
Bilayer Forming Phospholipids as Targets for Cancer Therapy



Abstract
:1. Introduction
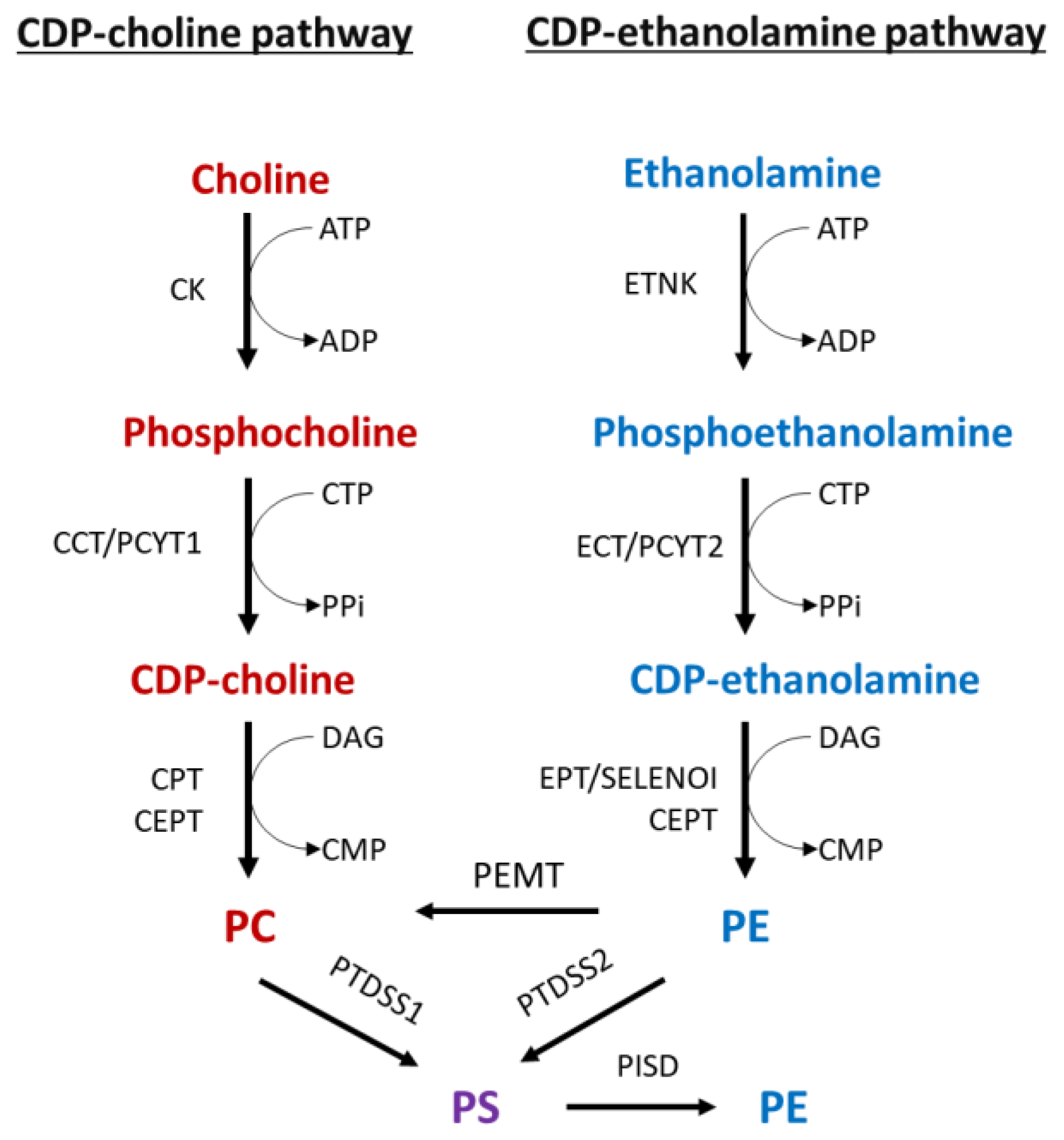
2. Phospholipid Metabolism over the Course of Cancer Progression
2.1. PC Metabolism and CKα Are Elevated in Cancers
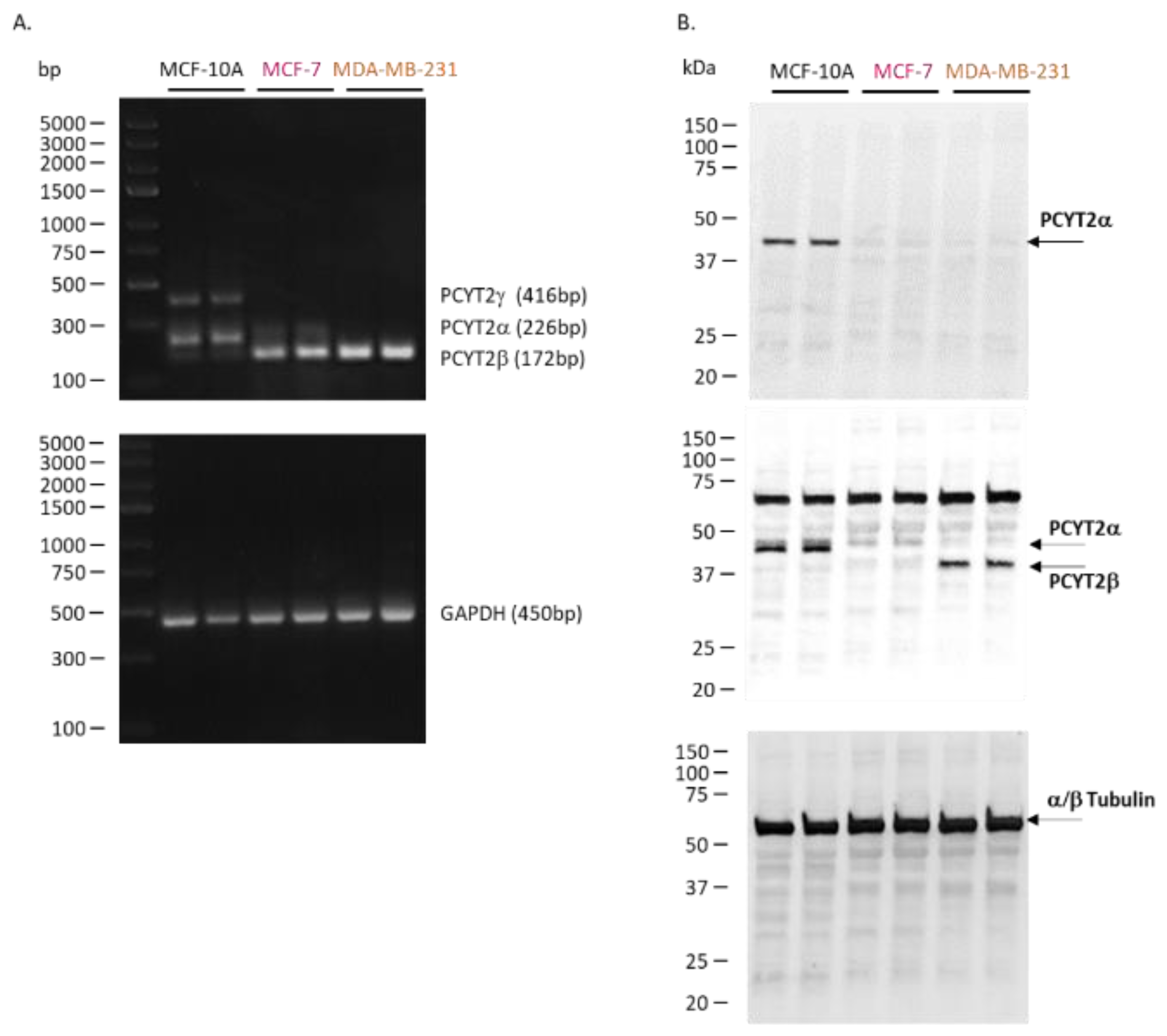
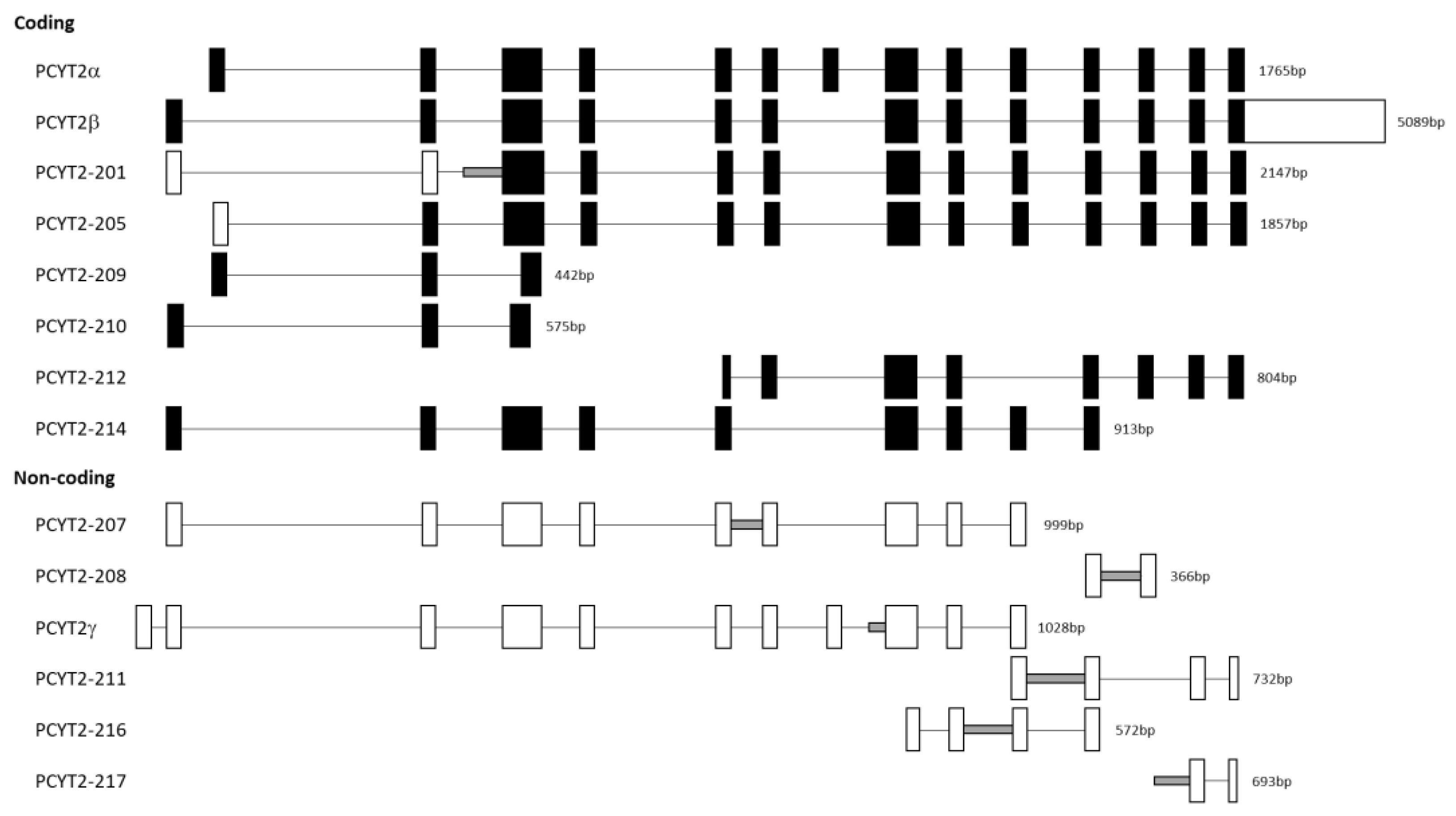
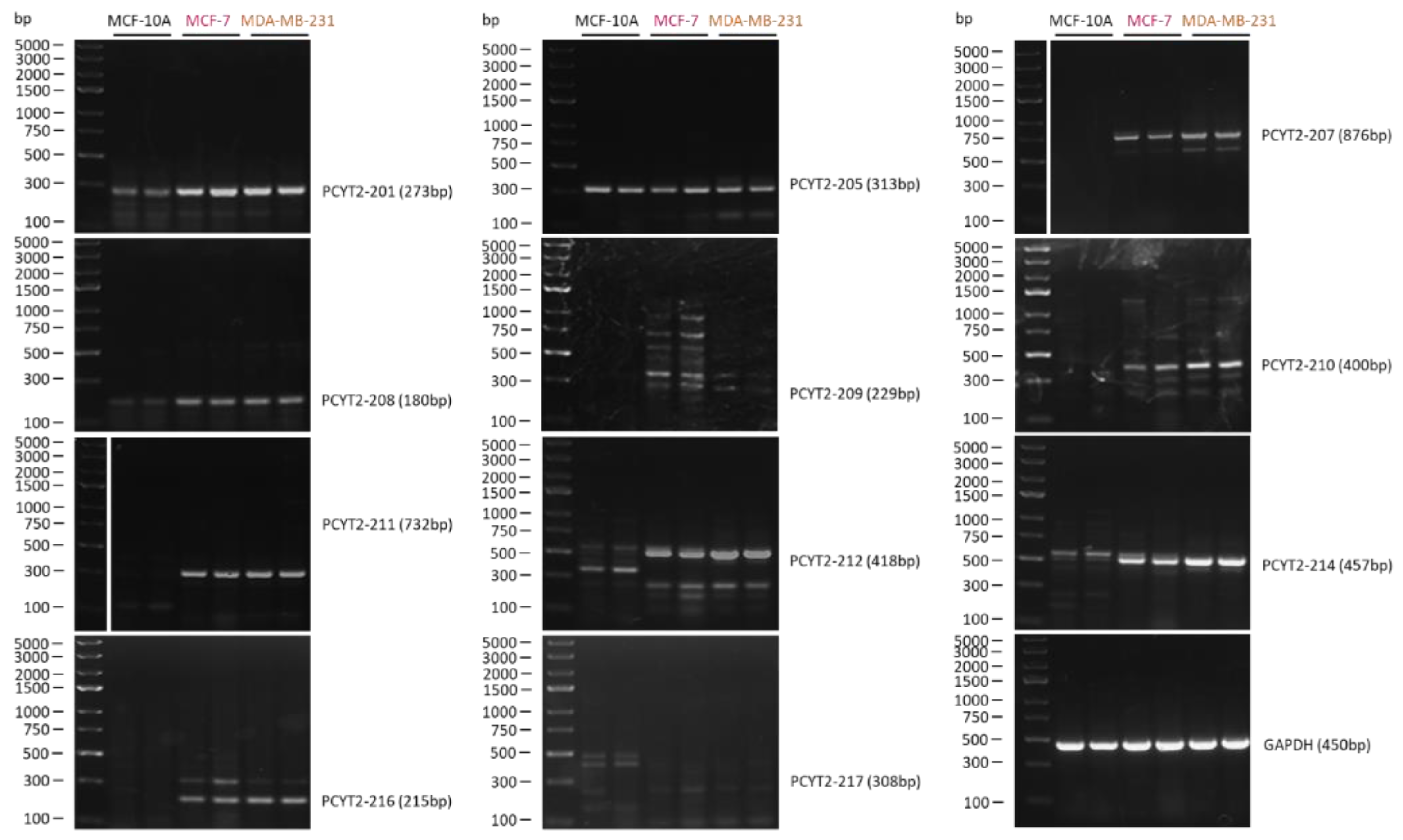
2.2. Cancer PE Metabolism Is Complexly Regulated at the Level of PCYT2
PCYT2 Alternative Splicing Is Modified in Cancer Cells and Tissues
3. Kennedy Pathway Enzymes as Prognostic Markers in Multiple Cancers
4. Phospholipid Content and Composition Can Differentiate Cancer Subtypes
4.1. Phospholipid and Fatty Acid Profiling of Cancer Cells and Tissue Biopsies
4.2. Phospholipid and Fatty Acid Profiling of Patient Plasma
5. Kennedy Pathway Inhibitors as Novel Cancer Therapeutics
5.1. Targeting the Pathway Regulators
5.2. Ethanolamine and Phosphoethanolamine
5.3. Other Targets for Cancer Therapies
5.4. Clinical Imaging
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McMaster, C.R. From yeast to humans–roles of the Kennedy pathway for phosphatidylcholine synthesis. FEBS Lett. 2018, 592, 1256–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibellini, F.; Smith, T.K. The Kennedy pathway—De novo synthesis of phosphatidylethanolamine and phosphatidylcholine. IUBMB Life 2018, 62, 414–428. [Google Scholar]
- Patel, D.; Witt, S.N. Ethanolamine and phosphatidylethanolamine: Partners in health and disease. Oxid. Med. Cell. Longev. 2017, 2017, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiedeman, A.M.; Barr, S.I.; Green, T.J.; Xu, Z.; Innis, S.M.; Kitts, D.D. Dietary choline intake: Current state of knowledge across the life cycle. Nutrients 2018, 10, 1513. [Google Scholar] [CrossRef] [Green Version]
- Zeisel, S.H.; da Costa, K.A. Choline: An essential nutrient for public health. Nutr. Rev. 2009, 67, 615–623. [Google Scholar] [CrossRef] [Green Version]
- Schenkel, L.C.; Bakovic, M. Formation and regulation of mitochondrial membranes. Int. J. Cell Biol. 2014, 2014, 709828. [Google Scholar] [CrossRef] [Green Version]
- Vance, J.E. Molecular and cell biology of phosphatidylserine and phosphatidylethanolamine metabolism. Prog. Nucleic Acid Res. Mol. Biol. 2003, 75, 69–111. [Google Scholar]
- Blunsom, N.J.; Cockcroft, S. CDP-diacylglycerol synthases (CDS): Gateway to phosphatidylinositol and cardiolipin synthesis. Front. Cell Dev. Biol. 2020, 8, 15–18. [Google Scholar] [CrossRef] [Green Version]
- Vance, J.E.; Tasseva, G. Formation and function of phosphatidylserine and phosphatidylethanolamine in mammalian cells. Biochim. Biophys. Acta 2013, 1831, 543–554. [Google Scholar] [CrossRef]
- Vance, D.E. Physiological roles of phosphatidylethanolamine N-methyltransferase. Biochim. Biophys. Acta 2013, 1831, 626–632. [Google Scholar] [CrossRef] [Green Version]
- Vance, J.E.; Vance, D.E. Phospholipid biosynthesis in mammalian cells. Biochem. Cell Biol. 2004, 82, 113–128. [Google Scholar] [CrossRef]
- Ahmed, M.Y.; Al-Khayat, A.; Al-Murshedi, F.; Al-Futaisi, A.; Chioza, B.A.; Pedro Fernandez-Murray, J.; Self, J.E.; Salter, C.G.; Harlalka, G.V.; Rawlins, L.E.; et al. A mutation of EPT1 (SELENOI) underlies a new disorder of Kennedy pathway phospholipid biosynthesis. Brain 2017, 140, 547–554. [Google Scholar]
- Henneberry, A.L.; Lagace, T.A.; Ridgway, N.D.; McMaster, C.R. Phosphatidylcholine synthesis influences the diacylglycerol homeostasis required for SEC14p-dependent Golgi function and cell growth. Mol. Biol. Cell 2001, 12, 511–520. [Google Scholar] [CrossRef] [Green Version]
- McMaster, C.R.; Bell, R.M. Phosphatidylcholine biosynthesis in Saccharomyces cerevisiae. Regulatory insights from studies employing null and chimeric sn-1,2-diacylglycerol choline- and ethanolaminephosphotransferases. J. Biol. Chem. 1994, 269, 28010–28016. [Google Scholar] [CrossRef]
- Wright, M.M.; McMaster, C.R. PC and PE synthesis: Mixed micellar analysis of the cholinephosphotransferase and ethanolaminephosphotransferase activities of human choline/ethanolamine phosphotransferase 1 (CEPT1). Lipids 2002, 37, 663–672. [Google Scholar] [CrossRef]
- Tams, R.N.; Cassilly, C.D.; Anaokar, S.; Brewer, W.T.; Dinsmore, J.T.; Chen, Y.L.; Patton-Vogt, J.; Reynolds, T.B. Overproduction of phospholipids by the Kennedy pathway leads to hypervirulence in Candida albicans. Front. Microbiol. 2019, 10, 86. [Google Scholar] [CrossRef] [Green Version]
- Vermeulen, P.S.; Geelen, M.J.H.; van Golde, L.M.G. Substrate specificity of CTP:phosphoethanolamine cytidylyltransferase purified from rat liver. Biochim. Biophys. Acta 1994, 1211, 343–349. [Google Scholar] [CrossRef]
- Pavlovic, Z.; Bakovic, M. Regulation of phosphatidylethanolamine homeostasis-the critical role of CTP:phosphoethanolamine cytidylyltransferase (Pcyt2). Int. J. Mol. Sci. 2013, 14, 2529–2550. [Google Scholar] [CrossRef] [Green Version]
- Bakovic, M.; Fullerton, M.D.; Michel, V. Metabolic and molecular aspects of ethanolamine phospholipid biosynthesis: The role of CTP:phosphoethanolamine cytidylyltransferase (Pcyt2). Biochem. Cell Biol. 2007, 85, 283–300. [Google Scholar] [CrossRef]
- Chaurio, R.A.; Janko, C.; Muñoz, L.E.; Frey, B.; Herrmann, M.; Gaipl, U.S. Phospholipids: Key players in apoptosis and immune regulation. Molecules 2009, 14, 4892–4914. [Google Scholar] [CrossRef] [Green Version]
- Crimi, M.; Esposti, M.D. Apoptosis-induced changes in mitochondrial lipids. Biochim. Biophys. Acta 2011, 1813, 551–557. [Google Scholar] [CrossRef]
- Fagerberg, C.R.; Taylor, A.; Distelmaier, F.; Schrøder, H.D.; Kibæk, M.; Wieczorek, D.; Tarnopolsky, M.; Brady, L.; Larsen, M.J.; Jamra, R.A.; et al. Choline transporter-like 1 deficiency causes a new type of childhood-onset neurodegeneration. Brain 2020, 143, 94–111. [Google Scholar] [CrossRef]
- Payne, F.; Lim, K.; Girousse, A.; Brown, R.J.; Kory, N.; Robbins, A.; Xue, Y.; Sleigh, A.; Cochran, E.; Adams, C.; et al. Mutations disrupting the Kennedy phosphatidylcholine pathway in humans with congenital lipodystrophy and fatty liver disease. Proc. Natl. Acad. Sci. USA 2014, 111, 8901–8906. [Google Scholar] [CrossRef] [Green Version]
- Basu, P.; Alibhai, F.J.; Tsimakouridze, E.V.; Singh, R.K.; Paglialunga, S.; Holloway, G.P.; Martino, T.A.; Bakovic, M. Male-specific cardiac dysfunction in CTP:phosphoethanolamine cytidylyltransferase (Pcyt2)-deficient mice. Mol. Cell. Biol. 2015, 35, 2641–2657. [Google Scholar] [CrossRef] [Green Version]
- Hofmanová, J.; Slavík, J.; Ovesná, P.; Tylichová, Z.; Dušek, L.; Straková, N.; Vaculová, A.H.; Ciganek, M.; Kala, Z.; Jíra, M.; et al. Phospholipid profiling enables to discriminate tumor- and non-tumor-derived human colon epithelial cells: Phospholipidome similarities and differences in colon cancer cell lines and in patient-derived cell samples. PLoS ONE 2020, 15, e0228010. [Google Scholar] [CrossRef] [Green Version]
- Rysman, E.; Brusselmans, K.; Scheys, K.; Timmermans, L.; Derua, R.; Munck, S.; Van Veldhoven, P.P.; Waltregny, D.; Daniëls, V.W.; Machiels, J.; et al. De novo lipogenesis protects cancer cells from free radicals and chemotherapeutics by promoting membrane lipid saturation. Cancer Res. 2010, 70, 8117–8126. [Google Scholar] [CrossRef] [Green Version]
- Marien, E.; Meister, M.; Muley, T.; Fieuws, S.; Bordel, S.; Derua, R.; Spraggins, J.; Van de Plas, R.; Dehairs, J.; Wouters, J.; et al. Non-small cell lung cancer is characterized by dramatic changes in phospholipid profiles. Int. J. Cancer 2015, 137, 1539–1548. [Google Scholar] [CrossRef]
- Jang, M.; Kim, S.S.; Lee, J. Cancer cell metabolism: Implications for therapeutic targets. Exp. Mol. Med. 2013, 45, e45. [Google Scholar] [CrossRef] [Green Version]
- Baxter, A.A.; Hulett, M.D.; Poon, I.K.H. The phospholipid code: A key component of dying cell recognition, tumor progression and host-microbe interactions. Cell Death Differ. 2015, 22, 1893–1905. [Google Scholar] [CrossRef] [Green Version]
- Snaebjornsson, M.T.; Janaki-Raman, S.; Schulze, A. Greasing the wheels of the cancer machine: The role of lipid metabolism in cancer. Cell Metab. 2020, 31, 62–76. [Google Scholar] [CrossRef]
- Punnonen, K.; Hietanen, E.; Auvinen, O.; Punnonen, R. Phospholipids and fatty acids in breast cancer tissue. J. Cancer Res. Clin. Oncol. 1989, 115, 575–578. [Google Scholar] [CrossRef] [PubMed]
- Azordegan, N.; Fraser, V.; Le, K.; Hillyer, L.M.; Ma, D.W.L.; Fischer, G.; Moghadasian, M.H. Carcinogenesis alters fatty acid profile in breast tissue. Mol. Cell. Biochem. 2013, 374, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Munir, R.; Lisec, J.; Swinnen, J.V.; Zaidi, N. Lipid metabolism in cancer cells under metabolic stress. Br. J. Cancer 2019, 120, 1090–1098. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Cheng, C.; Tan, Z.; Li, N.; Tang, M.; Yang, L.; Cao, Y. Emerging roles of lipid metabolism in cancer metastasis. Mol. Cancer 2017, 16, 76. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.K.; Lee, H.S.; Park, Y.S.; Lee, J.H.; Lee, S.C.; Lee, J.H.; Lee, S.J.; Shanta, S.R.; Park, H.M.; Kim, H.R.; et al. Lipid MALDI profile classifies non-small cell lung cancers according to the histologic type. Lung Cancer 2012, 76, 197–203. [Google Scholar] [CrossRef]
- Pakiet, A.; Kobiela, J.; Stepnowski, P.; Sledzinski, T.; Mika, A. Changes in lipids composition and metabolism in colorectal cancer: A review. Lipids Health Dis. 2019, 18, 29. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.A.D.; Bush, C.; Jameson, C.; Titley, J.C.; Leach, M.O.; Wilman, D.E.V.; McCready, V.R. Phospholipid metabolites, prognosis and proliferation in human breast carcinoma. NMR Biomed. 1993, 6, 318–323. [Google Scholar] [CrossRef]
- Mistry, D.A.H.; French, P.W. Circulating phospholipids as biomarkers of breast cancer: A review. Breast Cancer Basic Clin. Res. 2016, 10, 191–196. [Google Scholar] [CrossRef]
- Shin, H.W.; Takatsu, H. Phosphatidylserine exposure in living cells. Crit. Rev. Biochem. Mol. Biol. 2020, 55, 166–178. [Google Scholar] [CrossRef]
- Dayoub, A.S.; Brekken, R.A. TIMs, TAMs, and PS- antibody targeting: Implications for cancer immunotherapy. Cell Commun. Signal. 2020, 18, 29. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.; Fa, H.; Xiao, D.; Wang, J. Targeting phosphatidylserine for cancer therapy: Prospects and challenges. Theranostics 2020, 10, 9214–9229. [Google Scholar] [CrossRef]
- Belzile, O.; Huang, X.; Gong, J.; Carlson, J.; Schroit, A.J.; Brekken, R.A.; Freimark, B.D. Antibody targeting of phosphatidylserine for the detection and immunotherapy of cancer. Immunotargets Ther. 2018, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Gozzelino, L.; De Santis, M.C.; Gulluni, F.; Hirsch, E.; Martini, M. PI (3,4)P2 signaling in cancer and metabolism. Front. Oncol. 2020, 10, e360. [Google Scholar] [CrossRef] [Green Version]
- Ijuin, T. Phosphoinositide phosphatases in cancer cell dynamics—beyond PI3K and PTEN. Semin. Cancer Biol. 2019, 59, 50–65. [Google Scholar] [CrossRef]
- Owusu Obeng, E.; Rusciano, I.; Marvi, M.V.; Fazio, A.; Ratti, S.; Follo, M.Y.; Xian, J.; Manzoli, L.; Billi, A.M.; Mongiorgi, S.; et al. Phosphoinositide-dependent signaling in cancer: A focus on phospholipase C isozymes. Int. J. Mol. Sci. 2020, 21, 2581. [Google Scholar] [CrossRef] [Green Version]
- Kurabe, N.; Hayasaka, T.; Ogawa, M.; Masaki, N.; Ide, Y.; Waki, M.; Nakamura, T.; Kurachi, K.; Kahyo, T.; Shinmura, K.; et al. Accumulated phosphatidylcholine (16:0/16:1) in human colorectal cancer; possible involvement of LPCAT4. Cancer Sci. 2013, 104, 1295–1302. [Google Scholar] [CrossRef]
- Ackerstaff, E.; Glunde, K.; Bhujwalla, Z.M. Choline phospholipid metabolism: A target in cancer cells? J. Cell. Biochem. 2003, 90, 525–533. [Google Scholar] [CrossRef]
- Sonkar, K.; Ayyappan, V.; Tressler, C.M.; Adelaja, O.; Cai, R.; Cheng, M.; Glunde, K. Focus on the glycerophosphocholine pathway in choline phospholipid metabolism of cancer. NMR Biomed. 2019, 32, 1–33. [Google Scholar] [CrossRef]
- Shah, T.; Wildes, F.; Penet, M.-F.; Winnard Jr, P.T.; Glunde, K.; Artemov, D.; Ackerstaff, E.; Gimi, B.; Kakkad, S.; Raman, V.; et al. Choline kinase overexpression increases invasiveness and drug resistance of human breast cancer cells. NMR Biomed. 2010, 23, 633–642. [Google Scholar] [CrossRef] [Green Version]
- Falcon, S.C.; Hudson, C.S.; Huang, Y.; Mortimore, M.; Golec, J.M.; Charlton, P.A.; Weber, P.; Sundaram, H. A non-catalytic role of choline kinase alpha is important in promoting cancer cell survival. Oncogenesis 2013, 2, e38. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Wang, R.Y.; Cai, J.; Feng, D.; Yang, G.Z.; Xu, Q.G.; Zhai, Y.X.; Zhang, Y.; Zhou, W.P.; Cai, Q.P. Overexpression of CHKA contributes to tumor progression and metastasis and predicts poor prognosis in colorectal carcinoma. Oncotarget 2016, 7, 66660–66678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Human Protein Atlas. CHKA. 2021. Available online: https://www.proteinatlas.org/ (accessed on 12 April 2021).
- Osawa, T.; Shimamura, T.; Saito, K.; Hasegawa, Y.; Ishii, N.; Nishida, M.; Ando, R.; Kondo, A.; Anwar, M.; Tsuchida, R.; et al. Phosphoethanolamine accumulation protects cancer cells under glutamine starvation through downregulation of PCYT2. Cell Rep. 2019, 29, 89–103.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Johnson, C.; Bakovic, M. Stimulation of the human CTP:phosphoethanolamine cytidylyltransferase gene by early growth response protein 1. J. Lipid Res. 2008, 49, 2197–2211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Bakovic, M. Breast cancer cells adapt to metabolic stress by increasing ethanolamine phospholipid synthesis and CTP:ethanolaminephosphate cytidylyltransferase-Pcyt2 activity. Biochem. Cell Biol. 2012, 90, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Imae, R.; Manya, H.; Tsumoto, H.; Miura, Y.; Endo, T. PCYT2 synthesizes CDP-glycerol in mammals and reduced PCYT2 enhances the expression of functionally glycosylated α-dystroglycan. J. Biochem. 2021, 170, 183–194. [Google Scholar] [CrossRef]
- Imae, R.; Manya, H.; Tsumoto, H.; Osumi, K.; Tanaka, T.; Mizuno, M.; Kanagawa, M.; Kobayashi, K.; Toda, T.; Endo, T. CDP-glycerol inhibits the synthesis of the functional O-mannosyl glycan of α-dystroglycan. J. Biol. Chem. 2018, 293, 12186–12198. [Google Scholar] [CrossRef] [Green Version]
- Sgambato, A.; Migaldi, M.; Montanari, M.; Camerini, A.; Brancaccio, A.; Rossi, G.; Cangiano, R.; Losasso, C.; Capelli, G.; Trentini, G.P.; et al. Dystroglycan expression is frequently reduced in human breast and colon cancers and is associated with tumor progression. Am. J. Pathol. 2003, 162, 849–860. [Google Scholar] [CrossRef] [Green Version]
- Qi, F.; Li, Y.; Yang, X.; Wu, Y.P.; Lin, L.J.; Liu, X.M. Significance of alternative splicing in cancer cells. Chin. Med. J. 2020, 133, 221–228. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, J.; Huang, B.O.; Xu, Y.M.; Li, J.; Huang, L.F.; Lin, J.; Zhang, J.; Min, Q.H.; Yang, W.M.; et al. Mechanism of alternative splicing and its regulation. Biomed. Rep. 2015, 3, 152–158. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Wang, J.; Zhu, X.; Chyr, J.; Zhou, X.; Wu, X.; Huang, L. The functional impact of alternative splicing on the survival prognosis of triple-negative breast cancer. Front. Genet. 2021, 11, 604262. [Google Scholar] [CrossRef]
- Pavlovic, Z.; Zhu, L.; Pereira, L.; Singh, R.K.; Cornel, R.B.; Bakovic, M. Isoform-specific and protein kinase C-mediated regulation of CTP:phosphoethanolamine cytidylyltransferase phosphorylation. J. Biol. Chem. 2014, 289, 9053–9064. [Google Scholar] [CrossRef] [Green Version]
- Tie, A.; Bakovic, M. Alternative splicing of CTP:phosphoethanolamine cytidylyltransferase produces two isoforms that differ in catalytic properties. J. Lipid Res. 2007, 48, 2172–2181. [Google Scholar] [CrossRef] [Green Version]
- Human Protein Atlas. PCYT2. 2021. Available online: https://www.proteinatlas.org/ (accessed on 12 April 2021).
- Vaz, F.M.; McDermott, J.H.; Alders, M.; Wortmann, S.B.; Kölker, S.; Pras-Raves, M.L.; Vervaart, M.A.T.; van Lenthe, H.; Luyf, A.C.M.; Elfrink, H.L.; et al. Mutations in PCYT2 disrupt etherlipid biosynthesis and cause a complex hereditary spastic paraplegia. Brain 2019, 142, 3382–3397. [Google Scholar] [CrossRef] [Green Version]
- Vaz, F.M.; McDermott, J.H.; Engelen, M.; Banka, S. Reply: Expanding the clinical and genetic spectrum of PCYT2-related disorders. Brain 2020, 143, e77. [Google Scholar] [CrossRef]
- Vélez-Santamaría, V.; Verdura, E.; Macmurdo, C.; Planas-Serra, L.; Schlüter, A.; Casas, J.; Martínez, J.J.; Casasnovas, C.; Si, Y.; Thompson, S.S.; et al. Expanding the clinical and genetic spectrum of PCYT2-related disorders. Brain 2020, 143, e76. [Google Scholar] [CrossRef]
- Kurosaki, T.; Maquat, L.E. Nonsense-mediated mRNA decay in humans at a glance. J. Cell Sci. 2016, 129, 461–467. [Google Scholar] [CrossRef] [Green Version]
- Hug, N.; Longman, D.; Cáceres, J.F. Mechanism and regulation of the nonsense-mediated decay pathway. Nucleic Acids Res. 2016, 44, 1483–1495. [Google Scholar] [CrossRef] [Green Version]
- Kovalak, C.; Donovan, S.; Bicknell, A.A.; Metkar, M.; Moore, M.J. Deep sequencing of pre-translational mRNPs reveals hidden flux through evolutionarily conserved alternative splicing nonsense-mediated decay pathways. Genome Biol. 2021, 22, 132. [Google Scholar] [CrossRef]
- Klupczynska, A.; Plewa, S.; Kasprzyk, M.; Dyszkiewicz, W.; Kokot, Z.J.; Matysiak, J. Serum lipidome screening in patients with stage I non-small cell lung cancer. Clin. Exp. Med. 2019, 19, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Zinrajh, D.; Hörl, G.; Jürgens, G.; Marc, J.; Sok, M.; Cerne, D. Increased phosphatidylethanolamine N-methyltransferase gene expression in non-small-cell lung cancer tissue predicts shorter patient survival. Oncol. Lett. 2014, 7, 2175–2179. [Google Scholar] [CrossRef] [Green Version]
- Glunde, K.; Bhujwalla, Z.M.; Ronen, S.M. Choline metabolism in malignant transformation. Nat. Rev. Cancer 2011, 11, 835–848. [Google Scholar] [CrossRef] [Green Version]
- de Molina, A.R.; Sarmentero-Estrada, J.; Belda-Iniesta, C.; Tarón, M.; de Molina, V.R.; Cejas, P.; Skrzypski, M.; Gallego-Ortega, D.; de Castro, J.; Casado, E.; et al. Expression of choline kinase alpha to predict outcome in patients with early-stage non-small-cell lung cancer: A retrospective study. Lancet Oncol. 2007, 8, 889–897. [Google Scholar] [CrossRef]
- Han, X.; Tan, Q.; Yang, S.; Li, J.; Xu, J.; Hao, X.; Hu, X.; Xing, P.; Liu, Y.; Lin, L.; et al. Comprehensive profiling of gene copy number alterations predicts patient prognosis in resected stages I–III lung adenocarcinoma. Front. Oncol. 2019, 9, 556. [Google Scholar] [CrossRef] [Green Version]
- Lesko, J.; Triebl, A.; Stacher-Priehse, E.; Fink-Neuböck, N.; Lindenmann, J.; Smolle-Jüttner, F.M.; Köfeler, H.C.; Hrzenjak, A.; Olschewski, H.; Leithner, K. Phospholipid dynamics in ex vivo lung cancer and normal lung explants. Exp. Mol. Med. 2021, 53, 81–90. [Google Scholar] [CrossRef]
- Berglund, L.; Björling, E.; Oksvold, P.; Fagerberg, L.; Asplund, A.; Szigyarto, C.A.-K.; Persson, A.; Ottosson, J.; Wernérus, H.; Nilsson, P.; et al. A genecentric Human Protein Atlas for expression profiles based on antibodies. Mol. Cell. Proteom. 2008, 7, 2019–2027. [Google Scholar] [CrossRef] [Green Version]
- Human Protein Atlas. PEMT. Available online: https://www.proteinatlas.org/ (accessed on 12 April 2021).
- Human Protein Atlas. EPT1. Available online: https://www.proteinatlas.org/ (accessed on 12 April 2021).
- Human Protein Atlas. SELENOI. Available online: https://www.proteinatlas.org/ (accessed on 12 April 2021).
- Urbanelli, L.; Buratta, S.; Logozzi, M.; Mitro, N.; Sagini, K.; Di Raimo, R.; Caruso, D.; Fais, S.; Emiliani, C. Lipidomic analysis of cancer cells cultivated at acidic pH reveals phospholipid fatty acids remodelling associated with transcriptional reprogramming. J. Enzym. Inhib. Med. Chem. 2020, 35, 963–973. [Google Scholar] [CrossRef]
- Koundouros, N.; Poulogiannis, G. Reprogramming of fatty acid metabolism in cancer. Br. J. Cancer 2020, 122, 4–22. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Xie, F.; Yang, Y.; Wang, S. Reprogramming of fatty acid metabolism in breast cancer: A narrative review. Transl. Breast Cancer Res. 2021, 2. [Google Scholar] [CrossRef]
- Fichtali, K.; Bititi, A.; Elghanmi, A.; Ghazi, B. Serum lipidomic profiling in breast cancer to identify screening, diagnostic, and prognostic biomarkers. Bioresearch 2020, 9, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Pickens, C.A.; Lane-Elliot, A.; Comstock, S.S.; Fenton, J.I. Altered saturated and monounsaturated plasma phospholipid fatty acid profiles in adult males with colon adenomas. Am. Assoc. Cancer Res. 2016, 25, 498–506. [Google Scholar] [CrossRef] [Green Version]
- Baró, L.; Hermoso, J.C.; Núñez, M.C.; Jiménez-Rios, J.A.; Gil, A. Abnormalities in plasma and red blood cell fatty acid profiles of patients with colorectal cancer. Br. J. Cancer 1998, 77, 1978–1983. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Díaz-Beltrán, L.; González-Olmedo, C.; Luque-Caro, N.; Díaz, C.; Martín-Blázquez, A.; Fernández-Navarro, M.; Ortega-Granados, A.L.; Gálvez-Montosa, F.; Vicente, F.; Del Palacio, J.P.; et al. Human plasma metabolomics for biomarker discovery: Targeting the molecular subtypes in breast cancer. Cancers 2021, 13, 147. [Google Scholar] [CrossRef] [PubMed]
- Cava, C.; Pisati, M.; Frasca, M.; Castiglioni, I. Identification of breast cancer subtype-specific biomarkers by integrating copy number alterations and gene expression profiles. Medicina 2021, 57, 261. [Google Scholar] [CrossRef] [PubMed]
- Yonekubo, Y.; Wu, P.; Esechie, A.; Zhang, Y.; Du, G. Characterization of new serum biomarkers in breast cancer using lipid microarrays. Tumor Biol. 2010, 31, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S. Empowering clinical diagnostics with mass spectrometry. ACS Omega 2020, 5, 2041–2048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jelonek, K.; Ros, M.; Pietrowska, M.; Widlak, P. Cancer biomarkers and mass spectrometry-based analyses of phospholipids in body fluids. Clin. Lipidol. 2013, 8, 137–150. [Google Scholar] [CrossRef]
- Hernández-Alcoceba, R.; Fernández, F.; Lacal, J.C. In vivo antitumor activity of choline kinase inhibitors. Cancer Res. 1999, 59, 3112–3118. [Google Scholar]
- Arlauckas, S.P.; Popov, A.V.; Delikatny, E.J. Choline kinase alpha-putting the ChoK-hold on tumor metabolism. Prog. Lipid Res. 2016, 63, 28–40. [Google Scholar] [CrossRef] [Green Version]
- Trousil, S.; Kaliszczak, M.; Schug, Z.; Nguyen, Q.-D.; Tomasi, G.; Favicchio, R.; Brickute, D.; Fortt, R.; Twyman, F.J.; Carroll, L.; et al. The novel choline kinase inhibitor ICL-CCIC-0019 reprograms cellular metabolism and inhibits cancer cell growth. Oncotarget 2016, 7, 37103–37120. [Google Scholar] [CrossRef] [Green Version]
- Glunde, K.; Ackerstaff, E.; Mori, N.; Jacobs, M.A.; Bhujwalla, Z.M. Choline phospholipid metabolism in cancer: Consequences for molecular pharmaceutical interventions. Mol. Pharm. 2006, 3, 496–506. [Google Scholar] [CrossRef]
- Inazu, M.; Yamada, T.; Kubota, N.; Yamanaka, T. Functional expression of choline transporter-like protein 1 (CTL1) in small cell lung carcinoma cells: A target molecule for lung cancer therapy. Pharmacol. Res. 2013, 76, 119–131. [Google Scholar] [CrossRef] [Green Version]
- Cheng, M.; Bhujwalla, Z.M.; Glunde, K. Targeting phospholipid metabolism in cancer. Front. Oncol. 2016, 6, 00266. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.; Grapentine, S.; Ichhpuniani, J.; Bakovic, M. Choline transporter-like proteins 1 and 2 are newly identified plasma membrane and mitochondrial ethanolamine transporters. J. Biol. Chem. 2021, 296, 100604. [Google Scholar] [CrossRef]
- Inazu, M. Choline transporter-like proteins CTLs/SLC44 family as a novel molecular target for cancer therapy. Biopharm. Drug Dispos. 2014, 35, 431–449. [Google Scholar] [CrossRef]
- Guan, Y.; Chen, X.; Wu, M.; Zhu, W.; Arslan, A.; Takeda, S.; Nguyen, M.H.; Majeti, R.; Thomas, D.; Zheng, M.; et al. The phosphatidylethanolamine biosynthesis pathway provides a new target for cancer chemotherapy. J. Hepatol. 2020, 72, 746–760. [Google Scholar] [CrossRef]
- Shibata, K.; Nishijima, N.; Hirai, K.; Watanabe, S.; Yamanaka, T.; Chikazu, D.; Inazu, M. A novel plant-derived choline transporter-like protein 1 inhibitor, Amb544925, induces apoptotic cell Death via the ceramide/survivin pathway in tongue squamous cell carcinoma. Cancers 2022, 14, 329. [Google Scholar] [CrossRef]
- Gupta, D.; Bhattacharjee, O.; Mandal, D.; Sen, M.K.; Dey, D.; Dasgupta, A.; Kazi, T.A.; Gupta, R.; Sinharoy, S.; Acharya, K.; et al. CRISPR-Cas9 system: A new-fangled dawn in gene editing. Life Sci. 2019, 232, 116636. [Google Scholar] [CrossRef]
- Dalaklioglu, S.; Sahin, P.; Ordueri, E.G.; Celik-Ozenci, C.; Tasatargil, A. Potential role of poly (ADP-ribose) polymerase (PARP) activation in methotrexate-induced nephrotoxicity and tubular apoptosis. Int. J. Toxicol. 2012, 31, 430–440. [Google Scholar] [CrossRef]
- Michels, J.; Kepp, O.; Senovilla, L.; Lissa, D.; Castedo, M.; Kroemer, G.; Galluzzi, L. Functions of BCL-XL at the interface between cell death and metabolism. Int. J. Cell Biol. 2013, 2013, 705294. [Google Scholar] [CrossRef] [Green Version]
- Saxena, R.; Yang, C.; Rao, M.; Turaga, R.C.; Garlapati, C.; Gundala, S.R.; Myers, K.; Ghareeb, A.; Bhattarai, S.; Kamalinia, G.; et al. Preclinical development of a nontoxic oral formulation of monoethanolamine, a lipid precursor, for prostate cancer treatment. Clin. Cancer Res. 2017, 23, 3781–3793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, A.K.; Meneguelo, R.; Pereira, A.; Filho, O.M.R.; Chierice, G.O.; Maria, D.A. Anticancer effects of synthetic phosphoethanolamine on Ehrlich ascites tumor: An experimental study. Anticancer. Res. 2012, 32, 95–104. [Google Scholar] [PubMed]
- Ferreira, A.K.; Freitas, V.M.; Levy, D.; Ruiz, J.L.M.; Bydlowski, S.P.; Rici, R.E.G.; Filho, O.M.R.; Chierice, G.O.; Maria, D.A. Anti-angiogenic and anti-metastatic activity of synthetic phosphoethanolamine. PLoS ONE 2013, 8, e57937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, A.K.; Meneguelo, R.; Pereira, A.; Filho, O.M.R.; Chierice, G.O.; Maria, D.A. Synthetic phosphoethanolamine induces cell cycle arrest and apoptosis in human breast cancer MCF-7 cells through the mitochondrial pathway. Biomed. Pharmacother. 2013, 67, 481–487. [Google Scholar] [CrossRef]
- Ferreira, A.K.; Meneguelo, R.; Marques, F.L.N.; Radin, A.; Filho, O.M.R.; Neto, S.C.; Chierice, G.O.; Maria, D.A. Synthetic phosphoethanolamine a precursor of membrane phospholipids reduce tumor growth in mice bearing melanoma B16-F10 and in vitro induce apoptosis and arrest in G2/M phase. Biomed. Pharmacother. 2012, 66, 541–548. [Google Scholar] [CrossRef]
- Fontana, D.; Mauri, M.; Renso, R.; Docci, M.; Crespiatico, I.; Røst, L.M.; Jang, M.; Niro, A.; D’Aliberti, D.; Massimino, L.; et al. ETNK1 mutations induce a mutator phenotype that can be reverted with phosphoethanolamine. Nat. Commun. 2020, 11, 5938. [Google Scholar] [CrossRef]
- Casares, D.; Escribá, P.V.; Rosselló, C.A. Membrane lipid composition: Effect on membrane and organelle structure, function and compartmentalization and therapeutic avenues. Int. J. Mol. Sci. 2019, 20, 2167. [Google Scholar] [CrossRef] [Green Version]
- Martin, M.L.; Barceló-Coblijn, G.; de Almeida, R.F.M.; Noguera-Salvà, M.A.; Terés, S.; Higuera, M.; Liebisch, G.; Schmitz, G.; Busquets, X.; Escribá, P.V. The role of membrane fatty acid remodeling in the antitumor mechanism of action of 2-hydroxyoleic acid. Biochim. Biophys. Acta 2013, 1828, 1405–1413. [Google Scholar] [CrossRef] [Green Version]
- Terés, S.; Lladó, V.; Higuera, M.; Barceló-Coblijn, G.; Martin, M.L.; Noguera-Salvà, M.A.; Marcilla-Etxenike, A.; García-Verdugo, J.M.; Soriano-Navarro, M.; Saus, C.; et al. 2-Hydroxyoleate, a nontoxic membrane binding anticancer drug, induces glioma cell differentiation and autophagy. Proc. Natl. Acad. Sci. USA 2012, 109, 8489–8494. [Google Scholar] [CrossRef] [Green Version]
- Marcilla-Etxenike, A.; Martín, M.L.; Noguera-Salvà, M.A.; García-Verdugo, J.M.; Soriano-Navarro, M.; Dey, I.; Escribá, P.V.; Busquets, X. 2-Hydroxyoleic acid induces ER stress and autophagy in various human glioma cell lines. PLoS ONE 2012, 7, e48235. [Google Scholar]
- Chidley, C.; Trauger, S.A.; Birsoy, K.; O’Shea, E.K. The anticancer natural product ophiobolin A induces cytotoxicity by covalent modification of phosphatidylethanolamine. eLife 2016, 5, e14601. [Google Scholar] [CrossRef]
- Preta, G. New insights into targeting membrane lipids for cancer therapy. Front. Cell Dev. Biol. 2020, 8, 876. [Google Scholar] [CrossRef]
- Tan, L.T.H.; Chan, K.G.; Pusparajah, P.; Lee, W.L.; Chuah, L.H.; Khan, T.M.; Lee, L.H.; Goh, B.H. Targeting membrane lipids: A potential cancer cure? Front. Pharmacol. 2017, 8, 12. [Google Scholar] [CrossRef]
- Jaswal, A.P.; Hazari, P.P.; Prakash, S.; Sethi, P.; Kaushik, A.; Roy, B.G.; Kathait, S.; Singh, B.; Mishra, A.K. [99mTc]Tc-DTPA-Bis(cholineethylamine) as an oncologic tracer for the detection of choline transporter (ChT) and choline kinase (ChK) expression in cancer. ACS Omega 2022, 55, 48–50. [Google Scholar] [CrossRef]
- Giovacchini, G.; Guglielmo, P.; Mapelli, P.; Incerti, E.; Gajate, A.M.S.; Giovannini, E.; Riondato, M.; Briganti, A.; Gianolli, L.; Ciarmiello, A.; et al. 11C-choline PET/CT predicts survival in prostate cancer patients with PSA <1ngml. Eur. J. Nucl. Med. Mol. Imaging 2019, 46, 921–929. [Google Scholar]
- Bagnoli, M.; Granata, A.; Nicoletti, R.; Krishnamachary, B.; Bhujwalla, Z.M.; Canese, R.; Podo, F.; Canevari, S.; Iorio, E.; Mezzanzanica, D. Choline metabolism alteration: A focus on ovarian cancer. Front. Oncol. 2016, 6, 156. [Google Scholar] [CrossRef] [Green Version]
- Abrantes, A.M.; Pires, A.S.; Monteiro, L.; Teixo, R.; Neves, A.R.; Tavares, N.T.; Marques, I.A.; Botelho, M.F. Tumour functional imaging by PET. Biochim. Biophys. Acta 2020, 1866, 165717. [Google Scholar] [CrossRef]
- Filippi, L.; Schillaci, O.; Bagni, O. Recent advances in PET probes for hepatocellular carcinoma characterization. Expert Rev. Med. Devices 2019, 16, 341–350. [Google Scholar] [CrossRef]
- Contractor, K.; Challapalli, A.; Barwick, T.; Winkler, M.; Hellawell, G.; Hazell, S.; Tomasi, G.; Al-Nahhas, A.; Mapelli, P.; Kenny, L.M.; et al. Use of [11C] choline PET-CT as a noninvasive method for detecting pelvic lymph node status from prostate cancer and relationship with choline kinase expression. Clin. Cancer Res. 2011, 17, 7673–7683. [Google Scholar] [CrossRef] [Green Version]
- Trousil, S.; Lee, P.; Pinato, D.J.; Ellis, J.K.; Dina, R.; Aboagye, E.O.; Keun, H.C.; Sharma, R. Alterations of choline phospholipid metabolism in endometrial cancer are caused by choline kinase alpha overexpression and a hyperactivated deacylation pathway. Cancer Res. 2014, 74, 6867–6877. [Google Scholar] [CrossRef] [Green Version]
- Arlauckas, S.P.; Kumar, M.; Popov, A.V.; Poptani, H.; Delikatny, E.J. Near infrared fluorescent imaging of choline kinase alpha expression and inhibition in breast tumors. Oncotarget 2017, 8, 16518–16530. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Upregulated Variants | Fold-Change Relative to Control | |||
| MCF-10A | MCF-7 | MDA-MB-231 | ||
| Coding | PCYT2-201 | 1 | 8.73 | 8.32 |
| Non-coding | PCYT2β | 1 | 53 * | 99 * |
| PCYT2-209 | 0 | 2074 ** | 2925 ** | |
| PCYT2-210 | 0 | 2840 * | 2160 * | |
| PCYT2-212 | 1 | 39.5 * | 68.0 | |
| PCYT2-214 | 1 | 34.5 * | 59.6 * | |
| PCYT2-207 | 0 | 1620 * | 3380 * | |
| PCYT2-208 | 1 | 3.47 * | 3.99 * | |
| PCYT2-211 | 1 | 36.5 ** | 51.8 * | |
| PCYT2-216 | 0 | 1889 ** | 2792 ** † | |
| Downregulated variants | Fold-change relative to control | |||
| MCF-10A | MCF-7 | MDA-MB-231 | ||
| Coding | PCYT2α | 1 | 0.04 ** | 0 ** † |
| PCYT2-205 | 1 | 0.92 | 0.87 | |
| Non-coding | PCYT2-213/PCYT2γ | 1 | 0 ** | 0 ** |
| Protein | Role | Cancer Type | Outcome | % of Patients with Increased Expression |
|---|---|---|---|---|
| CKα | Phosphorylation of choline | Liver cancer | Unfavorable | 16 |
| ETNK1 | Phosphorylation of ethanolamine | Urothelial cancer Liver cancer | Favorable Unfavorable | 25 16–33 |
| PCYT2 | Formation of CDP-ethanolamine | Renal cancer | Favorable | 33–36 |
| PEMT | Methylation of PE to PC | Endometrial cancer | Favorable | 33 |
| SELENO1 | Formation of PE | No | N/A | N/A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stoica, C.; Ferreira, A.K.; Hannan, K.; Bakovic, M. Bilayer Forming Phospholipids as Targets for Cancer Therapy. Int. J. Mol. Sci. 2022, 23, 5266. https://doi.org/10.3390/ijms23095266
Stoica C, Ferreira AK, Hannan K, Bakovic M. Bilayer Forming Phospholipids as Targets for Cancer Therapy. International Journal of Molecular Sciences. 2022; 23(9):5266. https://doi.org/10.3390/ijms23095266
Chicago/Turabian StyleStoica, Celine, Adilson Kleber Ferreira, Kayleigh Hannan, and Marica Bakovic. 2022. "Bilayer Forming Phospholipids as Targets for Cancer Therapy" International Journal of Molecular Sciences 23, no. 9: 5266. https://doi.org/10.3390/ijms23095266
APA StyleStoica, C., Ferreira, A. K., Hannan, K., & Bakovic, M. (2022). Bilayer Forming Phospholipids as Targets for Cancer Therapy. International Journal of Molecular Sciences, 23(9), 5266. https://doi.org/10.3390/ijms23095266