
Consistent DNA Hypomethylations in Prostate Cancer
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
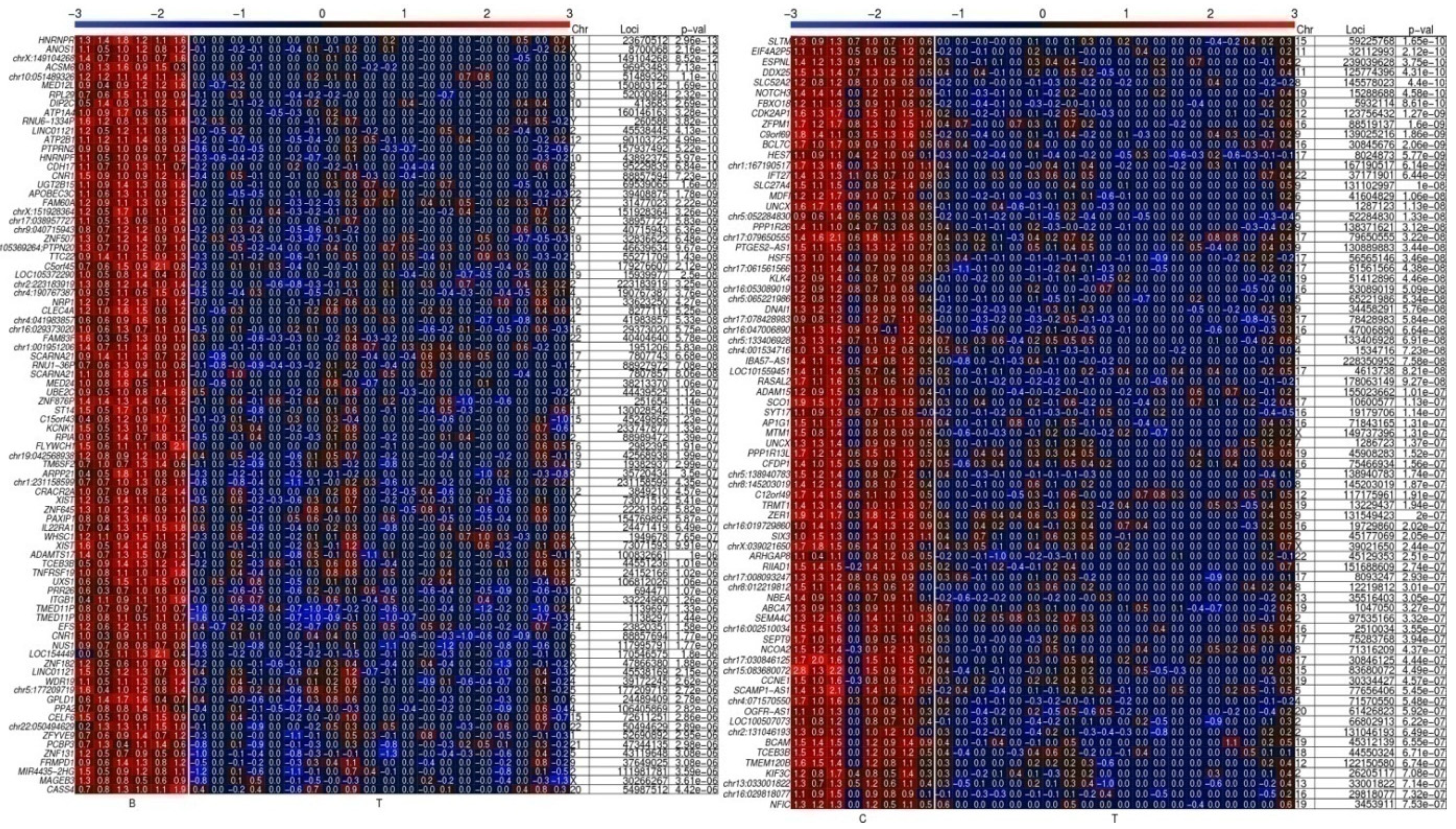
Distinct Hypomethylated 5′ Gene Regions in Tissue Specimens of PCa Patients
3. Discussion
4. Materials and Methods
4.1. Cell Line and Tissue Samples
- No. 1: University Hospital of Düsseldorf (UKD), Tumorbank
- 2.
- No. 2: University Hospital, Clinic of Urology, Tübingen
- 3.
- No. 3: Erasmus Medical Center Rotterdam
4.2. Methylated DNA Immunoprecipitation (MeDIP)
4.3. Amplification and Labelling of DNA
4.4. Hybridization of Microarrays
4.5. DNA Methylation Microarray Scanning and Data Analyses
4.6. Bisulfite Genomic Sequencing
4.7. Panther Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grossman, D.; Curry, S.; Owens, D.; Bibbins-Domingo, K.; Caughey, A.; Davidson, K.; Doubeni, C.; Ebell, M.; Epling, J.J.; Kemper, A.; et al. Screening for Prostate Cancer: US Preventive Services Task Force Recommendation Statement. JAMA 2018, 319, 1901–1913. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, S.; Vickers, A. Screening for Prostate Cancer. Med. Clin. N. Am. 2020, 104, 1051–1062. [Google Scholar] [CrossRef] [PubMed]
- Roobol, M.J. The Prostate-Specific Antigen Test. Expert Opin. Med. Diagn. 2013, 7, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, S.; Bashir, M.; Yaginuddin, A. Utility of DNA Methylation Markers for Diagnosing Cancer. Int. J. Surg. 2010, 8, 194–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidransky, D. Emerging Molecular Markers of Cancer. Nat. Rev. Cancer 2002, 2, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Bird, A. DNA Methylation Patterns and Epigenetic Memory. Genes Dev. 2002, 16, 6–21. [Google Scholar] [CrossRef] [Green Version]
- Luu, P.L.; Schöler, H.R.; Araúzo-Bravo, M.J. Disclosing the crosstalk among DNA methylation, transcription factors, and histone marks in human pluripotent cells through discovery of DNA methylation motifs. Genome Res. 2013, 23, 2013–2029. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.; Maié, T.; Dahl, E.; Costa, I.; Wagner, W. Deconvolution of Cellular Subsets in Human Tissue Based on Targeted DNA Methylation Analysis at Individual CpG Sites. BMC Biol. 2020, 18, 178. [Google Scholar] [CrossRef]
- Giovanni, C.; Catalona, C.; Guarrera, S.; Allione, A.; Casalone, E.; Russo, A.; Grosso, F.; Ferrante, D.; Viberti, C.; Aspesi, A.; et al. DNA Methylation of FKBP5 as Predictor of Overall Survival in Malignant Pleural Mesothelioma. Cancers 2020, 12, 3470. [Google Scholar] [CrossRef]
- Dor, Y.; Cedar, H. Principles of DNA Methylation and Their Implications for Biology and Medicine. Lancet 2018, 392, 777–786. [Google Scholar] [CrossRef]
- Chiam, K.; Ricciardelli, C.; Bianco-Miotto, T. Epigenetic Biomarkers in Prostate Cancer: Current and Future Uses. Cancer Lett. 2014, 342, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y. Promoter Hypermethylation in Prostate Cancer. Cancer Control 2010, 17, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Wu, Y.-P.; Ke, Z.-B.; Liang, Y.-C.; Cai, H.; Su, W.-T.; Tao, X.; Chen, S.-H.; Zheng, Q.-S.; Wei, Y.; et al. Identification of Key DNA Methylation-Driven Genes in Prostate Adenocarcinoma: An Integrative Analysis of TCGA Methylation Data. J. Transl. Med. 2019, 17, 311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devaney, J.M.; Wang, S.; Funda, S.; Long, J.; Taghipour, D.J.; Tbaishat, R.; Furbert-Harris, P.; Ittmann, M.; Kwabi-Addo, B. Identification of Novel DNA-Methylated Genes That Correlate with Human Prostate Cancer and High-Grade Prostatic Intraepithelial Neoplasia. Prostate Cancer Prostatic Dis. 2013, 16, 292–300. [Google Scholar] [CrossRef] [Green Version]
- Geybels, M.S.; Zhao, S.; Wong, C.-J.; Bibikova, M.; Klotzle, B.; Wu, M.; Ostrander, E.A.; Fan, J.-B.; Feng, Z.; Stanford, J. Epigenomic Profiling of DNA Methylation in Paired Prostate Cancer versus Adjacent Benign Tissue. Prostate 2015, 75, 1941–1950. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.W.; Kim, S.-T.; Turner, A.R.; Young, T.; Smith, S.; Liu, W.; Lindberg, J.; Egevad, L.; Gronberg, H.; Isaacs, W.B.; et al. Identification of New Differentially Methylated Genes That Have Potential Functional Consequences in Prostate Cancer. PLoS ONE 2012, 7, e48455. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Park, J.Y. DNA Methylation in Promoter Region as Biomarkers in Prostate Cancer. Methods Mol. Biol. 2012, 863, 67–109. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, M. DNA Hypomethylation in Cancer Cells. Epigenomics 2009, 1, 239–259. [Google Scholar] [CrossRef] [Green Version]
- Ateeq, B.; Unterberger, A.; Szyf, M.; Rabbani, S.A. Pharmacological Inhibition of DNA Methylation Induces Proinvasive and Prometastatic Genes in Vitro and in Vivo. Neoplasia 2008, 10, 266–278. [Google Scholar] [CrossRef]
- Schulz, W.A.; Steinhoff, C.; Florl, A.R. Methylation of Endogenous Human Retroelements in Health and Disease. Curr. Top. Microbiol. Immunol. 2006, 310, 211–250. [Google Scholar] [CrossRef] [PubMed]
- Santourlidis, S.; Florl, A.; Ackermann, R.; Wirtz, H.C.; Schulz, W.A. High Frequency of Alterations in DNA Methylation in Adenocarcinoma of the Prostate. Prostate 1999, 39, 166–174. [Google Scholar] [CrossRef]
- The Cancer Genome Atlas Research Network. The Molecular Taxonomy of Primary Prostate Cancer. Cell 2015, 163, 1011–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Results Referred to Here Are in Whole or Part Based Upon Data Generated by the TCGA Research Network. Available online: https://www.cancer.gov/tcga (accessed on 31 May 2022).
- Park, E.M.; Scott, P.M.; Clutario, K.; Cassidy, K.B.; Zhan, K.; Gerber, S.A.; Holland, A.J. WBP11 is required for splicing the TUBGCP6 pre-mRNA to promote centriole duplication. J. Cell Biol. 2020, 219, e201904203. [Google Scholar] [CrossRef] [PubMed]
- Pudova, E.A.; Lukyanova, E.N.; Nyushko, K.M.; Mikhaylenko, D.; Zaretsky, A.R.; Snezhkina, A.; Savvateeva, M.V.; Kobelyatskaya, A.; Melnikova, N.; Volchenko, N.N.; et al. Differentially Expressed Genes Associated With Prognosis in Locally Advanced Lymph Node-Negative Prostate Cancer. Front. Genet. 2019, 10, 730. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Bunn, P.A.; Zhou, C.; Chan, D. KIR 2D (L1, L3, L4, S4) and KIR 3DL1 protein expression in non-small cell lung cancer. Oncotarget 2016, 7, 82104–82111. [Google Scholar] [CrossRef] [Green Version]
- Seruggia, D.; Oti, M.; Tripathi, P.; Canver, M.C.; LeBlanc, L.; Di Giammartino, D.C.; Bullen, M.J.; Nefzger, C.M.; Sun, Y.B.Y.; Farouni, R.; et al. TAF5L and TAF6L Maintain Self-Renewal of Embryonic Stem Cells via the MYC Regulatory Network. Mol. Cell 2019, 74, 1148–1163.e1147. [Google Scholar] [CrossRef]
- Kim, T.; Jeong, K.; Kim, E.; Yoon, K.; Choi, J.; Park, J.H.; Kim, J.H.; Kim, H.S.; Youn, H.D.; Cho, E.J. Menin Enhances Androgen Receptor-Independent Proliferation and Migration of Prostate Cancer Cells. Mol. Cells 2022, 45, 202–215. [Google Scholar] [CrossRef]
- Li, Y.; Su, J.; DingZhang, X.; Zhang, J.; Yoshimoto, M.; Liu, S.; Bijian, K.; Gupta, A.; Squire, J.A.; AlaouiJamali, M.A.; et al. PTEN deletion and heme oxygenase-1 overexpression cooperate in prostate cancer progression and are associated with adverse clinical outcome. J. Pathol. 2011, 224, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhao, H.; Li, J.; Liu, H.; Wang, F.; Wei, Y.; Su, J.; Zhang, D.; Liu, T.; Zhang, Y. The identification of specific methylation patterns across different cancers. PLoS ONE 2015, 10, e0120361. [Google Scholar] [CrossRef]
- Stopsack, K.H.; Gerke, T.A.; Andrén, O.; Andersson, S.O.; Giovannucci, E.L.; Mucci, L.A.; Rider, J.R. Cholesterol uptake and regulation in high-grade and lethal prostate cancers. Carcinogenesis 2017, 38, 806–811. [Google Scholar] [CrossRef] [Green Version]
- Shafran, J.S.; Jafari, N.; Casey, A.N.; Győrffy, B.; Denis, G.V. BRD4 regulates key transcription factors that drive epithelial-mesenchymal transition in castration-resistant prostate cancer. Prostate Cancer Prostatic Dis. 2021, 24, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Zong, L.; Hattori, N.; Yoda, Y.; Yamashita, S.; Takeshima, H.; Takahashi, T.; Maeda, M.; Katai, H.; Nanjo, S.; Ando, T.; et al. Establishment of a DNA methylation marker to evaluate cancer cell fraction in gastric cancer. Gastric Cancer 2016, 19, 361–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.F.; Cai, X.L.; Li, Z.Z.; Lv, J.; Xiang, Y.A.; Chen, J.J.; Chen, W.J.; Sun, W.Y.; Liu, X.M.; Chen, J.B. LncRNA SNHG16 Functions as an Oncogene by Sponging MiR-4518 and Up-Regulating PRMT5 Expression in Glioma. Cell Physiol. Biochem. 2018, 45, 1975–1985. [Google Scholar] [CrossRef] [PubMed]
- Nagano, K.; Yamashita, T.; Inoue, M.; Higashisaka, K.; Yoshioka, Y.; Abe, Y.; Mukai, Y.; Kamada, H.; Tsutsumi, Y.; Tsunoda, S. Eph Receptor A10 Has a Potential as a Target for a Prostate Cancer Therapy. Biochem. Biophys. Res. Commun. 2014, 450, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Perrouin-Verbe, M.A.; Schoentgen, N.; Talagas, M.; Garlantezec, R.; Uguen, A.; Doucet, L.; Rosec, S.; Marcorelles, P.; Potier-Cartereau, M.; Vandier, C.; et al. Overexpression of certain transient receptor potential and Orai channels in prostate cancer is associated with decreased risk of systemic recurrence after radical prostatectomy. Prostate 2019, 79, 1793–1804. [Google Scholar] [CrossRef]
- Zolochevska, O.; Figueiredo, M.L. Cell Cycle Regulator Cdk2ap1 Inhibits Prostate Cancer Cell Growth and Modifies Androgen-Responsive Pathway Function. Prostate 2009, 69, 1586–1597. [Google Scholar] [CrossRef]
- Erichsen, L.; Ghanjati, F.; Beermann, A.; Poyet, C.; Hermanns, T.; Schulz, W.A.; Seifert, H.-H.; Wild, P.J.; Buser, L.; Kröning, A.; et al. Aberrant Methylated Key Genes of Methyl Group Metabolism within the Molecular Etiology of Urothelial Carcinogenesis. Sci. Rep. 2018, 8, 3477. [Google Scholar] [CrossRef] [Green Version]
- Erichsen, L.; Seifert, H.-H.; Schulz, W.A.; Hoffmann, M.J.; Niegisch, G.; Araúzo-Bravo, M.J.; Bendhack, M.L.; Poyet, C.; Hermanns, T.; Beermann, A.; et al. Basic Hallmarks of Urothelial Cancer Unleashed in Primary Uroepithelium by Interference with the Epigenetic Master Regulator ODC1. Sci. Rep. 2020, 10, 3808. [Google Scholar] [CrossRef] [Green Version]
- Santourlidis, S.; Trompeter, H.-I.; Weinhold, S.; Eisermann, B.; Meyer, K.L.; Wernet, P.; Uhrberg, M. Crucial Role of DNA Methylation in Determination of Clonally Distributed Killer Cell Ig-like Receptor Expression Patterns in NK Cells. J. Immunol. 2002, 169, 4253–4261. [Google Scholar] [CrossRef]
- Santourlidis, S.; Graffmann, N.; Christ, J.; Uhrberg, M. Lineage-Specific Transition of Histone Signatures in the Killer Cell Ig-like Receptor Locus from Hematopoietic Progenitor to NK Cells. J. Immunol. 2008, 180, 418–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mi, H.; Muruganujan, A.; Huang, X.; Ebert, D.; Mills, C.; Guo, X.; Thomas, P.D. Protocol Update for Large-Scale Genome and Gene Function Analysis with PANTHER Classification System (v.14.0). Nat. Protoc. 2019, 14, 703–721. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Ebert, D.; Muruganujan, A.; Mills, C.; Albou, L.-P.; Mushayamaha, T.; Thomas, P.D. PANTHER Version 16: A Revised Family Classification, Tree-Based Classification Tool, Enhancer Regions and Extensive API. Nucleic Acids Res. 2021, 49, D394–D403. [Google Scholar] [CrossRef] [PubMed]
- Pedrosa, A.R.; Graça, J.L.; Carvalho, S.; Peleteiro, M.C.; Duarte, A.; Trindade, A. Notch signaling dynamics in the adult healthy prostate and in prostatic tumor development. Prostate 2016, 76, 80–96. [Google Scholar] [CrossRef]
- Tse, B.W.-C.; Kryza, T.; Yeh, M.-C.; Dong, Y.; Sokolowski, K.A.; Walpole, C.; Dreyer, T.; Felber, J.; Harris, J.; Magdolen, V.; et al. KLK4 Induces Anti-Tumor Effects in Human Xenograft Mouse Models of Orthotopic and Metastatic Prostate Cancer. Cancers 2020, 12, 3501. [Google Scholar] [CrossRef]
- Burdelski, C.; Fitzner, M.; HubeMagg, C.; Kluth, M.; Heumann, A.; Simon, R.; Krech, T.; Clauditz, T.; Büscheck, F.; Steurer, S.; et al. Overexpression of the A Disintegrin and Metalloproteinase ADAM15 Is Linked to a Small but Highly Aggressive Subset of Prostate Cancers. Neoplasia 2017, 19, 279–287. [Google Scholar] [CrossRef]
- Qin, G.Q.; He, H.C.; Han, Z.D.; Liang, Y.X.; Yang, S.B.; Huang, Y.Q.; Zhou, L.; Fu, H.; Li, J.X.; Jiang, F.N.; et al. Combined overexpression of HIVEP3 and SOX9 predicts unfavorable biochemical recurrence-free survival in patients with prostate cancer. OncoTargets Ther. 2014, 7, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, L.; Chen, S.; Peng, H.; Xiao, L.; Du, E.; Liu, Y.; Lin, D.; Wang, Y.; Xu, Y.; et al. PKMYT1 is associated with prostate cancer malignancy and may serve as a therapeutic target. Gene 2020, 744, 144608. [Google Scholar] [CrossRef]
- Li, D.; Liu, J.; Huang, S.; Bi, X.; Wang, B.; Chen, Q.; Chen, H.; Pu, X. CCAAT enhancer binding protein β promotes tumor growth and inhibits apoptosis in prostate cancer by methylating estrogen receptor β. Neoplasma 2018, 65, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Cai, Q.; He, W.; Lam, T.B.; Lin, J.; Zhao, Y.; Chen, X.; Gu, P.; Huang, H.; Xue, M.; et al. AP4 modulated by the PI3K/AKT pathway promotes prostate cancer proliferation and metastasis of prostate cancer via upregulating L-plastin. Cell Death Dis. 2017, 8, e3060. [Google Scholar] [CrossRef]
- Zhu, A.; Hopkins, K.M.; Friedman, R.A.; Bernstock, J.D.; Broustas, C.G.; Liebermann, H.B. DNMT1 and DNMT3B Regulate Tumorigenicity of Human Prostate Cancer Cells by Controlling RAD9 Expression through Targeted Methylation. Carcinogenesis 2021, 42, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Weizhong, Z.; Shuohui, G.; Hanjiao, Q.; Yuhong, M.; Xiaohua, Y.; Jian, C.; Lisen, L. Inhibition of cytohesin-1 by siRNA leads to reduced IGFR signaling in prostate cancer. Braz. J. Med. Biol. Res. 2011, 44, 642–646. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Wilfred, P.; Korbie, D.; Trau, M. Regulation of Canonical Oncogenic Signaling Pathways in Cancer via DNA Methylation. Cancers 2020, 12, 3199. [Google Scholar] [CrossRef] [PubMed]
- Irizarry, R.; Ladd-Acosta, C.; Wen, B.; Wu, Z.; Montano, C.; Onyango, P.; Cui, H.; Gabo, K.; Rongione, M.; Webster, M.; et al. The Human Colon Cancer Methylome Shows Similar Hypo- and Hypermethylation at Conserved Tissue-Specific CpG Island Shores. Nat. Genet. 2009, 41, 178–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertoli, G.; Cava, C.; Castiglioni, I. MicroRNAs as Biomarkers for Diagnosis, Prognosis and Theranostics in Prostate Cancer. Int. J. Mol. Sci. 2016, 17, 421. [Google Scholar] [CrossRef] [Green Version]
- Filipska, M.; Skrzypski, M.; Czetyrbok, K.; Stokowy, T.; Stasiłojć, G.; Supernat, A.; Jassem, J.; Żaczek, A.J.; Bigda, J. MiR-192 and MiR-662 Enhance Chemoresistance and Invasiveness of Squamous Cell Lung Carcinoma. Lung Cancer 2018, 118, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Murillo-Garzón, V.; Kypta, R. WNT Signalling in Prostate Cancer. Nat. Rev. Urol. 2017, 14, 683–696. [Google Scholar] [CrossRef]
- Lee, D. MiR-769-5p Is Associated with Prostate Cancer Recurrence and Modulates Proliferation and Apoptosis of Cancer Cells. Exp. Ther. Med. 2021, 21, 335. [Google Scholar] [CrossRef]
- Zhang, S.; Wan, H.; Zhang, X. LncRNA LHFPL3-AS1 Contributes to Tumorigenesis of Melanoma Stem Cells via the MiR-181a-5p/BCL2 Pathway. Cell Death Dis. 2020, 11, 950. [Google Scholar] [CrossRef]
- Ding, M.; Fu, Y.; Guo, F.; Chen, H.; Fu, X.; Tan, W.; Zhang, H. Long Non-Coding RNA MAFG-AS1 Knockdown Blocks Malignant Progression in Breast Cancer Cells by Inactivating JAK2/STAT3 Signaling Pathway via MAFG-AS1/MiR-3196/TFAP2A Axis. Int. J. Clin. Exp. Pathol. 2020, 13, 2455–2473. [Google Scholar]
- Ruan, Z.; Deng, H.; Liang, M.; Xu, Z.; Lai, M.; Ren, H.; Deng, X.; Su, X. Downregulation of Long Non-Coding RNA MAFG-AS1 Represses Tumorigenesis of Colorectal Cancer Cells through the MicroRNA-149-3p-Dependent Inhibition of HOXB8. Cancer Cell Int. 2020, 20, 511. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.-W.; Kurago, Z.B.; Stewart, C.A.; Wilson, M.J.; Martin, M.P.; Mace, B.E.; Carrington, M.; Trowsdale, J.; Lutz, C.T. DNA Methylation Maintains Allele-Specific KIR Gene Expression in Human Natural Killer Cells. J. Exp. Med. 2003, 197, 245–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, R.; Khan, A.P.; Asangani, I.A.; Cieślik, M.; Prensner, J.R.; Wang, X.; Iyer, M.K.; Jiang, X.; Borkin, D.; Escara-Wilke, J.; et al. Targeting the MLL Complex in Castration-Resistant Prostate Cancer. Nat. Med. 2015, 21, 344–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tirado-Magallanes, R.; Rebbani, K.; Lim, R.; Pradhan, S.; Benoukraf, T. Whole Genome DNA Methylation: Beyond Genes Silencing. Oncotarget 2017, 8, 5629–5637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharifi-Zarchi, A.; Gerovska, D.; Adachi, K.; Totonchi, M.; Pezeshk, H.; Taft, R.J.; Schöler, H.R.; Chitsaz, H.; Sadeghi, M.; Baharvand, H.; et al. DNA methylation regulates discrimination of enhancers from promoters through a H3K4me1-H3K4me3 seesaw mechanism. BMC Genom. 2017, 12, 964. [Google Scholar] [CrossRef]
- Luu, P.L.; Gerovska, D.; Schöler, H.R.; Araúzo-Bravo, M.J. Rules governing the mechanism of epigenetic reprogramming memory. Epigenomics 2018, 10, 149–174. [Google Scholar] [CrossRef]
- Ghanjati, F.; Beermann, A.; Hermanns, T.; Poyet, C.; Araúzo-Bravo, M.J.; Seifert, H.-H.; Schmidtpeter, M.; Goering, W.; Sorg, R.; Wernet, P.; et al. Unreserved Application of Epigenetic Methods to Define Differences of DNA Methylation between Urinary Cellular and Cell-Free DNA. Cancer Biomark. 2014, 14, 295–302. [Google Scholar] [CrossRef]
- Santourlidis, S.; Ghanjati, F.; Beermann, A.; Hermanns, T.; Poyet, C. IDLN-MSP: Idiolocal Normalization of Real-Time Methylation-Specific PCR for Genetic Imbalanced DNA Specimens. BioTechniques 2016, 60, 84–87. [Google Scholar] [CrossRef] [Green Version]
- Ebrahim, S.; Avenarius, M.R.; Grati, M.; Krey, J.F.; Windsor, A.M.; Sousa, A.D.; Ballesteros, A.; Cui, R.; Millis, B.A.; Salles, F.T.; et al. Stereocilia-Staircase Spacing Is Influenced by Myosin III Motors and Their Cargos Espin-1 and Espin-Like. Nat. Commun. 2016, 1, 10833. [Google Scholar] [CrossRef] [Green Version]
- Guichard, C.; Harricane, M.C.; Lafitte, J.J.; Godard, P.; Zaegel, M.; Tack, V.; Lalau, G.; Bouvagnet, P. Axonemal Dynein Intermediate-Chain Gene (DNAI1) Mutations Result in Situs Inversus and Primary Ciliary Dyskinesia (Kartagener Syndrome). Am. J. Hum. Genet. 2001, 68, 1030–1035. [Google Scholar] [CrossRef] [Green Version]
- Ruhl, D.A.; Bomba-Warczak, E.; Watson, E.T.; Bradberry, M.M.; Peterson, T.A.; Frelka, A.; Evans, C.S.; Briguglio, J.S.; Basta, T.; Stowell, M.H.; et al. Synaptotagmin 17 Controls Neurite Outgrowth and Synaptic Physiology via Distinct Cellular Pathways. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Diekwisch, T.G.; Marches, F.; Williams, A.; Luan, X. Cloning, Gene Expression, and Characterization of CP27, a Novel Gene in Mouse Embryogenesis. Gene 1999, 235, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Messina, G.; Atterrato, M.T.; Prozzillo, Y.; Piacentini, L.; Losada, A.; Dimitri, P. The Human Cranio Facial Development Protein 1 (Cfdp1) Gene Encodes a Protein Required for the Maintenance of Higher-Order Chromatin Organization. Sci. Rep. 2017, 7, 45022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Yang, X.; Guan, H.; Mizokami, A.; Keller, E.; Xu, X.; Liu, X.; Tan, J.; Hu, L.; Lu, Y.; et al. Exosome-Derived MicroRNAs Contribute to Prostate Cancer Chemoresistance. J. Clin. Oncol. 2016, 49, 838–846. [Google Scholar] [CrossRef] [Green Version]
- Morales, S.; Monzo, M.; Navarro, A. Epigenetic Regulation Mechanisms of MicroRNA Expression. Biomolecular Concepts 2017, 8, 203–212. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, M.; Ding, Y.; Fan, Z.; Zhang, J.; Zhang, H.; Jiang, B.; Zhu, Y. Serum MicroRNA Profile in Patients with Colon Adenomas or Cancer. BMC Med. Genom. 2017, 10, 23. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Yang, P.; Zhang, B.; Yao, M.; Jia, Y.; Li, N.; Liu, H.; Bai, H.; Gong, X. Long Noncoding RNA TTC39A-AS1 Promotes Breast Cancer Tumorigenicity by Sponging MicroRNA-483-3p and Thereby Upregulating MTA2. Pharmacology 2021, 106, 573–587. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Feng, Y.; Li, F.; Jia, Y.; Peng, Y.; Zhao, W.; Hu, J.; He, A. LncRNA ST3GAL6-AS1 Promotes Invasion by Inhibiting HnRNPA2B1-mediated ST3GAL6 Expression in Multiple Myeloma. Int. J. Oncol. 2021, 58, 5. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, S.S.; Hostetter, G.; Tang, L.; Frank, S.B.; Saboda, K.; Mehra, R.; Wang, L.; Li, X.; Keller, E.T.; Miranti, C.K. Notch3 Promotes Prostate Cancer-Induced Bone Lesion Development via MMP-3. Oncogene 2019, 39, 204–218. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.R.; Gu, M.J. The Clinicopathologic Significance of Notch3 Expression in Prostate Cancer. Int. J. Clin. Exp. Pathol. 2019, 12, 3535–3541. [Google Scholar]
- Najy, A.J.; Day, K.C.; Day, M.L. ADAM15 Supports Prostate Cancer Metastasis by Modulating Tumor Cell-Endothelial Cell Interaction. Cancer Res. 2008, 68, 1092–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farah, E.; Zhang, Z.; Utturkar, S.M.; Liu, J.; Ratliff, T.L.; Liu, X. Targeting DNMTs to Overcome Enzalutamide Resistance in Prostate Cancer. Mol. Cancer Ther. 2022, 21, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Infante, A.; Gener, B.; Vázquez, M.; Olivares, N.; Arrieta, A.; Grau, G.; Llano, I.; Madero, L.; Bueno, A.M.; Sagastizabal, B.; et al. Reiterative infusions of MSCs improve pediatric osteogenesis imperfecta eliciting a pro-osteogenic paracrine response: TERCELOI clinical trial. Clin. Transl. Med. 2021, 11, e265. [Google Scholar] [CrossRef] [PubMed]
- Ascension, A.M.; Araúzo-Bravo, M.J. BigMPI4py: Python Module for Parallelization of Big Data Objects Discloses Germ Layer Specific DNA Demethylation Motifs. IEEE/ACM Trans. Comput. Biol. Bioinform. 2022, 19, 1507–1522. [Google Scholar] [CrossRef] [PubMed]
- Dorn, I.; Klich, K.; Araúzo-Bravo, M.J. Erythroid Differentiation of Human Induced Pluripotent Stem Cells Is Independent of Donor Cell Type of Origin. Haematologica 2015, 100, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Hargus, G.; Ehrlich, M.; Araúzo-Bravo, M.J.; Hemmer, K.; Hallmann, A.-L.; Reinhardt, P.; Kim, K.-P.; Adachi, K.; Santourlidis, S.; Ghanjati, F.; et al. Origin-Dependent Neural Cell Identities in Differentiated Human IPSCs in Vitro and after Transplantation into the Mouse Brain. Cell Rep. 2014, 25, 1697–1703. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol/ Name/ EntrezID/ (Chromosomal Location) | Function | Selected Publication on Gene’s Involvement in PCa and Cancer | Votes in 10 Tumors Compared to BPH | PCa Gene Expression |
|---|---|---|---|---|
| WBP11-Cor WW domain-binding protein 11 51,729 (12p12.3) | Nuclear protein. Binds to the Npw38WW domain activating pre-mRNA splicing | WBP11 is required for splicing the TUBGCP6 pre-mRNA to promote centriole duplication [25] | 9 | 1.07 |
| B4GALNT4-Ups ß-1,4-N-Acetyl-Galactosaminyltransferase 4 338,707 (11p15.5) | Transfers N-acetylgalactosamine (GalNAc) from UDP-GalNAc to N-acetylglucosamine-ß -benzyl | Differentially expressed genes associated with prognosis in locally advanced lymph node-negative PCa [26] | 8 | 3.66 |
| KIR2DL3-Cor Killer Cell Immunoglobulin-Like Receptor, Two Ig Domains And Long Cytoplasmic Tail 3 3804 (19q13.42) | Inhibits activity of NK cells, preventing cell lysis | KIR 2D (L1, L3, L4, S4) and KIR 3DL1 protein expression in non-small cell lung cancer [27] | 10 | 1.38 |
| TAF6L-Cor TATA-Box-Binding Protein Associated Factor 6-Like 10,629 (11q12.3) | Member of the PCAF complex, an epigenetic regulator essential for somatic reprogramming | TAF5L and TAF6L maintain self-renewal of embryonic stem cells via the MYC regulatory Network [28] | 10 | 1.36 |
| MEN1-Ups Menin 1 4221 (11q13.1) | Tumor suppressor functions in histone modification and epigenetic gene regulation by altering chromatin structure | Menin enhances androgen receptor-independent proliferation and migration of PCa cells [29] | 10 | 1.33 |
| HMOX1-Cor Heme Oxygenase 1 3162 (22q12.3) | HO catalyzes the degradation of heme. Has an anti-apoptotic function | PTEN deletion and heme oxygenase-1 overexpression cooperate in PCa progression and are associated with adverse clinical outcome [30] | 10 | 1.06 |
| WBSCR17-Cor Polypeptide N-Acetylgalactosaminyltransferase 17 64,409 (7q11.22) | Predicted to play a role in membrane trafficking | The identification of specific methylation patterns across different cancers [31] | 10 | 1.07 |
| NDUFB7-Cor NADH:Ubiquinone Oxidoreductase Subunit B7 4713 (19p13.12) | Subunit of NADH dehydrogenase (Complex I). Functions in the transfer of electrons from NADH to the respiratory chain | Cholesterol uptake and regulation in high-grade and lethal PCas [32] | 9 | 1.11 |
| BRD4-Cor Bromodomain-Containing 4 23,476 (19p13.12) | Chromatin reader. Recognizes and binds acetylated histones. Plays a key role in transmission of epigenetic memory across cell divisions and transcription regulation | BRD4 regulates key transcription factors that drive epithelial-mesenchymal transition in castration-resistant PCa [33] | 8 | 0.997 |
| PPFIA3-Cor PTPRF-Interacting Protein Alpha 3 8541 (19q13.33) | Localize receptor-like tyrosine phosphatases type 2A at specific sites on the plasma membrane | Establishment of a DNA methylation marker to evaluate cancer cell fraction in gastric cancer [34] | 8 | 1.35 |
| MIR4518-Cor microRNA 4518 100616405 (16p11.2) | Post-transcriptional regulation of gene expression | LncRNA SNHG16 functions as an oncogene by sponging MiR-4518 and up-regulating PRMT5 expression in glioma [35] | 10 | ND |
| EPH10 EPH Receptor A10 284,656 (1p34.3) | Receptor tyrosine kinases (RTKs), and their ephrin ligands, such as EPH10, are important mediators of cell–cell communication regulating cell attachment, shape, and mobility in neuronal and epithelial cells | Eph receptor A10 has a potential as a target for a PCa therapy [36] | 10 | 5.28 |
| ORAI3 ORAI Calcium Release-Activated Calcium Modulator 3 93,129 (16p11.2) | Ca(2+) release-activated Ca(2+)-like (CRAC-like) channel subunit which mediates Ca(2+) influx and increase in Ca(2+)-selective current by synergy with the Ca(2+) sensor, STIM1 | Overexpression of certain transient receptor potential and Orai channels in PCa is associated with decreased risk of systemic recurrence after radical prostatectomy [37] | 10 | 1.07 |
| CDK2AP1 Cyclin-Dependent Kinase 2 Associated Protein 1 8099 (12q24.31) | Specific inhibitor of the cell-cycle kinase CDK2. It plays a role in cell-cycle and epigenetic regulation | Cell cycle regulator cdk2ap1 inhibits PCa cell growth and modifies androgen-responsive pathway function [38] | 9 | 1.04 |
| Gene Symbol/ Name/ Entrez ID/ (Chromosomal Location) | Function | Selected Publication on Gene’s Involvement in PCa and Cancer | Votes in 20 Tumors Compared to Tumor-Adjacent Healthy Samples | PCa Gene Expression |
|---|---|---|---|---|
| NOTCH3 Notch Receptor 3 4854 (19p13.12) | Receptor for membrane-bound ligands Jagged1, Jagged2, and Delta1 to regulate cell-fate determination. Affects differentiation, proliferation, and apoptotic programs | Notch signaling dynamics in the adult healthy prostate and in prostatic tumor development [45] | 20 | 1.11 |
| CDK2AP1 Cyclin-Dependent Kinase 2 Associated Protein 1 8099 (12q24.31) | Forms a core subunit of the nucleosome remodeling and histone deacetylation (NURD) complex that epigenetically regulates embryonic stem cell differentiation. Specific inhibitor of the cell-cycle kinase CDK2 | Cell cycle regulator Cdk2ap1 inhibits PCa cell growth and modifies androgen-responsive pathway function [38] | 20 | 1.04 |
| KLK4 Kallikrein-Related Peptidase 4 9622 (19q13.41) | Serine protease with major role in enamel formation | KLK4 induces anti-tumor effects in human xenograft mouse models of orthotopic and metastatic PCa [46] | 20 | 1.86 |
| ADAM15 ADAM Metallopeptidase Domain 15 8751 (1q21.3) | Metalloproteinase with gelatinolytic and collagenolytic activity. Inhibits ß-1 integrin-mediated cell adhesion and migration of airway smooth muscle cells. Cleaves E-cadherin in response to growth factor deprivation | Overexpression of the A disintegrin and metalloproteinase ADAM15 is linked to a small but highly aggressive subset of PCas [47] | 20 | 1.32 |
| HIVEP3 Human Immunodeficiency Virus Type I Enhancer-Binding Protein 3 59,269 (1p34.2) | TF that regulates nuclear factor κB-mediated transcription by binding the κBmotif in target genes. Strongly inhibits TNF-α-induced NF-κB activation. Interaction with TRAF proteins inhibits both NF-κB-mediated and c-Jun N-terminal kinase/JNK-mediated responses that include apoptosis and proinflammatory cytokine gene expression | Combined overexpression of HIVEP3 and SOX9 predicts unfavorable biochemical recurrence-free survival in patients with PCa [48] | 20 | 1.17 |
| NDUFB7-Cor NADH:Ubiquinone Oxidoreductase Subunit B7 4713 (19p13.12) | Subunit of NADH dehydrogenase (Complex I) that functions in the transfer of electrons from NADH to the respiratory chain | Cholesterol uptake and regulation in high-grade and lethal PCas [32] | 20 | 1,11 |
| PPFIA3-Cor PTPRF-Interacting Protein Alpha 3 8541 (19q13.33) | Localize receptor-like tyrosine phosphatases type 2A at specific sites on the plasma membrane, possibly regulating their interaction with the extracellular environment and their association with substrates | Establishment of a DNA methylation marker to evaluate cancer cell fraction in gastric cancer [34] | 20 | 1.35 |
| MIR4518-Cor microRNA 4518 100616405 (16p11.2) | Post-transcriptional regulation of gene expression | LncRNA SNHG16 functions as an oncogene by sponging MiR-4518 and up-regulating PRMT5 expression in glioma [35] | 20 | ND |
| ORAI3 ORAI Calcium Release-Activated Calcium Modulator 3 93,129 (16p11.2) | Ca(2+) release-activated Ca(2+)-like (CRAC-like) channel subunit which mediates Ca(2+) influx and increase in Ca(2+)-selective current by synergy with the Ca(2+) sensor, STIM1 | Overexpression of certain transient receptor potential and Orai channels in PCa is associated with decreased risk of systemic recurrence after radical prostatectomy [37] | 20 | 1.07 |
| CCNE1 Cyclin E1 898 (19q12) | Essential for the control of the cell cycle at the G1/S (start) transition | PKMYT1 is associated with PCa malignancy and may serve as a therapeutic target [49] | 20 | 1.02 |
| CEBPB CCAAT Enhancer-Binding Protein Beta 1051 (20q13.13) | TF regulating expression of genes involved in immune and inflammatory responses. Promotes osteoblast differentiation and osteoclastogenesis | CCAAT enhancer-binding protein beta promotes tumor growth and inhibits apoptosis in PCa by methylating estrogen receptor beta [50] | 20 | 0.612 |
| REPIN1 Replication Initiator 1 29,803 (7q36.1) | Sequence-specific double-stranded DNA-binding protein required for initiation of chromosomal DNA replication | AP4 modulated by the PI3K/AKT pathway promotes PCa proliferation and metastasis of prostate cancer via upregulating L-plastin [51] | 20 | 1.48 |
| DNMT3B DNA Methyltransferase 3 Beta 1789 (20q11.21) | Required for genome-wide de novo methylation. Essential for the establishment of DNA methylation patterns during development | DNMT1 and DNMT3B regulate tumorigenicity of human PCa cells by controlling RAD9 expression through targeted methylation [52] | 20 | 1.37 |
| CYTH1 Cytohesin 1 9267 (17q25.3) | Promotes activation of ARF factors by replacement of GDP with GTP. Plays an important role in membrane trafficking during junctional remodeling and epithelial polarization | Inhibition of cytohesin-1 by siRNA leads to reduced IGFR signaling in PCa [53] | 20 | 1.1 |
| KEAP1 Kelch-Like ECH Associated Protein 1 9817 (19p13.2) | Substrate-specific adapter of a BCR E3 ubiquitin ligase complex. Regulates response to oxidative stress by targeting NFE2L2/NRF2 for ubiquitination | Regulation of canonical oncogenic signaling pathways in cancer via DNA methylation [54] | 20 | 1.11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Araúzo-Bravo, M.J.; Erichsen, L.; Ott, P.; Beermann, A.; Sheikh, J.; Gerovska, D.; Thimm, C.; Bendhack, M.L.; Santourlidis, S. Consistent DNA Hypomethylations in Prostate Cancer. Int. J. Mol. Sci. 2023, 24, 386. https://doi.org/10.3390/ijms24010386
Araúzo-Bravo MJ, Erichsen L, Ott P, Beermann A, Sheikh J, Gerovska D, Thimm C, Bendhack ML, Santourlidis S. Consistent DNA Hypomethylations in Prostate Cancer. International Journal of Molecular Sciences. 2023; 24(1):386. https://doi.org/10.3390/ijms24010386
Chicago/Turabian StyleAraúzo-Bravo, Marcos J., Lars Erichsen, Pauline Ott, Agnes Beermann, Jamal Sheikh, Daniela Gerovska, Chantelle Thimm, Marcelo L. Bendhack, and Simeon Santourlidis. 2023. "Consistent DNA Hypomethylations in Prostate Cancer" International Journal of Molecular Sciences 24, no. 1: 386. https://doi.org/10.3390/ijms24010386