
Bone Marrow-Derived Vasculogenic Mesenchymal Stem Cells Enhance In Vitro Angiogenic Sprouting of Human Umbilical Vein Endothelial Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
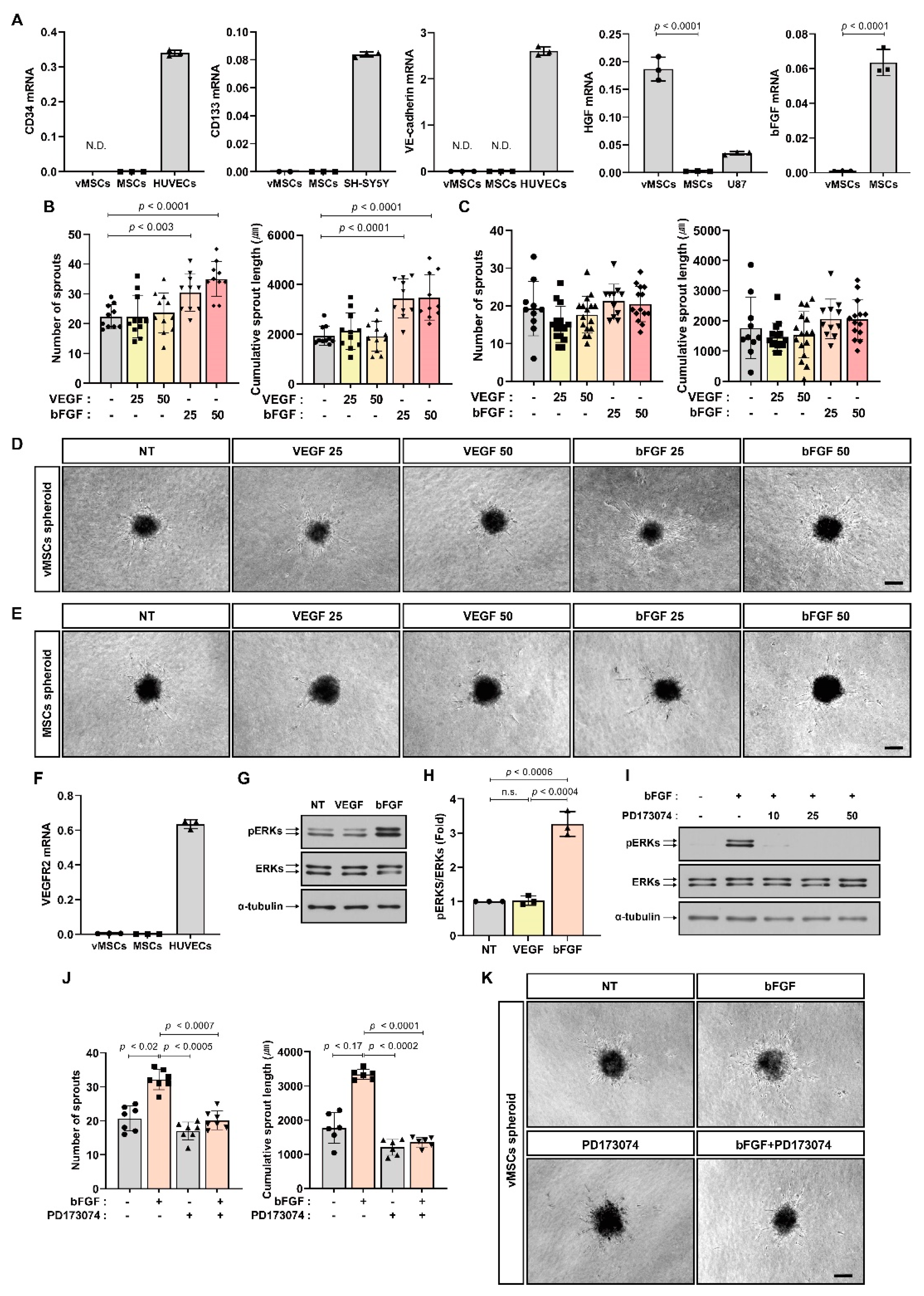
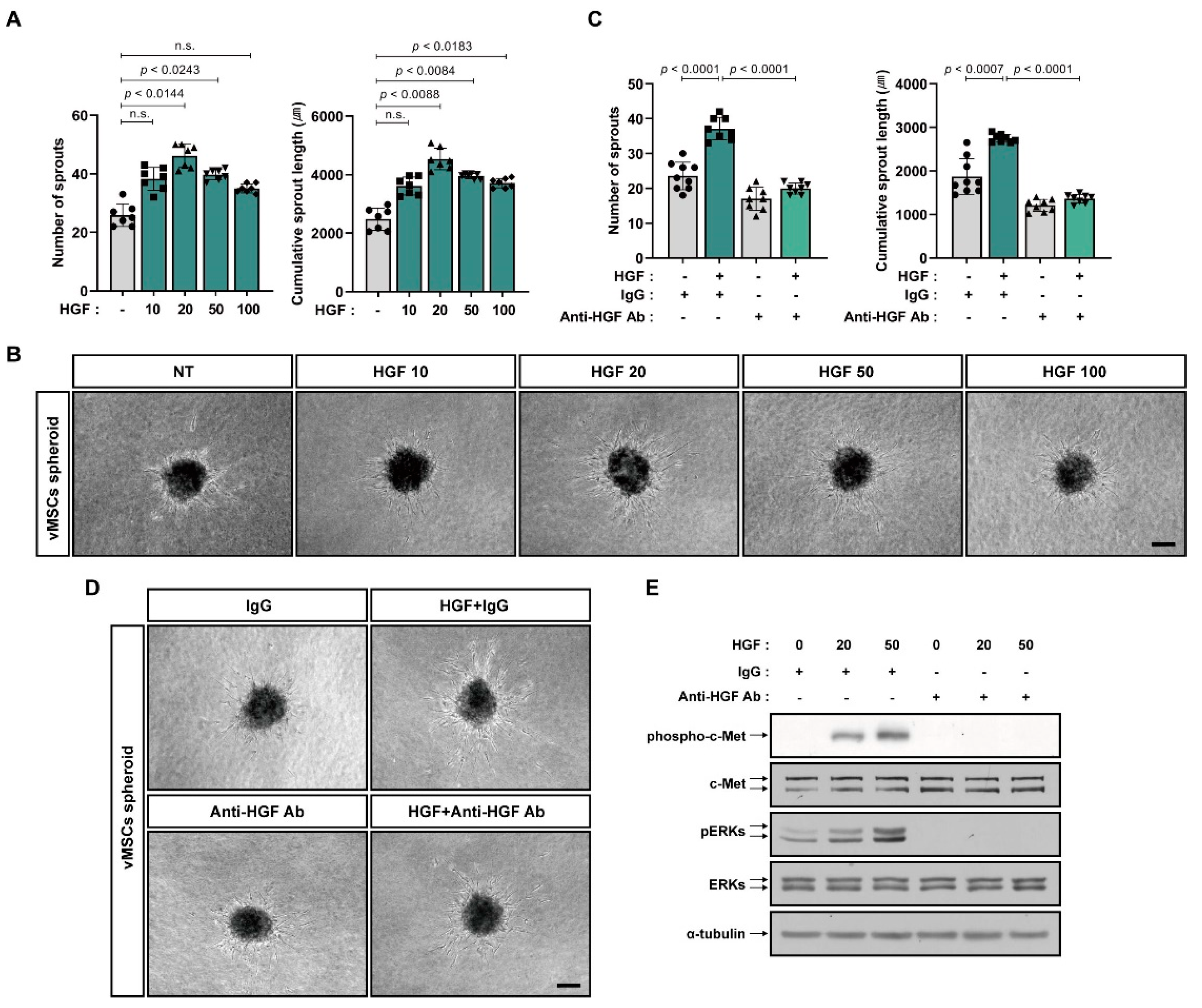
2.1. Increase in Angiogenic Sprouting Capacity of vMSCs in Response to bFGF and HGF
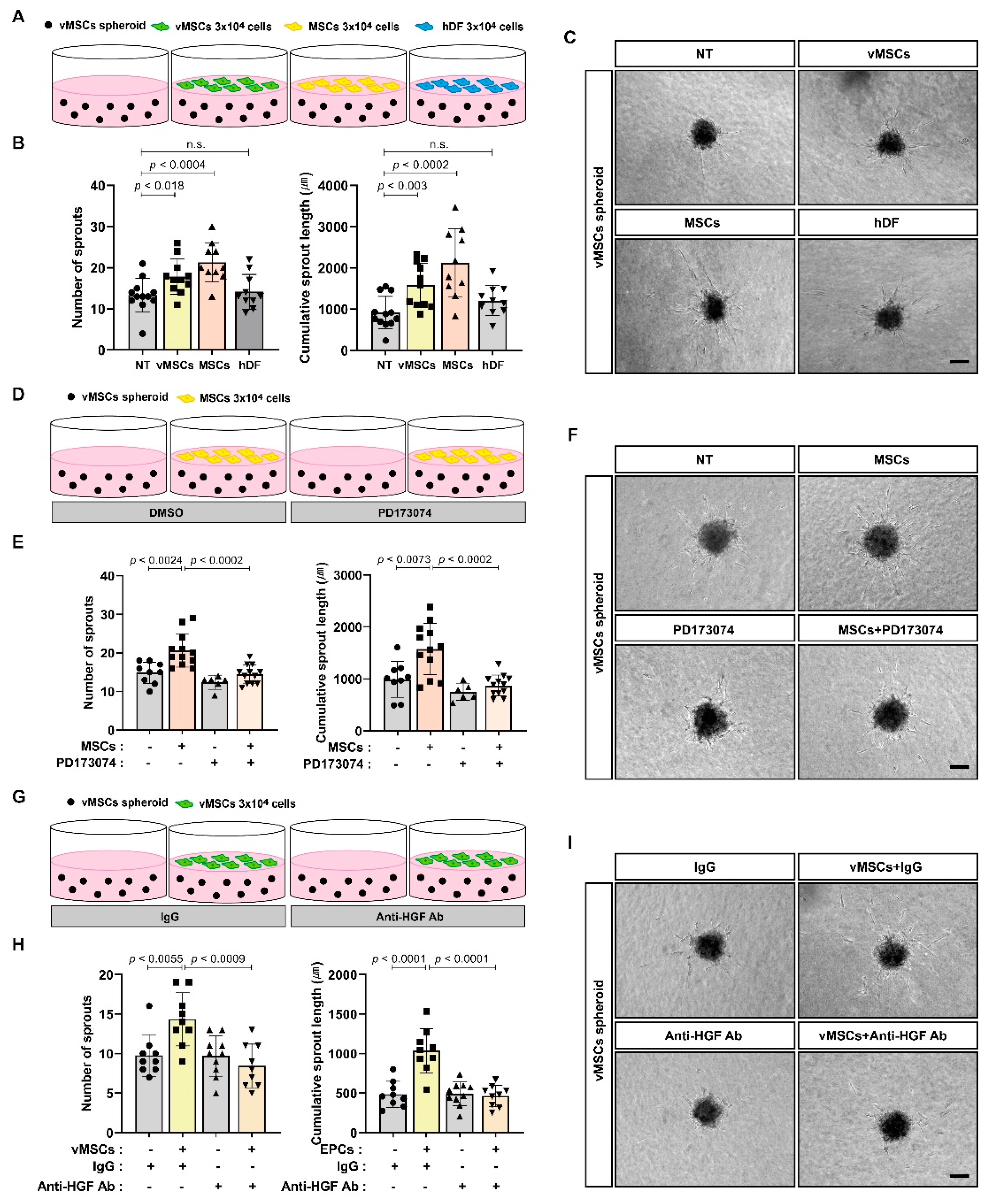
2.2. Increase in Angiogenic Sprouting Capacity of vMSCs in Response to Paracrine Activity of vMSCs and MSCs
2.3. Increase in Angiogenic Sprouting Capacity of HUVECs in Response to vMSCs-Derived HGF
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Real-Time Polymerase Chain Reaction (RT-PCR)
4.3. In Vitro Spheroid Sprouting Assay
4.4. Western Blot Analysis
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asahara, T.; Murohara, T.; Sullivan, A.; Silver, M.; van der Zee, R.; Li, T.; Witzenbichler, B.; Schatteman, G.; Isner, J.M. Isolation of putative progenitor endothelial cells for angiogenesis. Science 1997, 275, 964–967. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Liu, X.; Ding, H.; Zhang, W. Paracrine mechanisms of endothelial progenitor cells in vascular repair. Acta Histochem. 2022, 124, 151833. [Google Scholar] [CrossRef] [PubMed]
- Hur, J.; Yoon, C.H.; Kim, H.S.; Choi, J.H.; Kang, H.J.; Hwang, K.K.; Oh, B.H.; Lee, M.M.; Park, Y.B. Characterization of two types of endothelial progenitor cells and their different contributions to neovasculogenesis. Arter. Thromb. Vasc. Biol. 2004, 24, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Rafii, S.; Wu, M.H.; Wijelath, E.S.; Yu, C.; Ishida, A.; Fujita, Y.; Kothari, S.; Mohle, R.; Sauvage, L.R.; et al. Evidence for circulating bone marrow-derived endothelial cells. Blood 1998, 92, 362–367. [Google Scholar] [CrossRef]
- Medina, R.J.; O’Neill, C.L.; O’Doherty, T.M.; Wilson, S.E.; Stitt, A.W. Endothelial progenitors as tools to study vascular disease. Stem. Cells Int. 2012, 2012, 346735. [Google Scholar] [CrossRef]
- Lin, Y.; Weisdorf, D.J.; Solovey, A.; Hebbel, R.P. Origins of circulating endothelial cells and endothelial outgrowth from blood. J. Clin. Investig. 2000, 105, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Brandl, A.; Yuan, Q.; Boos, A.M.; Beier, J.P.; Arkudas, A.; Kneser, U.; Horch, R.E.; Bleiziffer, O. A novel early precursor cell population from rat bone marrow promotes angiogenesis in vitro. BMC Cell Biol. 2014, 15, 12. [Google Scholar] [CrossRef] [Green Version]
- Reyes, M.; Dudek, A.; Jahagirdar, B.; Koodie, L.; Marker, P.H.; Verfaillie, C.M. Origin of endothelial progenitors in human postnatal bone marrow. J. Clin. Investig. 2002, 109, 337–346. [Google Scholar] [CrossRef]
- Sekiguchi, H.; Ii, M.; Jujo, K.; Yokoyama, A.; Hagiwara, N.; Asahara, T. Improved culture-based isolation of differentiating endothelial progenitor cells from mouse bone marrow mononuclear cells. PLoS ONE 2011, 6, e28639. [Google Scholar] [CrossRef] [Green Version]
- Krenning, G.; van Luyn, M.J.; Harmsen, M.C. Endothelial progenitor cell-based neovascularization: Implications for therapy. Trends Mol. Med. 2009, 15, 180–189. [Google Scholar] [CrossRef]
- Murohara, T.; Ikeda, H.; Duan, J.; Shintani, S.; Sasaki, K.; Eguchi, H.; Onitsuka, I.; Matsui, K.; Imaizumi, T. Transplanted cord blood-derived endothelial precursor cells augment postnatal neovascularization. J. Clin. Investig. 2000, 105, 1527–1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalka, C.; Masuda, H.; Takahashi, T.; Kalka-Moll, W.M.; Silver, M.; Kearney, M.; Li, T.; Isner, J.M.; Asahara, T. Transplantation of ex vivo expanded endothelial progenitor cells for therapeutic neovascularization. Proc. Natl. Acad. Sci. USA 2000, 97, 3422–3427. [Google Scholar] [CrossRef] [PubMed]
- Heil, M.; Ziegelhoeffer, T.; Mees, B.; Schaper, W. A different outlook on the role of bone marrow stem cells in vascular growth: Bone marrow delivers software not hardware. Circ. Res. 2004, 94, 573–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinnaird, T.; Stabile, E.; Burnett, M.S.; Lee, C.W.; Barr, S.; Fuchs, S.; Epstein, S.E. Marrow-derived stromal cells express genes encoding a broad spectrum of arteriogenic cytokines and promote in vitro and in vivo arteriogenesis through paracrine mechanisms. Circ. Res. 2004, 94, 678–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamari, T.; Kawar-Jaraisy, R.; Doppelt, O.; Giladi, B.; Sabbah, N.; Zigdon-Giladi, H. The Paracrine Role of Endothelial Cells in Bone Formation via CXCR4/SDF-1 Pathway. Cells 2020, 9, 1325. [Google Scholar] [CrossRef]
- Yang, Z.; von Ballmoos, M.W.; Faessler, D.; Voelzmann, J.; Ortmann, J.; Diehm, N.; Kalka-Moll, W.; Baumgartner, I.; Di Santo, S.; Kalka, C. Paracrine factors secreted by endothelial progenitor cells prevent oxidative stress-induced apoptosis of mature endothelial cells. Atherosclerosis 2010, 211, 103–109. [Google Scholar] [CrossRef]
- Oswald, J.; Boxberger, S.; Jorgensen, B.; Feldmann, S.; Ehninger, G.; Bornhauser, M.; Werner, C. Mesenchymal stem cells can be differentiated into endothelial cells in vitro. Stem. Cells 2004, 22, 377–384. [Google Scholar] [CrossRef]
- Crisan, M.; Yap, S.; Casteilla, L.; Chen, C.W.; Corselli, M.; Park, T.S.; Andriolo, G.; Sun, B.; Zheng, B.; Zhang, L.; et al. A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem. Cell 2008, 3, 301–313. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Gronthos, S. Perivascular niche of postnatal mesenchymal stem cells in human bone marrow and dental pulp. J. Bone Min. Res. 2003, 18, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Du, W.J.; Chi, Y.; Yang, Z.X.; Li, Z.J.; Cui, J.J.; Song, B.Q.; Li, X.; Yang, S.G.; Han, Z.B.; Han, Z.C. Heterogeneity of proangiogenic features in mesenchymal stem cells derived from bone marrow, adipose tissue, umbilical cord, and placenta. Stem. Cell Res. 2016, 7, 163. [Google Scholar] [CrossRef]
- Zhang, H.; Xian, L.; Lin, Z.; Yang, C.; Zhang, M.; Feng, W.; Peng, X.; Chen, X.; Wu, X. Endothelial progenitor cells as a possible component of stem cell niche to promote self-renewal of mesenchymal stem cells. Mol. Cell. Biochem. 2014, 397, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Peng, X.; Wang, J.; Zhang, H.; Xia, J.; Ge, Q.; Wang, X.; Chen, X.; Wu, X. Mesenchymal stem cells promote endothelial progenitor cell proliferation by secreting insulinlike growth factor1. Mol. Med. Rep. 2017, 16, 1502–1508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Wei, J.; Da Fonseca Ferreira, A.; Wang, H.; Zhang, L.; Zhang, Q.; Bellio, M.A.; Chu, X.M.; Khan, A.; Jayaweera, D.; et al. Rejuvenation of Senescent Endothelial Progenitor Cells by Extracellular Vesicles Derived from Mesenchymal Stromal Cells. JACC Basic Transl. Sci. 2020, 5, 1127–1141. [Google Scholar] [CrossRef]
- Ge, Q.; Zhang, H.; Hou, J.; Wan, L.; Cheng, W.; Wang, X.; Dong, D.; Chen, C.; Xia, J.; Guo, J.; et al. VEGF secreted by mesenchymal stem cells mediates the differentiation of endothelial progenitor cells into endothelial cells via paracrine mechanisms. Mol. Med. Rep. 2018, 17, 1667–1675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fathi, E.; Azarbad, S.; Farahzadi, R.; Javanmardi, S.; Vietor, I. Effect of Rat Bone Marrow Derived-Mesenchymal Stem Cells on Granulocyte Differentiation of Mononuclear Cells as Preclinical Agent in Cellbased Therapy. Curr. Gene 2022, 22, 152–161. [Google Scholar] [CrossRef]
- Lv, F.J.; Tuan, R.S.; Cheung, K.M.; Leung, V.Y. Concise review: The surface markers and identity of human mesenchymal stem cells. Stem. Cells 2014, 32, 1408–1419. [Google Scholar] [CrossRef]
- Matsumoto, K.; Nakamura, T. Emerging multipotent aspects of hepatocyte growth factor. J. Biochem. 1996, 119, 591–600. [Google Scholar] [CrossRef]
- Folkman, J.; Shing, Y. Angiogenesis. J. Biol. Chem. 1992, 267, 10931–10934. [Google Scholar] [CrossRef]
- Ornitz, D.M.; Itoh, N. The Fibroblast Growth Factor signaling pathway. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 215–266. [Google Scholar] [CrossRef] [Green Version]
- Mohammadi, M.; Froum, S.; Hamby, J.M.; Schroeder, M.C.; Panek, R.L.; Lu, G.H.; Eliseenkova, A.V.; Green, D.; Schlessinger, J.; Hubbard, S.R. Crystal structure of an angiogenesis inhibitor bound to the FGF receptor tyrosine kinase domain. EMBO J. 1998, 17, 5896–5904. [Google Scholar] [CrossRef]
- Schroder, K.; Schutz, S.; Schloffel, I.; Batz, S.; Takac, I.; Weissmann, N.; Michaelis, U.R.; Koyanagi, M.; Brandes, R.P. Hepatocyte growth factor induces a proangiogenic phenotype and mobilizes endothelial progenitor cells by activating Nox2. Antioxid. Redox Signal 2011, 15, 915–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, X.; Yang, S.; Ingle, G.; Zlot, C.; Rangell, L.; Kowalski, J.; Schwall, R.; Ferrara, N.; Gerritsen, M.E. Hepatocyte growth factor enhances vascular endothelial growth factor-induced angiogenesis in vitro and in vivo. Am. J. Pathol. 2001, 158, 1111–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prockop, D.J. Marrow stromal cells as stem cells for nonhematopoietic tissues. Science 1997, 276, 71–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Yang, J.; Fang, J.; Zhou, Y.; Candi, E.; Wang, J.; Hua, D.; Shao, C.; Shi, Y. The secretion profile of mesenchymal stem cells and potential applications in treating human diseases. Signal Transduct. Target 2022, 7, 92. [Google Scholar] [CrossRef]
- Shi, W.; Xin, Q.; Yuan, R.; Yuan, Y.; Cong, W.; Chen, K. Neovascularization: The Main Mechanism of MSCs in Ischemic Heart Disease Therapy. Front. Cardiovasc. Med. 2021, 8, 633300. [Google Scholar] [CrossRef]
- Abedin, M.; Tintut, Y.; Demer, L.L. Mesenchymal stem cells and the artery wall. Circ. Res. 2004, 95, 671–676. [Google Scholar] [CrossRef] [Green Version]
- Schmelzer, E.; McKeel, D.T.; Gerlach, J.C. Characterization of Human Mesenchymal Stem Cells from Different Tissues and Their Membrane Encasement for Prospective Transplantation Therapies. Biomed Res. Int. 2019, 2019, 6376271. [Google Scholar] [CrossRef]
- Clavreul, A.; Pourbaghi-Masouleh, M.; Roger, E.; Lautram, N.; Montero-Menei, C.N.; Menei, P. Human mesenchymal stromal cells as cellular drug-delivery vectors for glioblastoma therapy: A good deal? J. Exp. Clin. Cancer Res. 2017, 36, 135. [Google Scholar] [CrossRef] [Green Version]
- Park, K.S.; Schecterson, L.; Gumbiner, B.M. Enhanced endothelial barrier function by monoclonal antibody activation of vascular endothelial cadherin. Am. J. Physiol. Heart Circ. Physiol. 2021, 320, H1403–H1410. [Google Scholar] [CrossRef]
- Fathi, E.; Vandghanooni, S.; Montazersaheb, S.; Farahzadi, R. Mesenchymal stem cells promote caspase-3 expression of SH-SY5Y neuroblastoma cells via reducing telomerase activity and telomere length. Iran. J. Basic Med. Sci. 2021, 24, 1583–1589. [Google Scholar] [CrossRef]
- Kim, W.; Park, A.; Jang, H.H.; Kim, S.E.; Park, K.S. Breast Tumor Cell-Stimulated Bone Marrow-Derived Mesenchymal Stem Cells Promote the Sprouting Capacity of Endothelial Cells by Promoting VEGF Expression, Mediated in Part through HIF-1alpha Increase. Cancers 2022, 14, 4711. [Google Scholar] [CrossRef] [PubMed]
- Dubon, M.J.; Yu, J.; Choi, S.; Park, K.S. Transforming growth factor beta induces bone marrow mesenchymal stem cell migration via noncanonical signals and N-cadherin. J. Cell. Physiol. 2018, 233, 201–213. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jang, H.H.; Son, Y.; Park, G.; Park, K.-S. Bone Marrow-Derived Vasculogenic Mesenchymal Stem Cells Enhance In Vitro Angiogenic Sprouting of Human Umbilical Vein Endothelial Cells. Int. J. Mol. Sci. 2023, 24, 413. https://doi.org/10.3390/ijms24010413
Jang HH, Son Y, Park G, Park K-S. Bone Marrow-Derived Vasculogenic Mesenchymal Stem Cells Enhance In Vitro Angiogenic Sprouting of Human Umbilical Vein Endothelial Cells. International Journal of Molecular Sciences. 2023; 24(1):413. https://doi.org/10.3390/ijms24010413
Chicago/Turabian StyleJang, Hyun Hee, Youngsook Son, Gabee Park, and Ki-Sook Park. 2023. "Bone Marrow-Derived Vasculogenic Mesenchymal Stem Cells Enhance In Vitro Angiogenic Sprouting of Human Umbilical Vein Endothelial Cells" International Journal of Molecular Sciences 24, no. 1: 413. https://doi.org/10.3390/ijms24010413