
Primary Focal Segmental Glomerulosclerosis Plasmas Increase Lipid Droplet Formation and Perilipin-2 Expression in Human Podocytes
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
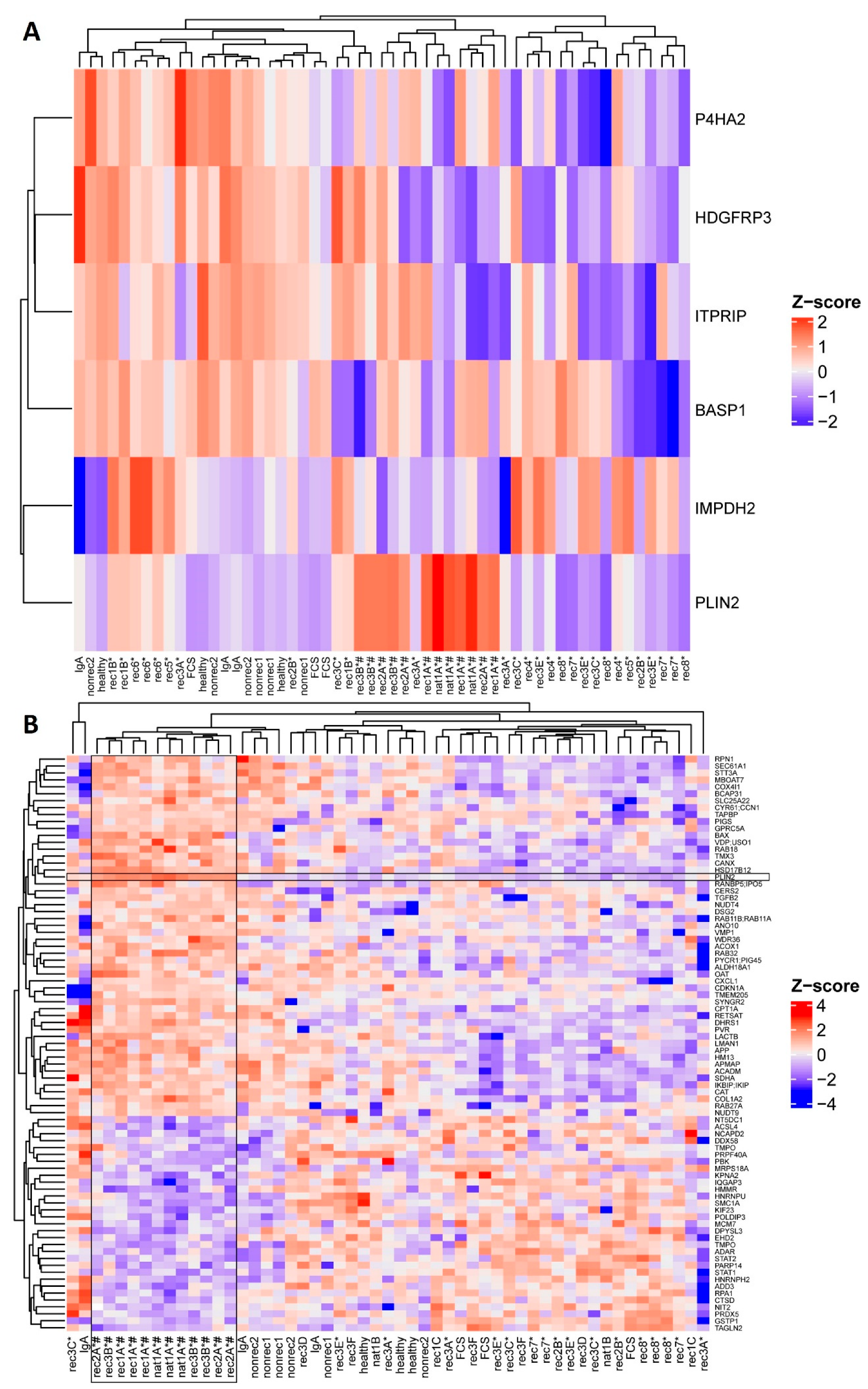
2.1. Active Disease FSGS Plasmas Increase Perilipin-2 Protein Expression in hPod
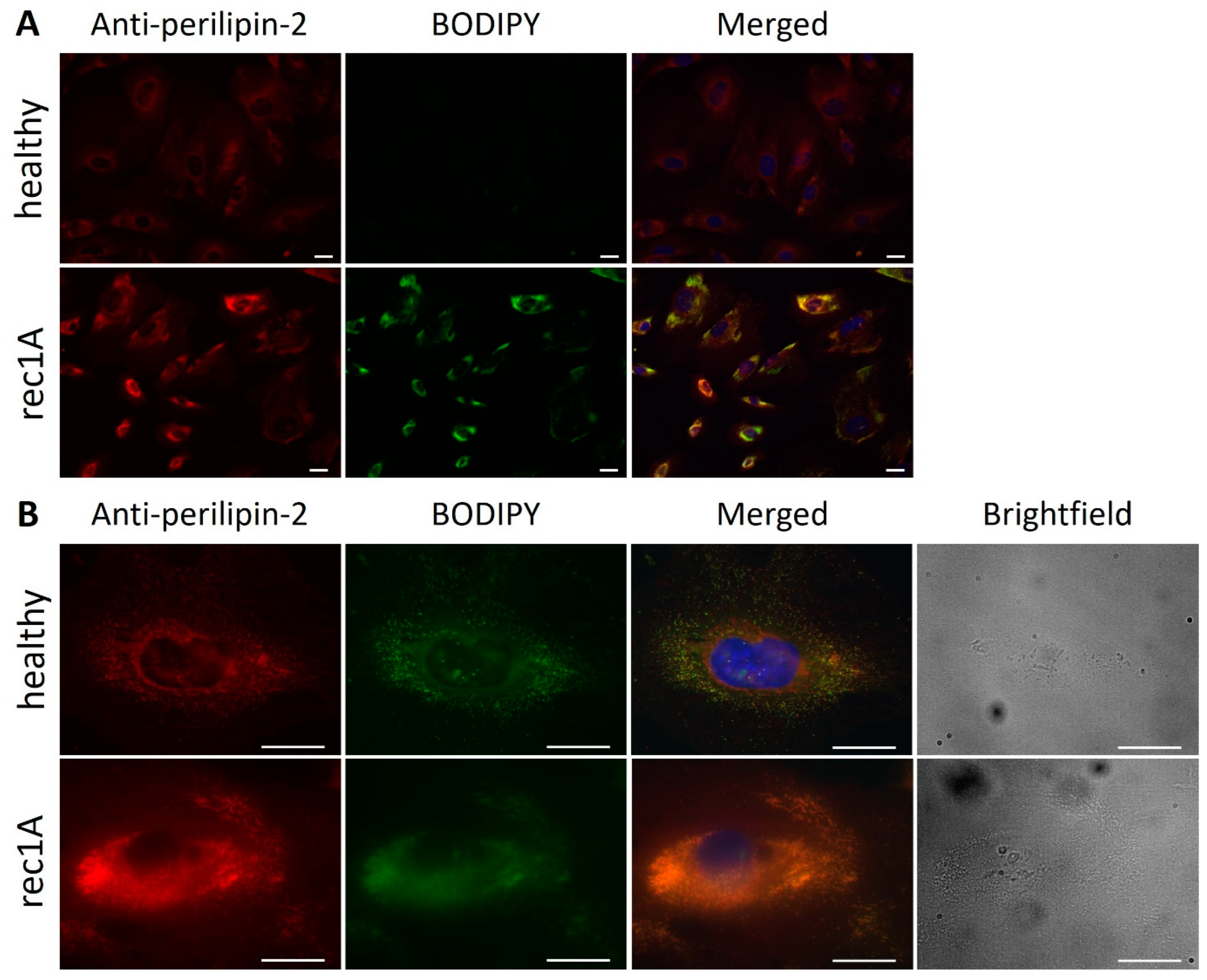
2.2. Increased Perilipin-2 Expression Is Correlated to hPod Granularity and the Presence of Lipid Droplets
2.3. Active Disease FSGS Plasmas Increase the Presence of Perilipin-2 Associated with Lipid Droplets in hPod
2.4. Glomerular Perilipin-2 Expression Is Increased in Patients with FSGS
3. Discussion
4. Materials and Methods
4.1. Patient and Control Material
4.2. Cell Culture
4.3. Sample Preparation for Liquid Chromatography–Mass Spectrometry (LC–MS)
4.4. Mass Spectrometry Analyses
4.5. Data Availability
4.6. RNA Isolation and Real-Time PCR
4.7. Flow Cytometry
4.8. Electron Microscopy Imaging hPod
4.9. Fluorescence Microscopy
4.10. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rudnicki, M. FSGS Recurrence in Adults after Renal Transplantation. BioMed. Res. Int. 2016, 2016, 3295618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maas, R.J.; Deegens, J.K.; van den Brand, J.A.; Cornelissen, E.A.; Wetzels, J.F. A retrospective study of focal segmental glomerulosclerosis: Clinical criteria can identify patients at high risk for recurrent disease after first renal transplantation. BMC Nephrol. 2013, 14, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maas, R.J.; Deegens, J.K.; Wetzels, J.F. Permeability factors in idiopathic nephrotic syndrome: Historical perspectives and lessons for the future. Nephrol. Dial. Transplant. 2014, 29, 2207–2216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Candelier, J.J.; Lorenzo, H.K. Idiopathic nephrotic syndrome and serum permeability factors: A molecular jigsaw puzzle. Cell Tissue Res. 2020, 379, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Shoji, J.; Mii, A.; Terasaki, M.; Shimizu, A. Update on Recurrent Focal Segmental Glomerulosclerosis in Kidney Transplantation. Nephron 2020, 144, 65–70. [Google Scholar] [CrossRef]
- De Vriese, A.S.; Wetzels, J.F.; Glassock, R.J.; Sethi, S.; Fervenza, F.C. Therapeutic trials in adult FSGS: Lessons learned and the road forward. Nat. Rev. Nephrol. 2021, 17, 619–630. [Google Scholar] [CrossRef]
- Deegens, J.K.; Andresdottir, M.B.; Croockewit, S.; Wetzels, J.F. Plasma exchange improves graft survival in patients with recurrent focal glomerulosclerosis after renal transplant. Transpl. Int. 2004, 17, 151–157. [Google Scholar] [CrossRef]
- Fine, R.N. Recurrence of nephrotic syndrome/focal segmental glomerulosclerosis following renal transplantation in children. Pediatr. Nephrol. 2007, 22, 496–502. [Google Scholar] [CrossRef] [Green Version]
- Savin, V.J.; Sharma, M.; Zhou, J.; Genochi, D.; Sharma, R.; Srivastava, T.; Ilahe, A.; Budhiraja, P.; Gupta, A.; McCarthy, E.T. Multiple Targets for Novel Therapy of FSGS Associated with Circulating Permeability Factor. BioMed. Res. Int. 2017, 2017, 6232616. [Google Scholar] [CrossRef] [Green Version]
- den Braanker, D.J.W.; Maas, R.J.; Deegens, J.K.; Yanginlar, C.; Wetzels, J.F.M.; van der Vlag, J.; Nijenhuis, T. Novel in vitro assays to detect circulating permeability factor(s) in idiopathic focal segmental glomerulosclerosis. Nephrol. Dial. Transplant. 2021, 36, 247–256. [Google Scholar] [CrossRef]
- Sharma, R.; Sharma, M.; McCarthy, E.T.; Ge, X.L.; Savin, V.J. Components of normal serum block the focal segmental glomerulosclerosis factor activity in vitro. Kidney Int. 2000, 58, 1973–1979. [Google Scholar] [CrossRef]
- Candiano, G.; Musante, L.; Carraro, M.; Faccini, L.; Campanacci, L.; Zennaro, C.; Artero, M.; Ginevri, F.; Perfumo, F.; Gusmano, R.; et al. Apolipoproteins prevent glomerular albumin permeability induced in vitro by serum from patients with focal segmental glomerulosclerosis. J. Am. Soc. Nephrol. 2001, 12, 143–150. [Google Scholar] [CrossRef]
- Chang, J.W.; Pardo, V.; Sageshima, J.; Chen, L.; Tsai, H.L.; Reiser, J.; Wei, C.; Ciancio, G.; Burke, G.W., 3rd; Fornoni, A. Podocyte foot process effacement in postreperfusion allograft biopsies correlates with early recurrence of proteinuria in focal segmental glomerulosclerosis. Transplantation 2012, 93, 1238–1244. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, T.; Ohsaki, Y.; Suzuki, M.; Cheng, J. Imaging lipid droplets by electron microscopy. Methods Cell Biol. 2013, 116, 227–251. [Google Scholar]
- Cohen, S. Lipid Droplets as Organelles. Int. Rev. Cell Mol. Biol. 2018, 337, 83–110. [Google Scholar]
- Agrawal, S.; Zaritsky, J.J.; Fornoni, A.; Smoyer, W.E. Dyslipidaemia in nephrotic syndrome: Mechanisms and treatment. Nat. Rev. Nephrol. 2018, 14, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Brasaemle, D.L. Thematic review series: Adipocyte biology. The perilipin family of structural lipid droplet proteins: Stabilization of lipid droplets and control of lipolysis. J. Lipid Res. 2007, 48, 2547–2559. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.S.; Kruth, H.S. Accumulation of cholesterol in the lesions of focal segmental glomerulosclerosis. Nephrology 2003, 8, 224–230. [Google Scholar] [CrossRef]
- Ranganathan, S. Pathology of Podocytopathies Causing Nephrotic Syndrome in Children. Front. Pediatr. 2016, 4, 32. [Google Scholar] [CrossRef] [Green Version]
- Jao, W.; Lewy, P.; Norris, S.H.; Pollak, V.E.; Pirani, C.L. Lipoid nephrosis: A reassessment. Perspect. Nephrol. Hypertens. 1973, 1 Pt 1, 183–198. [Google Scholar]
- Mitrofanova, A.; Drexler, Y.; Merscher, S.; Fornoni, A. Role of Sphingolipid Signaling in Glomerular Diseases: Focus on DKD and FSGS. J. Cell. Signal. 2020, 1, 56–69. [Google Scholar] [PubMed]
- Wahl, P.; Ducasa, G.M.; Fornoni, A. Systemic and renal lipids in kidney disease development and progression. Am. J. Physiol. Ren. Physiol. 2016, 310, F433–F445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman-Edelstein, M.; Scherzer, P.; Tobar, A.; Levi, M.; Gafter, U. Altered renal lipid metabolism and renal lipid accumulation in human diabetic nephropathy. J. Lipid Res. 2014, 55, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Brasaemle, D.L.; Barber, T.; Wolins, N.E.; Serrero, G.; Blanchette-Mackie, E.J.; Londos, C. Adipose differentiation-related protein is an ubiquitously expressed lipid storage droplet-associated protein. J. Lipid Res. 1997, 38, 2249–2263. [Google Scholar] [CrossRef] [PubMed]
- Robenek, H.; Hofnagel, O.; Buers, I.; Robenek, M.J.; Troyer, D.; Severs, N.J. Adipophilin-enriched domains in the ER membrane are sites of lipid droplet biogenesis. J. Cell Sci. 2006, 119 Pt 20, 4215–4224. [Google Scholar] [CrossRef] [Green Version]
- Chang, B.H.; Li, L.; Paul, A.; Taniguchi, S.; Nannegari, V.; Heird, W.C.; Chan, L. Protection against fatty liver but normal adipogenesis in mice lacking adipose differentiation-related protein. Mol. Cell. Biol. 2006, 26, 1063–1076. [Google Scholar] [CrossRef] [Green Version]
- Listenberger, L.L.; Ostermeyer-Fay, A.G.; Goldberg, E.B.; Brown, W.J.; Brown, D.A. Adipocyte differentiation-related protein reduces the lipid droplet association of adipose triglyceride lipase and slows triacylglycerol turnover. J. Lipid Res. 2007, 48, 2751–2761. [Google Scholar] [CrossRef] [Green Version]
- Straub, B.K.; Gyoengyoesi, B.; Koenig, M.; Hashani, M.; Pawella, L.M.; Herpel, E.; Mueller, W.; Macher-Goeppinger, S.; Heid, H.; Schirmacher, P. Adipophilin/perilipin-2 as a lipid droplet-specific marker for metabolically active cells and diseases associated with metabolic dysregulation. Histopathology 2013, 62, 617–631. [Google Scholar] [CrossRef]
- Conte, M.; Franceschi, C.; Sandri, M.; Salvioli, S. Perilipin 2 and Age-Related Metabolic Diseases: A New Perspective. Trends Endocrinol. Metab. 2016, 27, 893–903. [Google Scholar] [CrossRef]
- Gao, J.; Serrero, G. Adipose differentiation related protein (ADRP) expressed in transfected COS-7 cells selectively stimulates long chain fatty acid uptake. J. Biol. Chem. 1999, 274, 16825–16830. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Zou, F.; Diao, Z.; Zhang, S.; Deng, Y.; Zhu, X.; Cui, L.; Yu, J.; Zhang, Z.; Bamigbade, A.T.; et al. Perilipin 2 and lipid droplets provide reciprocal stabilization. Biophys. Rep. 2019, 5, 145–160. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yang, Q.; Yang, J.; Ma, Y.; Ding, G. Angiotensin II induces cholesterol accumulation and injury in podocytes. Sci. Rep. 2017, 7, 10672. [Google Scholar] [CrossRef] [Green Version]
- Dai, Z.W.; Cai, K.D.; Xu, L.C.; Wang, L.L. Perilipin2 inhibits diabetic nephropathy-induced podocyte apoptosis by activating the PPARgamma signaling pathway. Mol. Cell. Probes 2020, 53, 101584. [Google Scholar] [CrossRef]
- Urahama, Y.; Ohsaki, Y.; Fujita, Y.; Maruyama, S.; Yuzawa, Y.; Matsuo, S.; Fujimoto, T. Lipid droplet-associated proteins protect renal tubular cells from fatty acid-induced apoptosis. Am. J. Pathol. 2008, 173, 1286–1294. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Luo, Y.; Yang, S.; Zeng, M.; Zhang, S.; Liu, J.; Han, Y.; Liu, Y.; Zhu, X.; Wu, H.; et al. Ectopic lipid accumulation: Potential role in tubular injury and inflammation in diabetic kidney disease. Clin. Sci. 2018, 132, 2407–2422. [Google Scholar] [CrossRef]
- Conte, M.; Santoro, A.; Collura, S.; Martucci, M.; Battista, G.; Bazzocchi, A.; Morsiani, C.; Sevini, F.; Capri, M.; Monti, D.; et al. Circulating perilipin 2 levels are associated with fat mass, inflammatory and metabolic markers and are higher in women than men. Aging 2021, 13, 7931–7942. [Google Scholar] [CrossRef]
- Orlicky, D.J.; Libby, A.E.; Bales, E.S.; McMahan, R.H.; Monks, J.; La Rosa, F.G.; McManaman, J.L. Perilipin-2 promotes obesity and progressive fatty liver disease in mice through mechanistically distinct hepatocyte and extra-hepatocyte actions. J. Physiol. 2019, 597, 1565–1584. [Google Scholar] [CrossRef] [Green Version]
- McManaman, J.L.; Bales, E.S.; Orlicky, D.J.; Jackman, M.; MacLean, P.S.; Cain, S.; Crunk, A.E.; Mansur, A.; Graham, C.E.; Bowman, T.A.; et al. Perilipin-2-null mice are protected against diet-induced obesity, adipose inflammation, and fatty liver disease. J. Lipid Res. 2013, 54, 1346–1359. [Google Scholar] [CrossRef] [Green Version]
- Prasad, R.; Hadjidemetriou, I.; Maharaj, A.; Meimaridou, E.; Buonocore, F.; Saleem, M.; Hurcombe, J.; Bierzynska, A.; Barbagelata, E.; Bergada, I.; et al. Sphingosine-1-phosphate lyase mutations cause primary adrenal insufficiency and steroid-resistant nephrotic syndrome. J. Clin. Investig. 2017, 127, 942–953. [Google Scholar] [CrossRef] [Green Version]
- Morris, A.P.; Le, T.H.; Wu, H.; Akbarov, A.; van der Most, P.J.; Hemani, G.; Smith, G.D.; Mahajan, A.; Gaulton, K.J.; Nadkarni, G.N.; et al. Trans-ethnic kidney function association study reveals putative causal genes and effects on kidney-specific disease aetiologies. Nat. Commun. 2019, 10, 29. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Kidd, J.; Kaspar, C.; Dempsey, S.; Bhat, O.M.; Camus, S.; Ritter, J.K.; Gehr, T.W.B.; Gulbins, E.; Li, P.L. Podocytopathy and Nephrotic Syndrome in Mice with Podocyte-Specific Deletion of the Asah1 Gene: Role of Ceramide Accumulation in Glomeruli. Am. J. Pathol. 2020, 190, 1211–1223. [Google Scholar] [CrossRef] [PubMed]
- Fornoni, A.; Sageshima, J.; Wei, C.; Merscher-Gomez, S.; Aguillon-Prada, R.; Jauregui, A.N.; Li, J.; Mattiazzi, A.; Ciancio, G.; Chen, L.; et al. Rituximab targets podocytes in recurrent focal segmental glomerulosclerosis. Sci. Transl. Med. 2011, 3, 85ra46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kronbichler, A.; Gauckler, P.; Bruchfeld, A. Rituximab in minimal change disease and focal segmental glomerulosclerosis. Nephrol. Dial. Transplant. 2021, 36, 983–985. [Google Scholar] [CrossRef] [PubMed]
- Mitrofanova, A.; Mallela, S.K.; Ducasa, G.M.; Yoo, T.H.; Rosenfeld-Gur, E.; Zelnik, I.D.; Molina, J.; Varona Santos, J.; Ge, M.; Sloan, A.; et al. SMPDL3b modulates insulin receptor signaling in diabetic kidney disease. Nat. Commun. 2019, 10, 2692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallela, S.K.; Mitrofanova, A.; Merscher, S.; Fornoni, A. Regulation of the amount of ceramide-1-phosphate synthesized in differentiated human podocytes. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2019, 1864, 158517. [Google Scholar] [CrossRef]
- Muller-Deile, J.; Sarau, G.; Kotb, A.M.; Jaremenko, C.; Rolle-Kampczyk, U.E.; Daniel, C.; Kalkhof, S.; Christiansen, S.H.; Schiffer, M. Novel diagnostic and therapeutic techniques reveal changed metabolic profiles in recurrent focal segmental glomerulosclerosis. Sci. Rep. 2021, 11, 4577. [Google Scholar] [CrossRef]
- Saleem, M.A.; O’Hare, M.J.; Reiser, J.; Coward, R.J.; Inward, C.D.; Farren, T.; Xing, C.Y.; Ni, L.; Mathieson, P.W.; Mundel, P. A conditionally immortalized human podocyte cell line demonstrating nephrin and podocin expression. J. Am. Soc. Nephrol. 2002, 13, 630–638. [Google Scholar] [CrossRef]
- Hohne, M.; Frese, C.K.; Grahammer, F.; Dafinger, C.; Ciarimboli, G.; Butt, L.; Binz, J.; Hackl, M.J.; Rahmatollahi, M.; Kann, M.; et al. Single nephron proteomes connect morphology and function in proteinuric kidney disease. Kidney Int. 2018, 93, 1308–1319. [Google Scholar] [CrossRef]
- Rappsilber, J.; Mann, M.; Ishihama, Y. Protocol for micro-purification, enrichment, pre-fractionation and storage of peptides for proteomics using StageTips. Nat. Protoc. 2007, 2, 1896–1906. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate proteome-wide label-free quantification by delayed normalization and maximal peptide ratio extraction, termed MaxLFQ. Mol. Cell. Proteom. 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [Green Version]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Mi, H.; Ebert, D.; Muruganujan, A.; Mills, C.; Albou, L.P.; Mushayamaha, T.; Thomas, P.D. PANTHER version 16: A revised family classification, tree-based classification tool, enhancer regions and extensive API. Nucleic Acids Res. 2021, 49, D394–D403. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient Code | Gender | Sample Code | Age at Sampling Time | PP Session Number | PP-Responsive Yes/No | Kidney Transplant Number | Onset of Recurrence after TX |
| rec1 | M | rec1A *,# | 73 | 1 | yes | pre-TX | NA |
| rec1B * | 74 | 1 | yes | 1 | Day 3 | ||
| rec1C | 74 | 4 | yes | 1 | |||
| rec2 | M | rec2A *,# | 61 | 1 | no | pre-TX | Day 2 |
| rec2B * | 61 | 5 | no | pre-TX | |||
| rec3 | M | rec3A * | 21 | 1 | no | pre-TX | NA |
| rec3B *,# | 21 | 10 | no | pre-TX | NA | ||
| rec3C * | 24 | 1 | yes | 1 | Day 3 | ||
| rec3D | 24 | 9 | yes | 1 | |||
| rec3E * | 24 | 1 | yes | 1 | Day 29 | ||
| rec3F | 24 | 4 | yes | 1 | |||
| rec4 | M | rec4 * | 20 | 1 | no | 1 | Day 2 |
| rec5 | M | rec5 * | 26 | 1 | no | 1 | Day 2 |
| rec6 | F | rec6 * | 40 | 1 | yes | 1 | 2 months |
| rec7 | F | rec7 * | 32 | 1 | yes | 3 | Day 1 |
| rec8 | F | rec8 * | 39 | 1 | yes | 1 | Day 1 |
| nat1 | M | nat1A *,# | 18 | 1 | yes | NA | NA |
| nat1B | 21 | Maintenance PP | yes | NA | NA | ||
| healthy | 3F, 2unk | healthy | unk | no PP | NA | NA | NA |
| IgA | F | IgA | 66 | 1 | NA | NA | NA |
| nonrec1 | M | nonrec1 | 68 | no PP | NA | 1 | NA |
| nonrec2 | M | nonrec2 | 27 | no PP | NA | 1 | NA |
| Patient Code | Gender | Sample Code | Serum creatinine (µmol/L) | Serum albumin (g/L) | Proteinuria (g/10 mmol creatinine) | Immunosuppression at Sampling Time | |
| rec1 | M | rec1A *,# | 742 | 22 | 7.5 | None (pre-TX sample) | |
| rec1B * | 169 | 28 | 4.0 | Tacro, MMF, Pred | |||
| rec1C | 168 | 31 | 1.5 | Tacro, MMF, Pred | |||
| rec2 | M | rec2A *,# | 200 | 18 | 11.8 | Pred | |
| rec2B * | 203 | 24 | 11.4 | Pred | |||
| rec3 | M | rec3A * | 165 | 10 | 11.8 | Pred | |
| rec3B *,# | 127 | 12 | 10.5 | Pred | |||
| rec3C * | 200 | 18 | 5.4 | Tacro, MMF, Pred | |||
| rec3D | 185 | 26 | 1.8 g/day | Tacro, MMF, Pred | |||
| rec3E * | 193 | 29 | 4.5 | Tacro, MMF, Pred, Rtx | |||
| rec3F | 193 | 32 | 1.4 | Tacro, MMF, Pred, Rtx | |||
| rec4 | M | rec4 * | 127 | 24 | 23.0 | Tacro, MMF, Pred | |
| rec5 | M | rec5 * | 271 | 29 | 9.5 | Bas pre-TX, Tacro, MMF, Pred | |
| rec6 | F | rec6 * | 184 | 40 | 11.6 | Tacro, MMF, Pred | |
| rec7 | F | rec7 * | 678 | 40 | 9.0 | Rtx pre-TX, Tacro, MMF, Pred | |
| rec8 | F | rec8 * | 146 | 27 | 20.0 | Tacro, MMF, Pred | |
| nat1 | M | nat1A *,# | 459 | 9 | 7.3 | Mpred | |
| nat1B | 74 | 34.4 | 2.3 | Mpred | |||
| healthy | 3F, 2unk | healthy | unk | unk | unk | None | |
| IgA | F | IgA | 574 | 26 | Oliguric | None | |
| nonrec1 | M | nonrec1 | 86 | 39 | <0.1 | Tacro, Pred | |
| nonrec2 | M | nonrec2 | 224 | 34 | 0.4 | Tacro, MMF, Pred | |
| Patient Code | Gender | Age at Sampling Time | PP-Responsive Yes/No | Kidney Transplant Number | Onset of Recurrence after TX |
| rec1 | M | 68 | yes | pre-TX | Day 3 |
| nat1 | M | 18 | yes | NA | NA |
| IgA | F | 66 | NA | NA | NA |
| MN1 | F | 59 | NA | NA | NA |
| MN2 | M | 64 | NA | NA | NA |
| Patient Code | Gender | Serum creatinine (µmol/L) | Serum albumin (g/L) | Proteinuria (g/10 mmol creatinine) | Immunosuppression at Sampling Time |
| rec1 | M | 182 | 31 | 10.0 | None (pre-TX sample) |
| nat1 | M | 418 | 7 | 12.9 | Mpred |
| IgA | F | 468 | 24 | 3.5 | None |
| MN1 | F | 70 | 27 | 9.0 | None |
| MN2 | M | 130 | 21 | 3.7 | None |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
den Braanker, D.J.W.; Maas, R.J.H.; van Mierlo, G.; Parr, N.M.J.; Bakker-van Bebber, M.; Deegens, J.K.J.; Jansen, P.W.T.C.; Gloerich, J.; Willemsen, B.; Dijkman, H.B.; et al. Primary Focal Segmental Glomerulosclerosis Plasmas Increase Lipid Droplet Formation and Perilipin-2 Expression in Human Podocytes. Int. J. Mol. Sci. 2023, 24, 194. https://doi.org/10.3390/ijms24010194
den Braanker DJW, Maas RJH, van Mierlo G, Parr NMJ, Bakker-van Bebber M, Deegens JKJ, Jansen PWTC, Gloerich J, Willemsen B, Dijkman HB, et al. Primary Focal Segmental Glomerulosclerosis Plasmas Increase Lipid Droplet Formation and Perilipin-2 Expression in Human Podocytes. International Journal of Molecular Sciences. 2023; 24(1):194. https://doi.org/10.3390/ijms24010194
Chicago/Turabian Styleden Braanker, Dirk J. W., Rutger J. H. Maas, Guido van Mierlo, Naomi M. J. Parr, Marinka Bakker-van Bebber, Jeroen K. J. Deegens, Pascal W. T. C. Jansen, Jolein Gloerich, Brigith Willemsen, Henry B. Dijkman, and et al. 2023. "Primary Focal Segmental Glomerulosclerosis Plasmas Increase Lipid Droplet Formation and Perilipin-2 Expression in Human Podocytes" International Journal of Molecular Sciences 24, no. 1: 194. https://doi.org/10.3390/ijms24010194