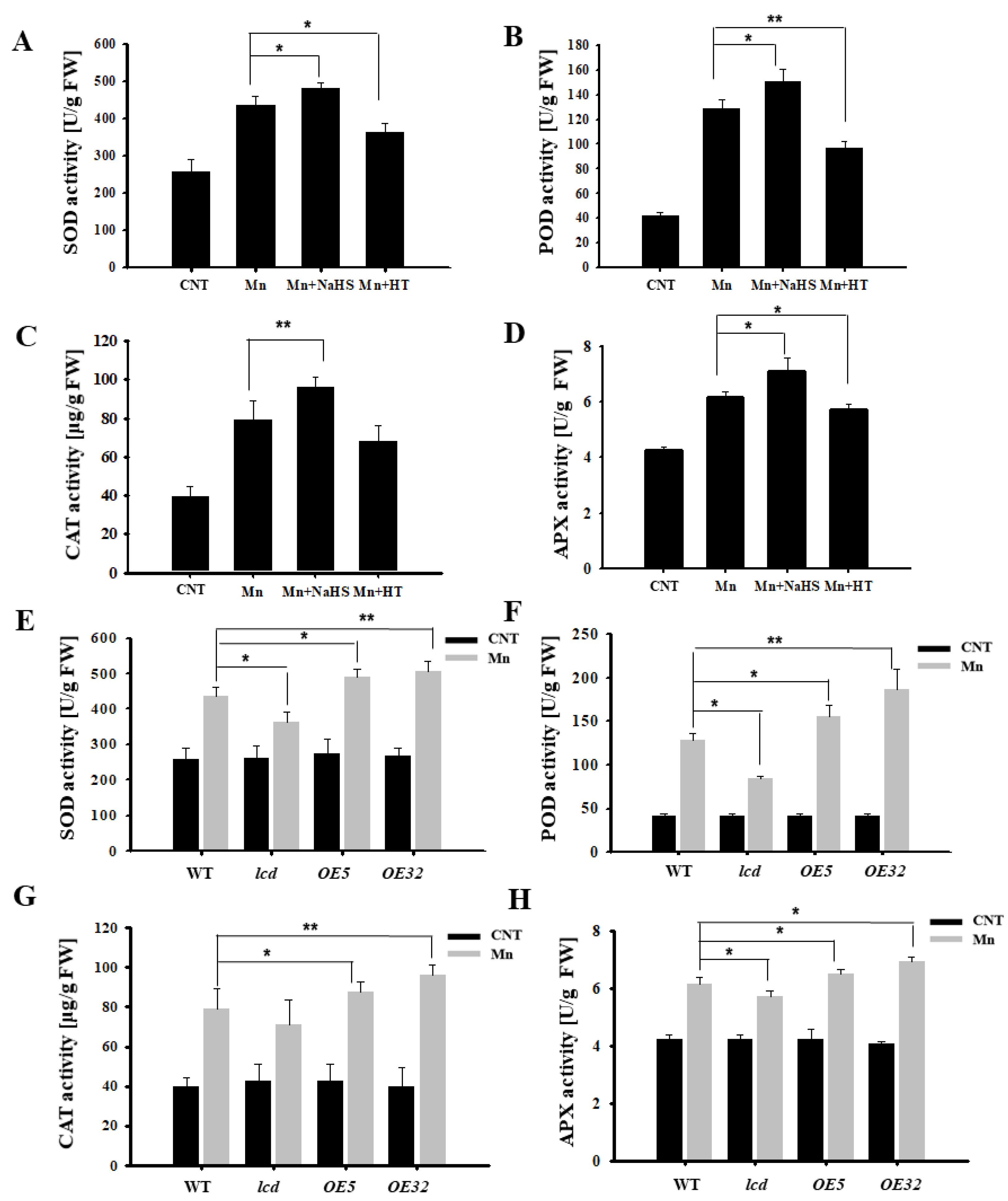
Figure 5 shows that Mn stress increased the content of ROS in Arabidopsis, so the antioxidant enzyme activity was further examined. From Figure 6A–D, it can be deduced that the SOD, POD, CAT, and APX activities of Arabidopsis under 4 mM Mn2+ treatment were significantly higher than those of control. Additionally, the antioxidant enzyme activity of Arabidopsis with Mn treatment was upregulated significantly after NaHS treatment, whereas the activity of the antioxidant enzyme decreased after HT application. Additional results found that antioxidant enzyme activity of overexpression lines was significantly higher than that in wild type, and deletion mutants were significantly lower than that in the wild type under Mn2+ treatment (Figure 6E–H). It is, therefore, suggested that H2S may alleviate Mn stress in Arabidopsis seedlings by increasing antioxidant enzyme activity.
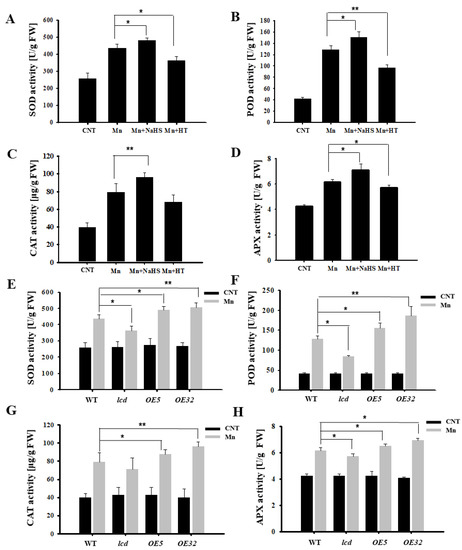
Figure 6.
Effects of H2S on antioxidant enzyme activities in Arabidopsis seedlings under Mn stress. Effects of NaHS and HT on the activity of SOD (A), POD (B), CAT (C), and APX (D) in wild-type Arabidopsis seedlings under Mn stress. The activity of SOD (E), POD (F), CAT (G), and APX (H) in wild type, lcd, and two independent overexpression line seedlings treated with and without manganese. Three independent experimental replications were conducted. Values are the means ± SE of three independent experiments (* p < 0.05; ** p < 0.01).
3. Discussion
It has been reported that H2S participates in a variety of growth and development processes, as well as stress responses, in plants [24]. In this study, we found that Mn stress can induce increased H2S content, and we also examined the enzyme activity and gene expression changes of the key synthase AtLCD in the H2S synthesis pathway. We speculate that H2S may participate in the Mn stress response (Figure 1). Through pharmacological experiments, wild-type Arabidopsis were used to observe the phenotype under Mn stress by external application of NaHS and HT, and the results showed that H2S was involved in the Mn stress activities and could alleviate the phenotype of Mn stress (Figure 2). Based on the results shown in Figure 1, the change in H2S content induced by Mn stress is from the AtLCD pathway, so further genetic evidence was provided. In Figure 3, we used 4 mM Mn at first, but the difference in phenotypes was not obvious, and the seedlings showed poor growth. It is speculated that the seeds were sowed directly in the culture medium to observe phenotype. And the time of treatment was long, the concentration was too high (4 mM), resulting in considerable damage to them. therefore, the concentration treatment was reduced (2 mM). Using lcd and OELCD as materials with Mn2+, Mn2+ and NaHS, and Mn2+ and HT treatments, phenotypic observations demonstrated that H2S alleviated Mn stress, and lcd was more pronounced than the wild-type Mn stress phenotype (Figure 3), which provided more evidence for H2S participation in Mn stress response. However, current studies have found that there are many types of synthetic H2S pathways in plants [25], and different synthetic pathways may be involved in different biological processes. Therefore, a question arises: Are there other sources of H2S in Arabidopsis besides the AtLCD pathway under Mn stress? Further studies are needed.
During plant response to heavy metal stress, the cell membrane can prevent or reduce the entry of metal ions into the cell. Additionally, plants can leave Mn in subcellular compartments, such as vacuoles, endoplasmic reticulum, Golgi, and cell walls to resist the toxic effects [23,26]. In Arabidopsis, AtNramp1 of the Nramp family is the main Mn transporter that participates in Mn uptake. AtNramp1 is localized to the plasma membrane of the epidermal cells of the root tips, and the expression of AtNramp1 is upregulated when Mn is deficient [27]. The transport protein of Arabidopsis vacuole cation exchanger (CAX) is mainly involved in the transport of Mn2+ to the vacuole. The T-DNA knockout mutant of AtCAX2 has a lower content of Mn2+ in the vacuole than that of the wild type. Overexpression of AtCAX2 in tobacco increases resistance to Mn toxicity by mediating Mn chelation into the vacuole [28]. Endoplasmic reticulum-localized Ca2+-ATPase (ECA1) is another transporter that is intended to reduce the concentration of Mn2+ in the cytoplasm, as ECA1 can pump Mn2+ from the cytoplasm into the endoplasmic reticulum. Under high Mn conditions, the Arabidopsis mutant eca1 exhibited severe Mn stress symptoms, and overexpression of ECA1 could restore the growth of eca1 mutants to normal [29]. Metal tolerance protein (MTP), which is a Mn transporter in the CDF family [30], is responsible for transporting Mn into the vacuole and Golgi bodies. AtMTP11 may be in the irregular compartment of the trans-Golgi body [9]. For this group, AtMTP11 has the highest expression and is more resistant to Mn in plant overexpression of AtMTP11 [31]. Thus, we detected the expression of Mn-transport-related genes in the above transporter. The results showed that the expression of Mn-uptake-related gene AtNramp1 in the mutant lcd line was higher than that in the wild type under Mn stress, while both the expression levels of OE5 and OE32 were significantly lower than that in wild type (Figure 4C). This suggests that H2S may have prevented the uptake of Mn2+ by root cells, in part, by inhibiting the expression of the AtNramp1 gene. This result is consistent with those related to hematoxylin staining (Figure 4A) and Mn content (Figure 4B). However, the expression levels of AtCAX2, AtMTP11, and AtECA1 in lcd lines were lower than that of wild type, while those of OE5 and OE32 were significantly higher than that of wild type (Figure 4D,F), which suggests that H2S may alleviate Mn stress by promoting Mn2+ transport to organelles.
Heavy metal stress can lead to excessive buildup of ROS, which may cause oxidative damage to biomolecules in plants [32]. H2S is a reductive substance and can directly scavenge ROS [33]. Therefore, we examined the content of ROS and the activity of antioxidant enzymes after Mn2+ treatment, and the results showed that Mn stress induced excessive ROS in plants (Figure 5). Both endogenous and exogenous H2S alleviated Mn stress in Arabidopsis seedlings by reducing ROS and significantly increasing antioxidant enzyme activity (Figure 6). It has previously been found that exogenous H2S can alleviate the degree of peroxidation caused to rice by mercury, thus alleviating the stress on rice and improving rice resistance [34]. Some studies have found that H2S alleviates oxidative stress and ionic toxicity in the cadmium-induced Arabidopsis roots through the hydrogen sulfide–cysteine circulatory system, thereby increasing the tolerance to cadmium [35]. Previous studies have found that plants respond to heavy metal copper ions, and H2S alleviates the Arabidopsis copper oxide stress process through a circulatory system with cysteine [36]. More research is warranted on whether cysteine participates in H2S involvement with Mn stress.
As a signaling molecule, how does H2S signaling occur when plants are subjected to stress? In recent years, the molecular mechanism by which H2S mediates the protein cysteine residue process in plants and animals, i.e., S-sulfhydration in post-translational modification, has been found [8,37]. Ethylene-induced hydrogen sulfide negatively regulates ethylene biosynthesis by the S-sulfhydration of ACO in tomatoes under osmotic stress [38]. Recently, it has been found that the persulfidation of SnRK2.6/OST1, a key regulatory protein of stomatal closure by H2S, promotes the activity of SnRK2.6 and its interaction with transcription factors downstream of the ABA signal, to promote stomatal closure and inhibit stomatal opening and improve the drought resistance of plants [39]. This characteristic of H2S provides an effective theoretical basis for finding its downstream regulatory proteins. Is the effect of H2S on the activity of plant antioxidant enzymes due to the S-sulfhydration modification or through other action modes? These issues need to be further studied to understand the diverse mechanisms of plant responses to Mn stress.
4. Materials and Methods
4.1. Experimental Materials
For Arabidopsis, the Columbia (Col-0) ecotype was taken as the genetic background, and the T-DNA insertion mutant (SALK_082099, named lcd) of AtLCD was purchased from the American Arabidopsis Biological Resource Center (ABRC). AtLCD overexpressing Arabidopsis was named OELCD and included 2 lines (OE5 and OE32).
4.2. Material Constructs, Cultivation, and Treatment
Full-length AtLCD (At3g62130) was obtained from the Arabidopsis Information Resource (TAIR). The cDNA fragment was amplified by PCR with the primers as follows: forward primer, 5′- CCCAAGCTTATGGAGGCGGGAGAGCG-3′ with restriction site Xba I (TaKaRa, Maebashi, Japan), reverse primer, 5′-GGGGTACCCTACAATGCAGGAAGGTTTTGAC-3′ with restriction site Kpn I (TaKaRa, Maebashi, Japan). Then, the cDNA fragment was inserted into the p-Super1300 vector (containing 35S promoter and GFP reporter gene) between restriction sites Xba I and Kpn I (TaKaRa, Maebashi, Japan). The construct was confirmed by restriction digestion and sequence analysis and then was named p-Super1300-AtLCD. Subsequently, the construct was transformed into Agrobacterium tumefaciens strain GV3101. The flower dip method was used to transform the Arabidopsis [40]. Two independent lines (OE5 and OE32) from the T3 generation were used in this research. T-DNA insertion mutant (lcd) was identified by the three-primer method. The primers sequence were as follows: LP, 5’-CACTTTTGCAAGCTTGGTTTC-3’; RP, 5’-TCAATCCAGTTGAATAAGCGC-3’; LBb1: 5’-GCCTTTTCAGAAATGGATAAATAGCCTTGCTTCCT-3’. The mutant lcd was homozygous.
The full-fledged seeds were placed in a dark treatment at 4 °C for 2 to 4 days to break the dormancy; then, the seeds were cultivated in 2% MGRL culture solution (pH = 5.8), and the incubator was put in a light environment, which was adjusted to 22 °C with a light–dark cycle of 16 h/8 h. After 7 days, the seedlings were treated with 4 mM MnCl2, which was added to the MGRL culture solution; then, the material was collected for 0, 3, 6, 9, and 12 h to detect H2S content, AtLCD enzymatic activity, and AtLCD gene expression.
For Arabidopsis culture in a solid medium, Arabidopsis seeds were treated with 10% NaClO for 10–15 min, after which the seeds were washed with aseptic water until there was no NaClO residue. The seeds were then placed at 4 °C for dark treatment for 2 to 4 days to break dormancy, sowed to a solid medium (pH = 5.8), and placed in a light incubator (22 °C, light–dark cycle 16 h/8 h).
Growth of wild-type Arabidopsis was allowed for 4–5 days in the solid medium (pH = 5.8); the seedlings were then moved to 1/2 MS solid medium, 1/2 MS solid medium with 4 mM MnCl2, 1/2 MS solid medium with 4 mM MnCl2 and 0.1 mM NaHS (Sigma, St. Louis, MO, USA), and 1/2 MS solid medium with 4 mM MnCl2 and 0.02 mM HT (Sigma, St. Louis, MO, USA). All seedlings continued to grow for 5 days and were sampled for phenotype observation, detection of the growth indicators, and H2S content.
Growth of Arabidopsis was allowed for 4–5 days in the solid medium (pH = 5.8). Wild-type, lcd, OE5, and OE32 Arabidopsis seeds were moved to 1/2 MS solid medium and 1/2 MS solid medium with 4 mM MnCl2. All seedlings continued to grow for 5 days and were sampled for the physiological index.
Wild-type, lcd, OE5, and OE32 Arabidopsis seeds (100 seedlings of each kind of Arabidopsis in each treatment) breaking dormancy were sowed on 1/2 MS solid medium, 1/2 MS solid medium containing 2 mM MnCl2, 1/2 MS solid medium containing 2 mM MnCl2 and 0.1 mM NaHS, and 1/2 MS solid medium containing 2 mM MnCl2 and 0.02 mM HT, and then the phenotypes were observed and photographed after two weeks of growth.
Wild-type, lcd, OE5, and OE32 Arabidopsis seeds were allowed for 4–5 days in the solid medium (pH = 5.8); the seedlings were moved to 1/2 MS solid medium with 4 mM MnCl2 for 5 days, the roots of Arabidopsis seedlings were sampled for Mn content and Mn-transporter-related gene expression detection.
4.3. Detection of the Growth Indicators
The main root length, which was the distance from the base of the main root to the root tip was measured. The whole seedling from the medium was dried using filter paper, and the fresh weight was measured. Then, the whole seedling was dried at 80 °C for 16 h, and the weight was measured. The determination of chlorophyll content was performed according to the method of Lichtenthaler [41].
4.4. Determination of H2S-Related Indicators
H2S content detection was performed according to the methylene blue method described in Li et al. [42]; the determination of AtLCD enzyme activity followed the method of Riemenschneider et al. [43].
4.5. Determination of Physiological Indicators
Determination of hydrogen peroxide and superoxide anion content was achieved using the method of Zhao et al. [44], and the determination of SOD, POD, and CAT activities were determined following He et al. [45]; lastly, nitrogen-blue tetrazolium (NBT) (Macklin, China) and 3, 3′-diaminobenzidine (DAB) (Macklin, Shanghai, China) staining followed Jiang et al. [46].
4.6. Hematoxylin Staining
Hematoxylin staining followed the method of Ownby (1993), with minor modifications [22]. Arabidopsis seedlings were treated with 4 mM MnCl2 for 24 h, and then the residual treatment solution was washed with deionized water. The roots of Arabidopsis seedlings were dyed in hematoxylin dye (0.2 g of hematoxylin and 0.02 g of potassium iodide, a constant volume of 0.1 L, and stored away from light) for 2 h. Then, the dye solution was washed with deionized water and observed by a microscope.
4.7. Detection of Mn Content
Mn content detection was performed using the method of inductively coupled plasma atomic emission spectrophotometry (ICP–MS), as described in Delhaize et al. [31].
4.8. RNA Extraction and qRT-PCR
Total RNA was extracted using a TRIzol reagent (Invitrogen, Waltham, MA, USA), following the manufacturer’s instructions, and the cDNA was obtained by reverse transcription using the M-MLV RT Kit (Promega, Madison, WI, USA). With β-actin as an internal reference, qRT-PCR was performed in the presence of SYBR green I (BioWhittaker Molecular Applications, Walkersville, MD, USA) in the amplification mixture, and the data were analyzed using a MyiQ Detection System. The qRT-PCR procedure included 95 °C for 5 min, 95 °C for 30 s, 58 °C for 30 s, and 72 °C for 30 s, for 40 cycles. Three replicates were run for each sample. qRT-PCR primers are shown in Table 1.

Table 1.
Primer sequences of qRT-PCR.
4.9. Statistical Methodology
Statistical analysis for all experiments was carried out using SPSS. Data were analyzed with independent t-tests (p < 0.05). All the values presented are means of replicates ± SE of three independent experiments.
5. Conclusions
In summary, H2S is involved in the response of Arabidopsis to Mn stress and may alleviate the inhibition of Mn stress on Arabidopsis seedling growth by reducing Mn2+ content, reducing reactive oxygen species content, and enhancing antioxidant enzyme activity. This study provides an important basis for further study of plant resistance to heavy metal stress.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms23095046/s1.
Author Contributions
All authors contributed to the study’s conception and design. Material preparation was performed by Z.W., G.G. and Y.Z. Data collection and analysis were performed by L.H., Q.Y. and S.L. The first draft of the manuscript was written by L.H. and Z.W. The manuscript was revised by X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (Grant Nos. 31770275, 31900235, 31701063, and 31872082).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article or Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviation
| APX | Ascorbate peroxidase |
| CAT | Catalase |
| DAB | 3, 3′-diaminobenzidine |
| H2S | Hydrogen sulfide |
| L-cysteine desulfhydrase | LCD |
| NaHS | Sodium hydrosulfide |
| Mn | Manganese |
| NBT | Nitrogen-blue tetrazolium |
| POD | Peroxides |
| ROS | Reactive oxygen species |
| SOD | Super oxide dismutase |
References
- You, X.; Yang, L.T.; Lu, Y.B.; Li, H.; Zhang, S.Q.; Chen, L.S. Proteomic changes of Citrus roots in response to long-term manganese toxicity. Trees-Struct. Funct. 2014, 28, 1383–1399. [Google Scholar] [CrossRef]
- Pittman, J.K. Managing the manganese: Molecular mechanisms of manganese transport and homeostasis. New Phytol. 2005, 167, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.; Yang, F.; Xu, C.; Yang, H.; Liu, W. Status of metal accumulation in farmland soils across China: From distribution to risk assessment. Environ. Pollut. 2013, 176, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Xue, S.; Zhu, F.; Kong, X.; Wu, C.; Huang, L.; Huang, N.; Hartley, W. A review of the characterization and revegetation of bauxite residues (Red mud). Environ. Sci. Pollut. R. 2016, 23, 1120–1132. [Google Scholar] [CrossRef]
- Weng, X.Y.; Zhao, L.L.; Zheng, C.J.; Zhu, J.W. Characteristics of the hyperaccumulator plant Phytolacca acinosa (Phytolaccaceae) in response to excess manganese. J. Plant Nutr. 2013, 36, 1355–1365. [Google Scholar] [CrossRef]
- Shi, Q.; Zhu, Z.; Xu, M.; Qian, Q.; Yu, J. Effect of excess manganese on the antioxidant system in Cucumis sativus L. under two light intensities. Environ. Exp. Bot. 2006, 58, 197–205. [Google Scholar] [CrossRef]
- Millaleo, R.; Reyes-Díaz, M.; Ivanov, A.G.; Mora, M.L.; Alberdi, M. Manganese as essential and toxic element for plants: Transport, accumulation and resistance mechanisms. J. Soil Sci. Plant Nutr. 2010, 10, 476–494. [Google Scholar] [CrossRef] [Green Version]
- Aroca, A.; Benito, J.M.; Gotor, C.; Romero, L.C. Persulfidation proteome reveals the regulation of protein function by hydrogen sulfide in diverse biological processes in Arabidopsis. J. Exp. Bot. 2017, 68, 4915–4927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Jia, Y.; Dong, R.; Huang, R.; Liu, P.; Li, X.; Wang, Z.; Liu, G.; Chen, Z. Advances in the mechanisms of plant tolerance to manganese toxicity. Int. J. Mol. Sci. 2019, 20, 5096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, L.G.; Bressan, R.A.; Filner, P. Light-dependent emission of hydrogen sulfide from plants. Plant Physiol. 1978, 61, 184–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rennenberg, H.; Arabatzis, N.; Grundel, I. Cysteine desulphydrase activity in higher plants: Evidence for the action of L- and D-cysteine specific enzymes. Phytochemistry 1987, 26, 1583–1589. [Google Scholar] [CrossRef]
- Jin, Z.; Pei, Y. Physiological implications of hydrogen sulfide in plants: Pleasant exploration behind its unpleasant odour. Oxid. Med. Cell Longev. 2015, 2015, 397502. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wu, F.H.; Wang, W.H.; Zheng, C.J.; Lin, G.H.; Dong, X.J.; He, J.X.; Pei, Z.M.; Zheng, H.L. Hydrogen sulphide enhances photosynthesis through promoting chloroplast biogenesis, photosynthetic enzyme expression, and thiol redox modification in Spinacia oleracea seedlings. J. Exp. Bot. 2011, 62, 4481–4493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Hu, S.L.; Zhang, Z.J.; Hu, L.Y.; Jiang, C.X.; Wei, Z.J.; Liu, J.; Wang, H.L.; Jiang, S.T. Hydrogen sulfide acts as a regulator of flower senescence in plants. Postharvest Biol. Tec. 2011, 60, 251–257. [Google Scholar] [CrossRef]
- Fang, H.; Jing, T.; Liu, Z.; Zhang, L.; Jin, Z.; Pei, Y. Hydrogen sulfide interacts with calcium signaling to enhance the chromium tolerance in Setaria italica. Cell Calcium 2014, 56, 472–481. [Google Scholar] [CrossRef]
- Fang, H.; Liu, Z.; Long, Y.; Liang, Y.; Jin, Z.; Zhang, L.; Liu, D.; Li, H.; Zhai, J.; Pei, Y. The Ca2+/calmodulin2-binding transcription factor TGA3 elevates LCD expression and H2S production to bolster Cr6+ tolerance in Arabidopsis. Plant J. 2017, 91, 1038–1050. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Fang, H.; Pei, Y.; Jin, Z.; Zhang, L.; Liu, D. WRKY transcription factors down-regulate the expression of H2S-generating genes, LCD and DES in Arabidopsis thaliana. Sci. Bull. 2015, 60, 995–1001. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Tan, Z.Q.; Hu, L.Y.; Wang, S.H.; Luo, J.P.; Jones, R.L. Hydrogen sulfide alleviates aluminum toxicity in germinating wheat seedlings. J. Integr. Plant Biol. 2010, 52, 556–567. [Google Scholar] [CrossRef]
- Dawood, M.; Cao, F.; Jahangir, M.M.; Zhang, G.; Wu, F. Alleviation of aluminum toxicity by hydrogen sulfide is related to elevated ATPase, and suppressed aluminum uptake and oxidative stress in barley. J. Hazard Mater. 2012, 209–210, 121–128. [Google Scholar] [CrossRef]
- Singh, V.P.; Singh, S.; Kumar, J.; Prasad, S.M. Hydrogen sulfide alleviates toxic effects of arsenate in pea seedlings through up-regulation of the ascorbate-glutathione cycle: Possible involvement of nitric oxide. J. Plant Physiol. 2015, 181, 20–29. [Google Scholar] [CrossRef]
- Fang, H.; Liu, Z.; Jin, Z.; Zhang, L.; Liu, D.; Pei, Y. An emphasis of hydrogen sulfide-cysteine cycle on enhancing the tolerance to chromium stress in Arabidopsis. Environ. Pollut. 2016, 213, 870–877. [Google Scholar] [CrossRef] [PubMed]
- Ownby, J.D. Mechanisms of reaction of hematoxylin with aluminium-treated wheat roots. Physiol. Plantarum. 1993, 87, 371–380. [Google Scholar] [CrossRef]
- Baldisserotto, C.; Ferroni, L.; Anfuso, E.; Pagnoni, A.; Fasulo, M.P.; Pancaldi, S. Responses of Trapa natans L. floating laminae to high concentrations of manganese. Protoplasma 2007, 231, 65–82. [Google Scholar] [CrossRef]
- Fang, T.; Cao, Z.; Li, J.; Shen, W.; Huang, L. Auxin-induced hydrogen sulfide generation is involved in lateral root formation in tomato. Plant Physiol. Biochem. 2014, 76, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Li, Z. Analysis of some enzymes activities of hydrogen sulfide metabolism in plants. Methods Enzymol. 2015, 555, 253–269. [Google Scholar]
- Yang, S.; Yi, K.; Chang, M.M.; Ling, G.Z.; Zhao, Z.K.; Li, X.F. Sequestration of Mn into the cell wall contributes to Mn tolerance in sugarcane (Saccharum officinarum L.). Plant Soil 2019, 436, 475–487. [Google Scholar] [CrossRef]
- Shao, J.F.; Yamaji, N.; Shen, R.F.; Ma, J.F. The key to Mn homeostasis in plants: Regulation of Mn transporters. Trends Plant Sci. 2017, 22, 215–224. [Google Scholar] [CrossRef]
- Hirschi, K.D.; Korenkov, V.D.; Wilganowski, N.L.; Wagner, G.J. Expression of Arabidopsis CAX2 in tobacco. Altered metal accumulation and increased manganese tolerance. Plant Physiol. 2000, 124, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Liang, F.; Hong, B.; Young, J.C.; Sussman, M.R.; Harper, J.F.; Sze, H. An endoplasmic reticulum-bound Ca2+/Mn2+ pump, ECA1, supports plant growth and confers tolerance to Mn2+ Stress. Plant Physiol. 2002, 130, 128–137. [Google Scholar] [CrossRef] [Green Version]
- Montanini, B.; Blaudez, D.; Jeandroz, S.; Sanders, D.; Chalot, M. Phylogenetic and functional analysis of the Cation Diffusion Facilitator (CDF) family: Improved signature and prediction of substrate specificity. BMC Genom. 2007, 8, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Delhaize, E.; Gruber, B.D.; Pittman, J.K.; White, R.G.; Leung, H.; Miao, Y.; Jiang, L.; Ryan, P.R.; Richardson, A.E. A role for the AtMTP11 gene of Arabidopsis in manganese transport and tolerance. Plant J 2007, 51, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Xie, Y. Heavy metal stress and some mechanisms of plant defense response. Scientific World J. 2015, 2015, 756120. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Li, Y.; He, L. The central role of hydrogen sulfide in plant responses to toxic metal stress. Ecotoxicol Environ. Saf. 2018, 157, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Chen, M.; Jiang, M. Hydrogen sulfide alleviates mercury toxicity by sequestering it in roots or regulating reactive oxygen species productions in rice seedlings. Plant Physiol. Biochem. 2017, 111, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Wang, X.; Dou, Y.; Liu, D.; Si, W.; Fang, H.; Zhao, C.; Chen, S.; Xi, J.; Li, J. Hydrogen sulfide-cysteine cycle system enhances cadmium tolerance through alleviating cadmium-induced oxidative stress and ion toxicity in Arabidopsis roots. Sci. Rep. 2016, 6, 39702. [Google Scholar] [CrossRef]
- Jia, H.; Yang, J.; Liu, H.; Liu, K.; Ma, P.; Chen, S.; Shi, W.; Wei, T.; Ren, X.; Guo, J.; et al. Hydrogen sulfide—Cysteine cycle plays a positive role in Arabidopsis responses to copper oxide nanoparticles stress. Environ. Exp. Bot. 2018, 155, 195–205. [Google Scholar] [CrossRef]
- Ge, S.N.; Zhao, M.M.; Wu, D.D.; Chen, Y.; Wang, Y.; Zhu, J.H.; Cai, W.J.; Zhu, Y.Z.; Zhu, Y.C. Hydrogen sulfide targets EGFR Cys797/Cys798 residues to induce Na+/K+-ATPase endocytosis and inhibition in renal tubular epithelial cells and increase sodium excretion in chronic salt-loaded rats. Antioxid Redox Signal 2014, 21, 2061–2082. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Chen, S.; Liu, D.; Liesche, J.; Shi, C.; Wang, J.; Ren, M.; Wang, X.; Yang, J.; Shi, W.; et al. Ethylene-induced hydrogen sulfide negatively regulates ethylene biosynthesis by persulfidation of ACO in tomato under osmotic stress. Front. Plant Sci. 2018, 871, 1517. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Jia, H.; Wang, X.; Shi, C.; Wang, X.; Ma, P.; Wang, J.; Ren, M.; Li, J. Hydrogen sulfide positively regulates abscisic acid signaling through persulfidation of SnRK2.6 in guard cells. Mol. Plant 2020, 13, 732–744. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic Biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Li, Z.G. Quantification of hydrogen sulfide concentration using methylene blue and 5,5′-dithiobis(2-nitrobenzoic acid) methods in plants. Methods Enzymol. 2015, 554, 101–110. [Google Scholar] [PubMed]
- Riemenschneider, A.; Nikiforova, V.; Hoefgen, R.; De Kok, L.J.; Papenbrock, J. Impact of elevated H2S on metabolite levels, activity of enzymes and expression of genes involved in cysteine metabolism. Plant Physiol. Biochem. 2005, 43, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Zhong, M.; He, L.; Wang, B.; Liu, Q.; Pan, Y.; Jiang, B.; Zhang, L. Overexpression of a chrysanthemum transcription factor gene DgNAC1 improves drought tolerance in chrysanthemum. Plant Cell Tiss. Org. 2018, 135, 119–132. [Google Scholar] [CrossRef]
- He, F.; Sheng, M.; Tang, M. Effects of rhizophagus irregularis on photosynthesis and antioxidative enzymatic system in robinia pseudoacacia L. under drought stress. Front. Plant Sci. 2017, 8, 183. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Qiu, Y.; Hu, Y.; Yu, D. Heterologous expression of at WRKY57 confers drought tolerance in oryza sativa. Front. Plant Sci. 2016, 7, 145. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).