
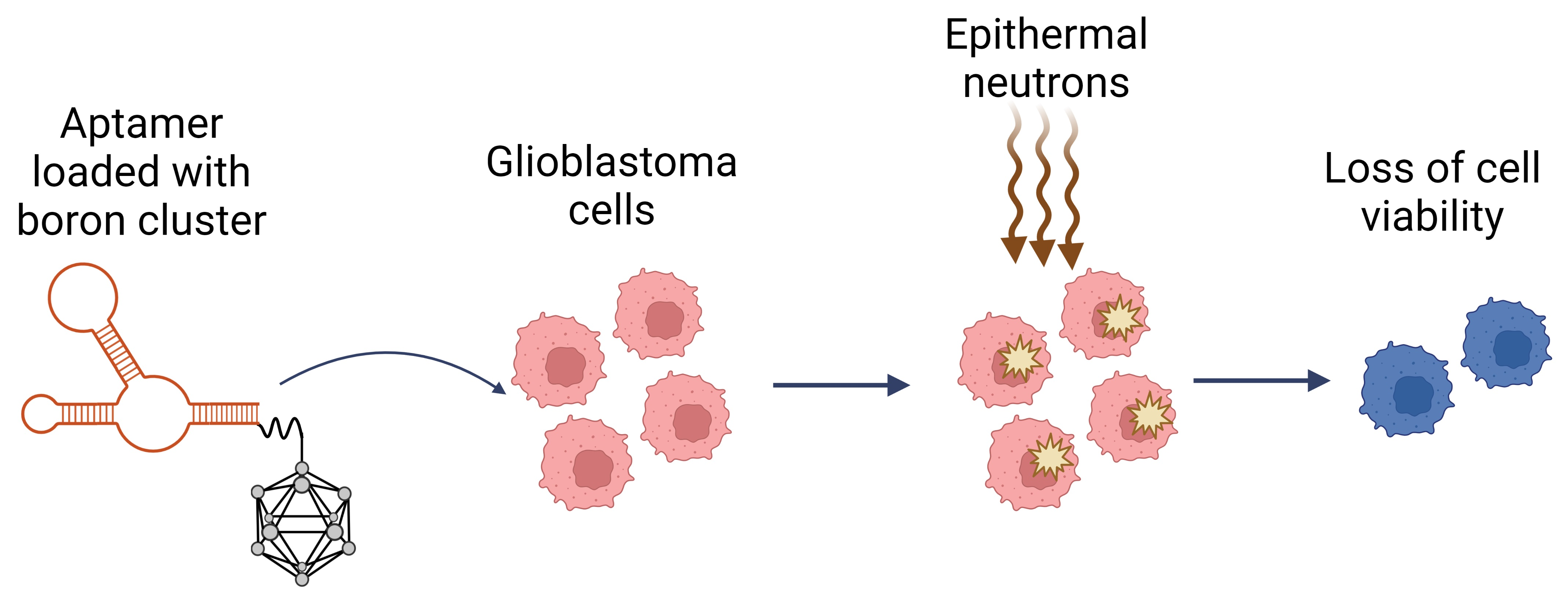
Aptamers for Addressed Boron Delivery in BNCT: Effect of Boron Cluster Attachment Site on Functional Activity

 , , , ,
, , , ,  and
and
Abstract
:
1. Introduction
2. Results
2.1. 2′-F-RNA Aptamers Capable of Internalizing into U-87 MG Human Glioblastoma Cells
2.2. Synthesis of Conjugates of 2′-F-RNA Aptamers with Closo-Dodecaborate
2.3. Evaluation of Cell Penetrating Efficiency for Boron-Containing Aptamer Conjugates
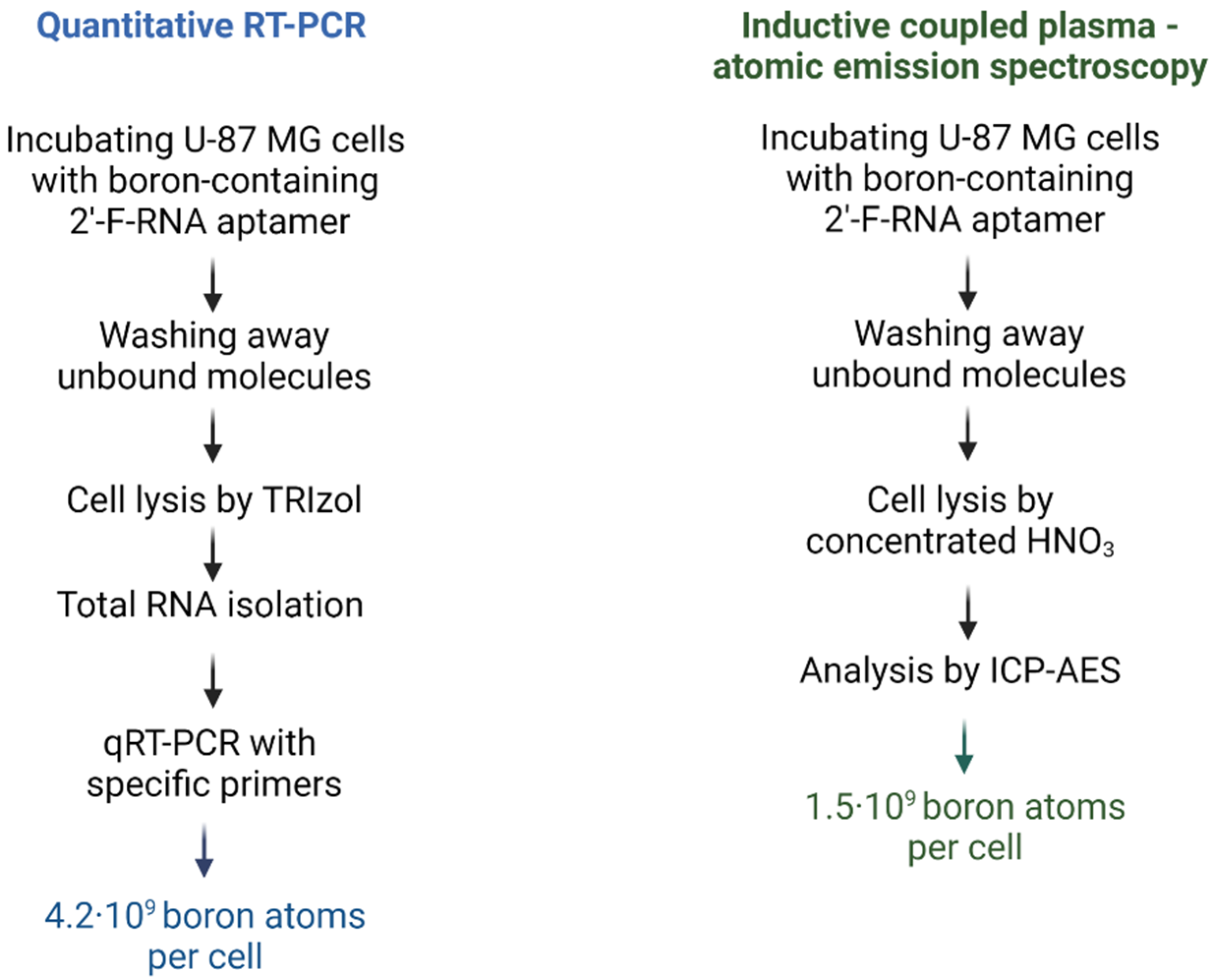
2.3.1. Evaluation of Intracellular Level of 2′-F-RNA Aptamer–Boron Cluster Conjugate by qRT-PCR
2.3.2. Evaluation of Intracellular Level of 2′-F-RNA Aptamer–Boron Cluster Conjugate by Direct Boron Measurement
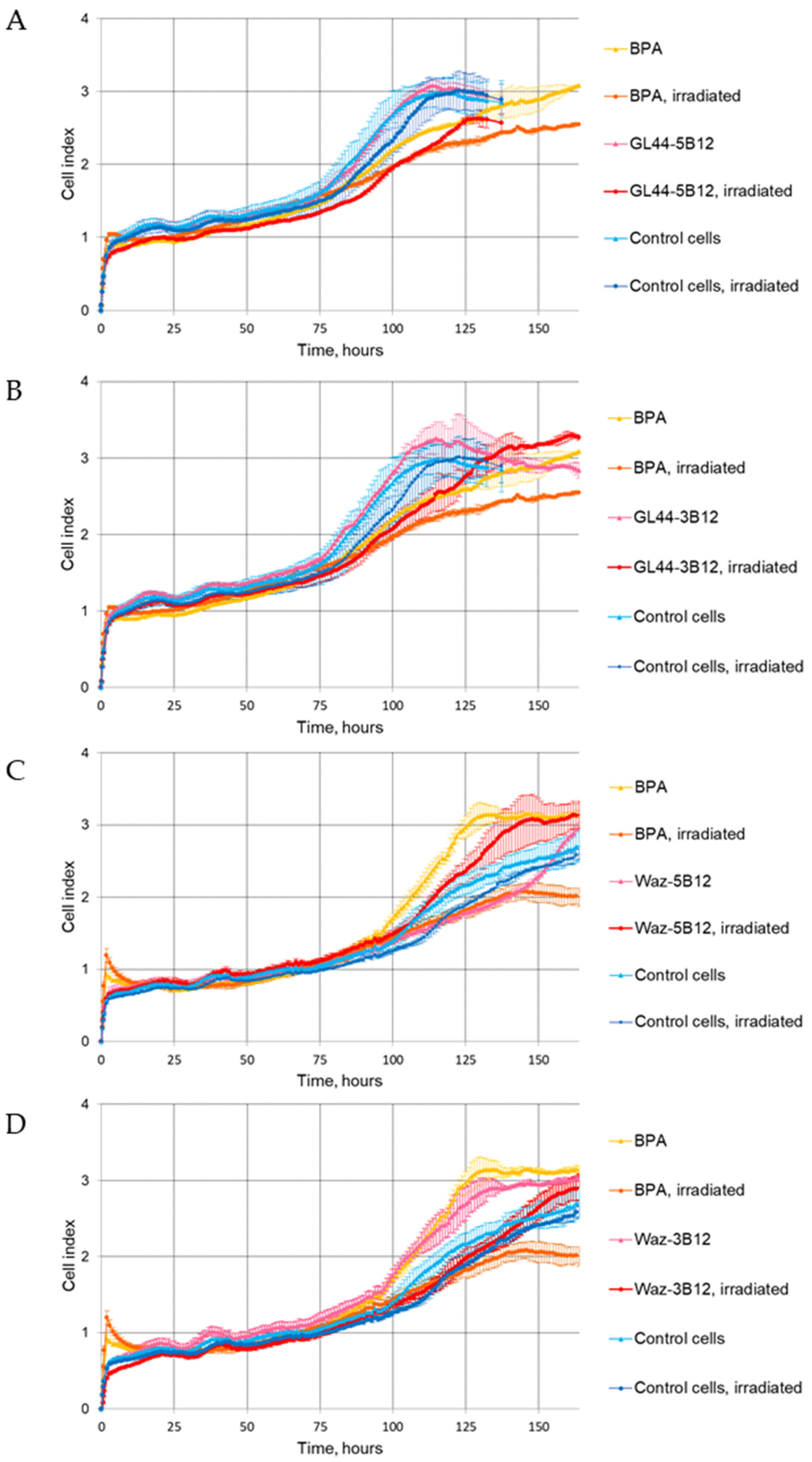
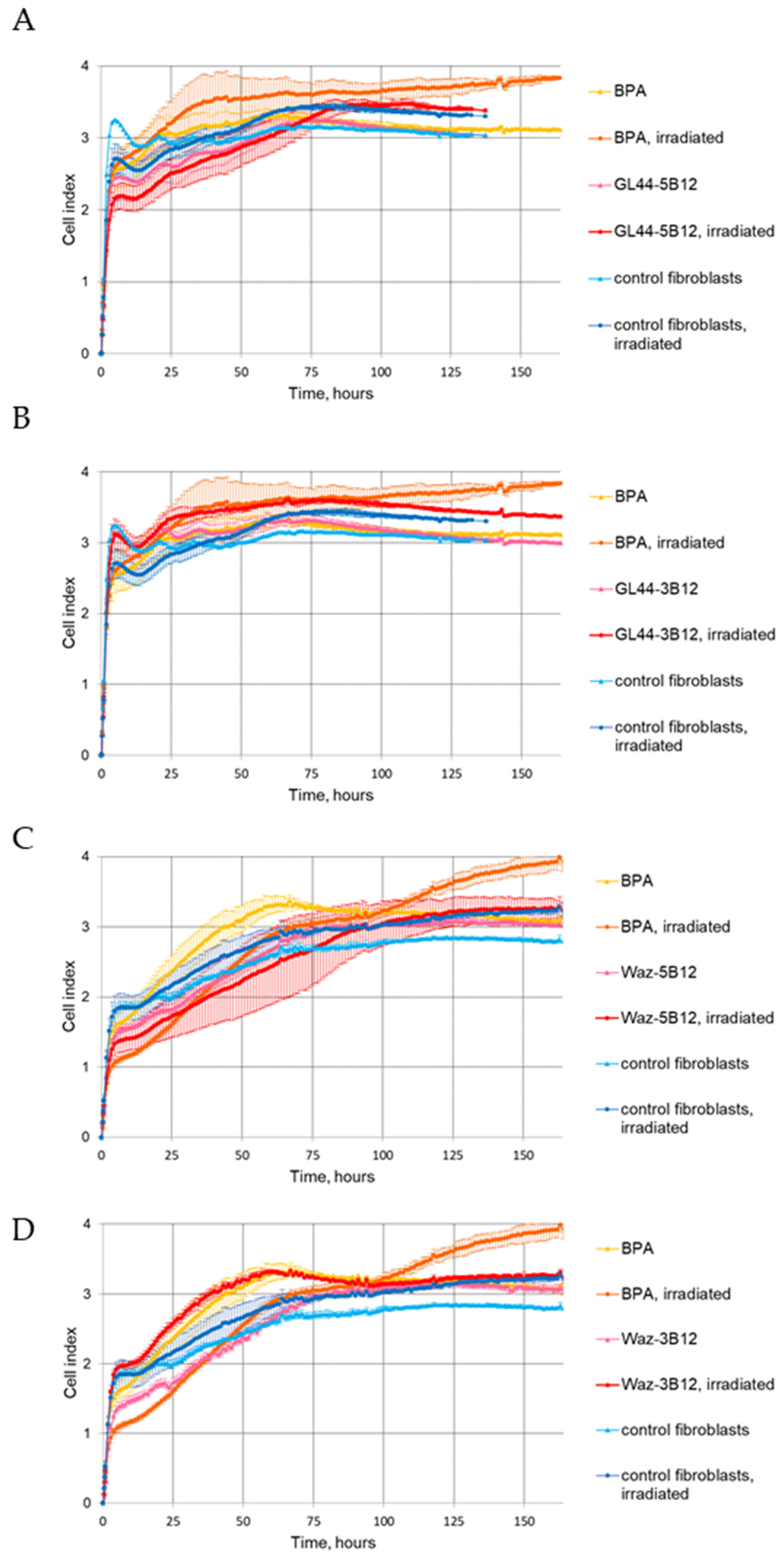
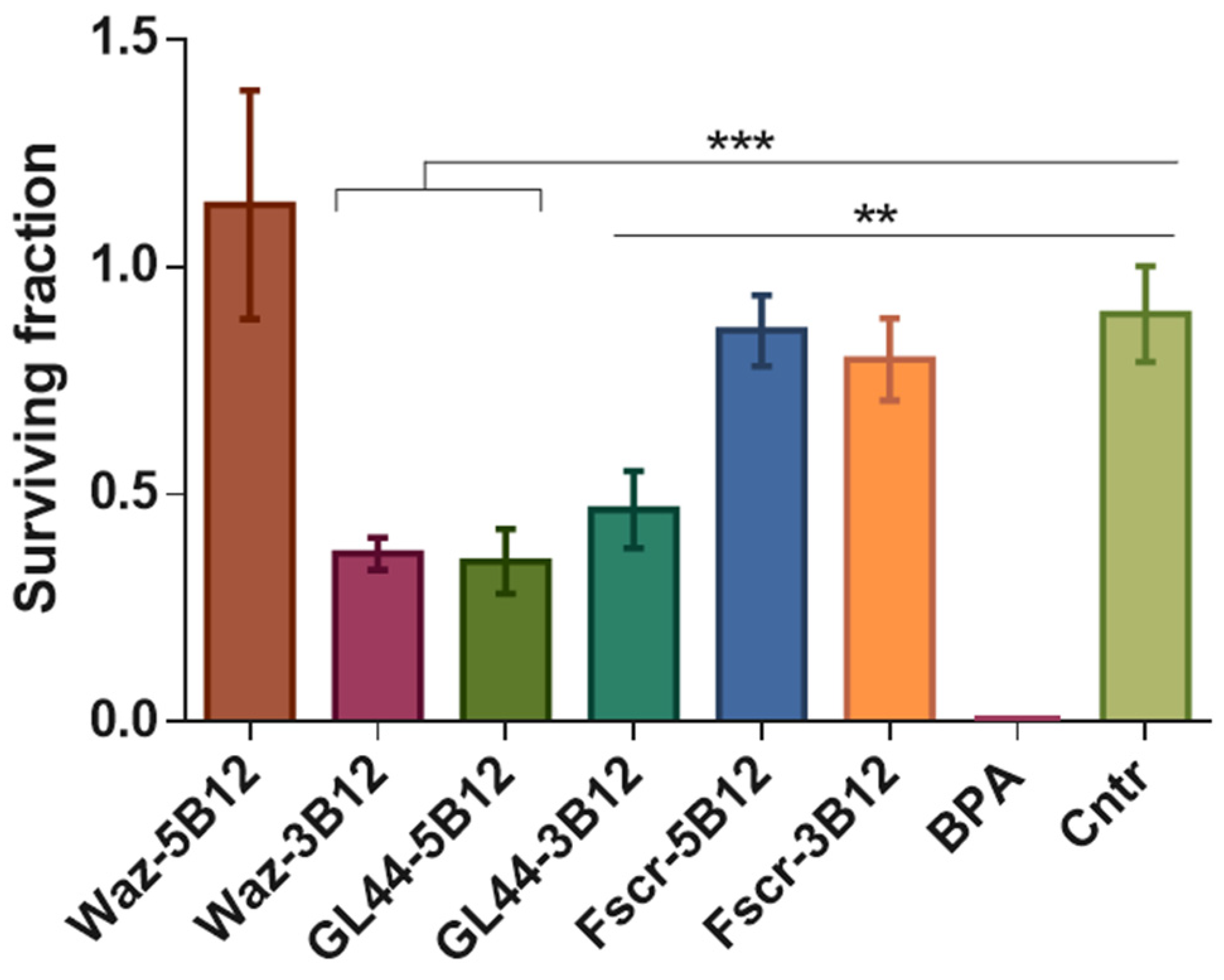
2.4. The Effect of Aptamer–Boron Cluster Conjugate on Cell Viability after Irradiation
2.4.1. The xCELLigence Real-Time Cell Analysis (RTCA)
2.4.2. Clonogenic Assay
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Lines
4.3. Synthesis of 2′-F-RNA Aptamers
4.4. Synthesis of Sulfo-Cy5-Labeled 2′-F-RNA Aptamers
4.5. Synthesis of 2′-F-RNA Conjugates with Closo-Dodecaborate
4.6. Confocal Microscopy
4.7. Evaluation of Cell Internalization of 2′-F-RNA Aptamer
4.7.1. Quantitative RT-PCR Assay
4.7.2. Inductive Coupled Plasma–Atomic Emission Spectrometry (ICP-AES) Assay
4.8. Model BNCT Experiments
4.9. The xCELLigence Real-Time Cell Analysis (RTCA)
4.10. Clonogenic Assay
4.11. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AMA | a mixture of ammonium hydroxide and 40% aqueous methylamine |
| BINP | Budker Institute of Nuclear Physics |
| BNCT | Boron Neutron Capture Therapy |
| BPA | Boronophenylalanine |
| BSH | Sodium borocaptate |
| CI | Cell Index |
| DAPI | 4′,6-Diamidino-2-phenylindole |
| DIPEA | N,N-Diisopropylethylamine |
| DMSO | Dimethyl sulfoxide |
| DSC | N,N′-Disuccinimidyl carbonate |
| ESI | Electrospray Ionization |
| IMDM | Iscove’s Modified Dulbecco’s Medium |
| ICP-AES | Inductively Coupled Plasma–Atomic Emission Spectrometry |
| αMEM | Modified Minimal Essential Medium |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide |
| NGF | Nerve Growth Factor |
| TBDMS | Tert-butyldimethylsilyl |
| TEA | Triethylamine |
| THF | Tetrahydrofuran |
| NHS | N-Hydroxysuccinimide |
| NMP | N-Methyl pyrrolidone |
| PBS | Phosphate Buffered Saline |
| RTCA | Real-Time Cell Analysis |
| TBTA | Tris(benzyltriazolylmethyl)amine |
References
- Skwierawska, D.; López-Valverde, J.A.; Balcerzyk, M.; Leal, A. Clinical Viability of Boron Neutron Capture Therapy for Personalized Radiation Treatment. Cancers 2022, 14, 2865. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Li, F.; Liang, L. Boron Neutron Capture Therapy: Clinical Application and Research Progress. Curr. Oncol. 2022, 29, 7868–7886. [Google Scholar] [CrossRef] [PubMed]
- Malouff, T.D.; Seneviratne, D.S.; Ebner, D.K.; Stross, W.C.; Waddle, M.R.; Trifiletti, D.M.; Krishnan, S. Boron Neutron Capture Therapy: A Review of Clinical Applications. Front. Oncol. 2021, 11, 601820. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Li, J.; Jiang, P.; Tian, S.; Wang, H.; Fan, R.; Liu, J.; Yang, Y.; Liu, Z.; Wang, J. The Basis and Advances in Clinical Application of Boron Neutron Capture Therapy. Radiat. Oncol. 2021, 16, 216. [Google Scholar] [CrossRef] [PubMed]
- Taskaev, S.; Berendeev, E.; Bikchurina, M.; Bykov, T.; Kasatov, D.; Kolesnikov, I.; Koshkarev, A.; Makarov, A.; Ostreinov, G.; Porosev, V.; et al. Neutron Source Based on Vacuum Insulated Tandem Accelerator and Lithium Target. Biology 2021, 10, 350. [Google Scholar] [CrossRef]
- Dymova, M.A.; Taskaev, S.Y.; Richter, V.A.; Kuligina, E.V. Boron Neutron Capture Therapy: Current Status and Future Perspectives. Cancer Commun 2020, 40, 406–421. [Google Scholar] [CrossRef]
- Hu, K.; Yang, Z.; Zhang, L.; Xie, L.; Wang, L.; Xu, H.; Josephson, L.; Liang, S.H.; Zhang, M.R. Boron Agents for Neutron Capture Therapy. Coord. Chem. Rev. 2020, 405, 213139. [Google Scholar] [CrossRef]
- Kanno, H.; Nagata, H.; Ishiguro, A.; Tsuzuranuki, S.; Nakano, S.; Nonaka, T.; Kiyohara, K.; Kimura, T.; Sugawara, A.; Okazaki, Y.; et al. Designation Products: Boron Neutron Capture Therapy for Head and Neck Carcinoma. Oncologist 2021, 26, e1250–e1255. [Google Scholar] [CrossRef]
- Barth, R.F.; Mi, P.; Yang, W. Boron Delivery Agents for Neutron Capture Therapy of Cancer. Cancer Commun. 2018, 38, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Shigdar, S.; Schrand, B.; Giangrande, P.H.; de Franciscis, V. Aptamers: Cutting Edge of Cancer Therapies. Mol. Ther. 2021, 29, 2396–2411. [Google Scholar] [CrossRef]
- Vorobyeva, M.A.; Dymova, M.A.; Novopashina, D.S.; Kuligina, E.V.; Timoshenko, V.V.; Kolesnikov, I.A.; Taskaev, S.Y.; Richter, V.A.; Venyaminova, A.G. Tumor Cell-Specific 2′-Fluoro RNA Aptamer Conjugated with Closo-Dodecaborate as A Potential Agent for Boron Neutron Capture Therapy. Int. J. Mol. Sci. 2021, 22, 7326. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.; Amini, S.; White, M.K. General Overview of Neuronal Cell Culture. In Neuronal Cell Culture. Methods in Molecular Biology; Amini, S., White, M., Eds.; Humana Press: Totowa, NJ, USA, 2013; Volume 1078, pp. 1–8. [Google Scholar]
- Zerrouqi, A.; Pyrzynska, B.; Febbraio, M.; Brat, D.J.; van Meir, E.G. P14ARF Inhibits Human Glioblastoma–Induced Angiogenesis by Upregulating the Expression of TIMP3. J. Clin. Investig. 2012, 122, 1283–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budden, T.; Gaudy-Marqueste, C.; Porter, A.; Kay, E.; Gurung, S.; Earnshaw, C.H.; Roeck, K.; Craig, S.; Traves, V.; Krutmann, J.; et al. Ultraviolet Light-Induced Collagen Degradation Inhibits Melanoma Invasion. Nat. Commun. 2021, 12, 2742. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Cao, X.; Liu, Q.; Zhu, Q.; Liu, K.; Deng, T.; Yu, Q.; Deng, W.; Yu, J.; Wang, Q.; et al. Hybrid Membrane-Derived Nanoparticles for Isoliquiritin Enhanced Glioma Therapy. Pharmaceuticals 2022, 15, 1059. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Tong, Q.; Chen, X.; Jiang, P.; Yu, H.; Zhao, Q.; Sun, L.; Liu, C.; Gu, B.; Zheng, Y.; et al. PHC1 Maintains Pluripotency by Organizing Genome-Wide Chromatin Interactions of the Nanog Locus. Nat. Commun. 2021, 12, 2829. [Google Scholar] [CrossRef]
- Griffante, G.; Gugliesi, F.; Pasquero, S.; Dell’Oste, V.; Biolatti, M.; Salinger, A.J.; Mondal, S.; Thompson, P.R.; Weerapana, E.; Lebbink, R.J.; et al. Human Cytomegalovirus-Induced Host Protein Citrullination Is Crucial for Viral Replication. Nat. Commun. 2021, 12, 3910. [Google Scholar] [CrossRef]
- Sierra-Fonseca, J.A.; Najera, O.; Martinez-Jurado, J.; Walker, E.M.; Varela-Ramirez, A.; Khan, A.M.; Miranda, M.; Lamango, N.S.; Roychowdhury, S. Nerve Growth Factor Induces Neurite Outgrowth of PC12 Cells by Promoting Gβγ-Microtubule Interaction. BMC Neurosci. 2014, 15, 132. [Google Scholar] [CrossRef] [Green Version]
- Hu, R.; Cao, Q.; Sun, Z.; Chen, J.; Zheng, Q.; Xiao, F. A Novel Method of Neural Differentiation of PC12 Cells by Using Opti-MEM as a Basic Induction Medium. Int. J. Mol. Med. 2017, 41, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Maier, K.E.; Jangra, R.K.; Shieh, K.R.; Cureton, D.K.; Xiao, H.; Snapp, E.L.; Whelan, S.P.; Chandran, K.; Levy, M. A New Transferrin Receptor Aptamer Inhibits New World Hemorrhagic Fever Mammarenavirus Entry. Mol. Ther. Nucleic Acids 2016, 5, e321. [Google Scholar] [CrossRef] [Green Version]
- Porciani, D.; Cardwell, L.N.; Tawiah, K.D.; Alam, K.K.; Lange, M.J.; Daniels, M.A.; Burke, D.H. Modular Cell-Internalizing Aptamer Nanostructure Enables Targeted Delivery of Large Functional RNAs in Cancer Cell Lines. Nat. Commun. 2018, 9, 2283. [Google Scholar] [CrossRef]
- Ramalho, M.J.; Loureiro, J.A.; Coelho, M.A.N.; Pereira, M.C. Transferrin Receptor-Targeted Nanocarriers: Overcoming Barriers to Treat Glioblastoma. Pharmaceutics 2022, 14, 279. [Google Scholar] [CrossRef] [PubMed]
- Novopashina, D.S.; Vorobyeva, M.A.; Lomzov, A.A.; Silnikov, V.N.; Venyaminova, A.G. Terminal Mono- and Bis-Conjugates of Oligonucleotides with Closo-Dodecaborate: Synthesis and Physico-Chemical Properties. Int. J. Mol. Sci. 2020, 22, 182. [Google Scholar] [CrossRef] [PubMed]
- Thiel, K.W.; Hernandez, L.I.; Dassie, J.P.; Thiel, W.H.; Liu, X.; Stockdale, K.R.; Rothman, A.M.; Hernandez, F.J.; McNamara, J.O.; Giangrande, P.H. Delivery of Chemo-Sensitizing SiRNAs to HER2+-Breast Cancer Cells Using RNA Aptamers. Nucleic Acids Res 2012, 40, 6319–6337. [Google Scholar] [CrossRef] [Green Version]
- Paterson, J.; Webster, C.I. Exploiting Transferrin Receptor for Delivering Drugs across the Blood-Brain Barrier. Drug Discov. Today Technol. 2016, 20, 49–52. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zheng, M.; Shimoni, O.; Banks, W.A.; Bush, A.I.; Gamble, J.R.; Shi, B. Development of Novel Therapeutics Targeting the Blood–Brain Barrier: From Barrier to Carrier. Adv. Sci. 2021, 8, e2101090. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Johnsen, K.B.; Kucharz, K.; Lauritzen, M.; Moos, T. Blood–Brain Barrier Transport of Transferrin Receptor-Targeted Nanoparticles. Pharmaceutics 2022, 14, 2237. [Google Scholar] [CrossRef] [PubMed]
- Kho, D.; MacDonald, C.; Johnson, R.; Unsworth, C.; O’Carroll, S.; Mez, E.; Angel, C.; Graham, E. Application of XCELLigence RTCA Biosensor Technology for Revealing the Profile and Window of Drug Responsiveness in Real Time. Biosensors 2015, 5, 199–222. [Google Scholar] [CrossRef] [Green Version]
- Coderre, J.A.; Turcotte, J.C.; Riley, K.J.; Binns, P.J.; Harling, O.K.; Kiger, W.S. Boron Neutron Capture Therapy: Cellular Targeting of High Linear Energy Transfer Radiation. Technol. Cancer Res. Treat. 2003, 2, 355–375. [Google Scholar] [CrossRef]
- Meschaninova, M.I.; Novopashina, D.S.; Semikolenova, O.A.; Silnikov, V.N.; Venyaminova, A.G. Novel Convenient Approach to the Solid-Phase Synthesis of Oligonucleotide Conjugates. Molecules 2019, 24, 4266. [Google Scholar] [CrossRef] [Green Version]
- Aptekar, S.; Arora, M.; Lawrence, C.L.; Lea, R.W.; Ashton, K.; Dawson, T.; Alder, J.E.; Shaw, L. Selective Targeting to Glioma with Nucleic Acid Aptamers. PLoS ONE 2015, 10, e0134957. [Google Scholar] [CrossRef]
- Davydova, A.; Krasitskaya, V.; Vorobjev, P.; Timoshenko, V.; Tupikin, A.; Kabilov, M.; Frank, L.; Venyaminova, A.; Vorobyeva, M. Reporter-Recruiting Bifunctional Aptasensor for Bioluminescent Analytical Assays. RSC Adv. 2020, 10, 32393–32399. [Google Scholar] [CrossRef] [PubMed]
- Şener, L.T.; Albeniz, G.; Dinç, B.; Albeniz, I. ICELLigence Real-Time Cell Analysis System for Examining the Cytotoxicity of Drugs to Cancer Cell Lines. Exp. Ther. Med. 2017, 14, 1866–1870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franken, N.A.P.; Rodermond, H.M.; Stap, J.; Haveman, J.; van Bree, C. Clonogenic Assay of Cells in Vitro. Nat. Protoc. 2006, 1, 2315–2319. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence, 5′-3′ | Molecular Mass | |
|---|---|---|---|
| Calculated | Found * | ||
| Waz-5B12 | 5′-B12-L1-GGGUUCUACGAUAAACGGUUAAUGACCAGCUUAUGGCUGGCAGUUCCC | 15,232.1 | 15,279.1 (+2Na+) |
| Waz-3B12 | 5′-GGGUUCUACGAUAAACGGUUAAUGACCAGCUU-AUGGCUGGCAGUUCCC-L2-B12 | 15,545.3 | 15,569.8 (+Na+) |
| GL44-5B12 | 5′-B12-L1-ACGUUACUCUUGCAACACCCAAACUUUAA-UAGCCUCUUAUAGUUC | 14,355.6 | 14,379.7 (+Na+) |
| GL44-3B12 | 5′-ACGUUACUCUUGCAACACCCAAACUUUAA-UAGCCUCUUAUAGUUC-L2-B12 | 14,668.8 | 14,714.3 (+2Na+) |
| Fscr-5B12 | 5′-B12-L1-ACUGGUAUGUCGAGCCAACAAUCGAUACCAAGACUAAGA | 12,712.8 | 12,713.6 |
| Fscr-3B12 | 5′-ACUGGUAUGUCGAGCCAACAAUCGAUACCAA-GACUAAGA-L2-B12 | 13,025.9 | 13,048.6 (+Na+) |
 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Novopashina, D.S.; Dymova, M.A.; Davydova, A.S.; Meschaninova, M.I.; Malysheva, D.O.; Kuligina, E.V.; Richter, V.A.; Kolesnikov, I.A.; Taskaev, S.Y.; Vorobyeva, M.A. Aptamers for Addressed Boron Delivery in BNCT: Effect of Boron Cluster Attachment Site on Functional Activity. Int. J. Mol. Sci. 2023, 24, 306. https://doi.org/10.3390/ijms24010306
Novopashina DS, Dymova MA, Davydova AS, Meschaninova MI, Malysheva DO, Kuligina EV, Richter VA, Kolesnikov IA, Taskaev SY, Vorobyeva MA. Aptamers for Addressed Boron Delivery in BNCT: Effect of Boron Cluster Attachment Site on Functional Activity. International Journal of Molecular Sciences. 2023; 24(1):306. https://doi.org/10.3390/ijms24010306
Chicago/Turabian StyleNovopashina, Darya S., Maya A. Dymova, Anna S. Davydova, Mariya I. Meschaninova, Daria O. Malysheva, Elena V. Kuligina, Vladimir A. Richter, Iaroslav A. Kolesnikov, Sergey Yu. Taskaev, and Mariya A. Vorobyeva. 2023. "Aptamers for Addressed Boron Delivery in BNCT: Effect of Boron Cluster Attachment Site on Functional Activity" International Journal of Molecular Sciences 24, no. 1: 306. https://doi.org/10.3390/ijms24010306
APA StyleNovopashina, D. S., Dymova, M. A., Davydova, A. S., Meschaninova, M. I., Malysheva, D. O., Kuligina, E. V., Richter, V. A., Kolesnikov, I. A., Taskaev, S. Y., & Vorobyeva, M. A. (2023). Aptamers for Addressed Boron Delivery in BNCT: Effect of Boron Cluster Attachment Site on Functional Activity. International Journal of Molecular Sciences, 24(1), 306. https://doi.org/10.3390/ijms24010306