
Phylogenetic, Structural and Functional Evolution of the LHC Gene Family in Plant Species
Abstract
:1. Introduction
2. Results
2.1. Identification and Distribution of LHC Genes in Plants
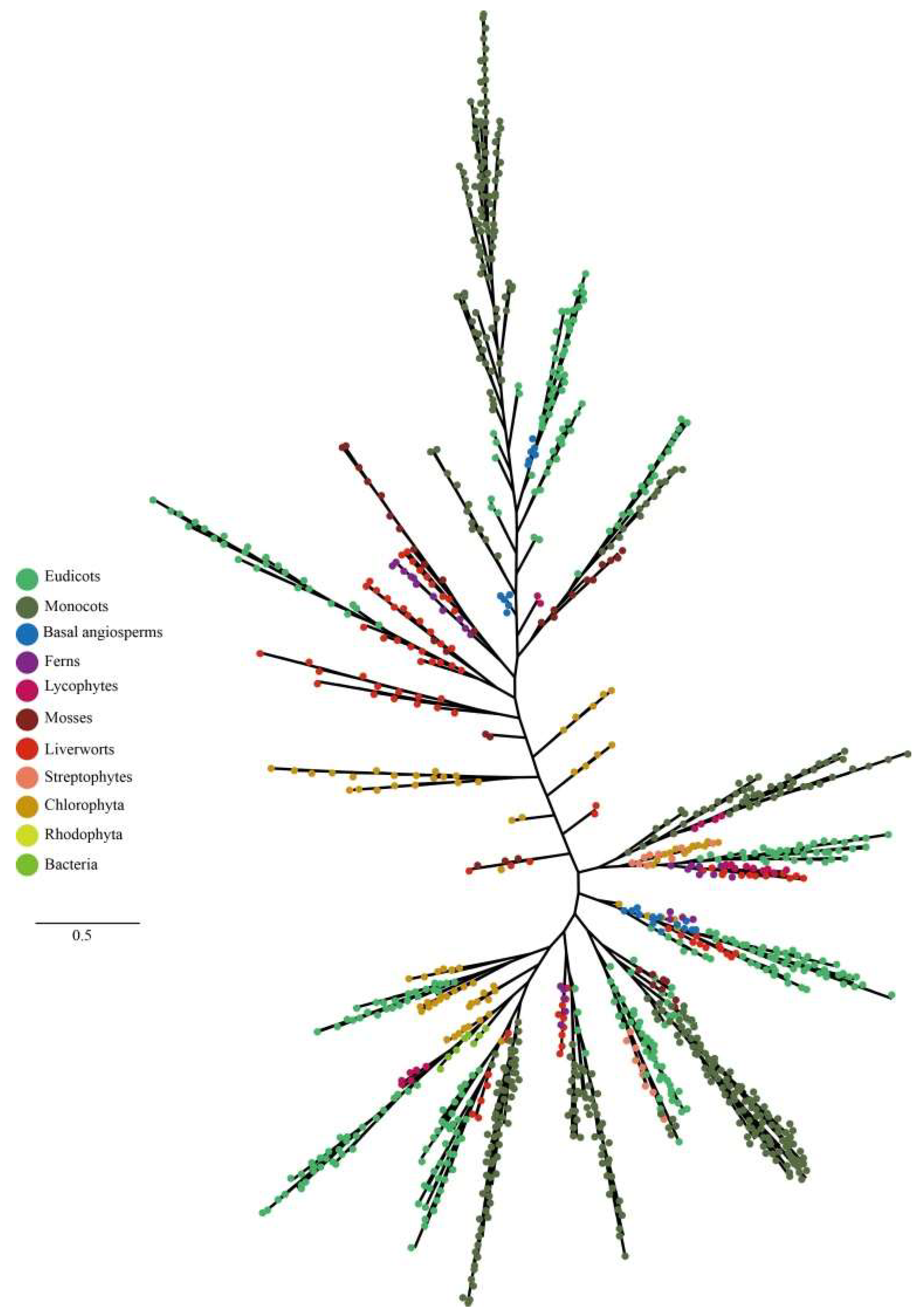
2.2. Phylogenetic Analysis and Conserved Motifs of the Plant LHC Gene Family Members
2.3. Expansion Pattern of LHC Genes and Collinearity Analysis
2.4. Expression Patterns of LHCs in Plants
3. Discussion
4. Methods and Materials
4.1. Identification of LHC Gene Family Members
4.2. Phylogeny Analysis and Identification of Conserved Motifs
4.3. Microarray Based Expression Analysis
4.4. Gene Duplication and Synteny Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiang, Q.; Xu, Z.S.; Wang, F.; Li, M.Y.; Ma, J.; Xiong, A.S. Effects of abiotic stresses on the expression of Lhcb1 gene and photosynthesis of Oenanthe javanica and Apium graveolens. Biol. Plantarum. 2014, 58, 256–264. [Google Scholar] [CrossRef]
- Klimmek, F.; Sjödin, A.; Noutsos, C.; Leister, D.; Jansson, S. Abundantly and rarely expressed Lhc protein genes exhibit distinct regulation patterns in plants. Plant Physiol. 2006, 140, 793–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Z.; Yang, J. Genomics analysis of the light-harvesting chlorophyll a/b-binding (Lhc) superfamily in cassava (Manihot esculenta Crantz). Gene 2019, 702, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Jansson, S. A guide to the Lhc genes and their relatives in Arabidopsis/IT>. Trends Plant Sci. 1999, 4, 236–240. [Google Scholar] [CrossRef]
- Green, B.R.; Pichersky, E.; Kloppstech, K. Chlorophyll a/b-binding proteins: An extended family. Trends Biochem. Sci. 1991, 16, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Engelken, J.; Brinkmann, H.; Adamska, I. Taxonomic distribution and origins of the extended LHC (light-harvesting complex) antenna protein superfamily. BMC Evol. Biol. 2010, 10, 233. [Google Scholar] [CrossRef] [Green Version]
- Zou, Z. Mining gene families in the castor bean genome. In The Castor Bean Genome; Rabinowicz, P., Kole, C., Eds.; Springer: Cham, Switzerland, 2018; pp. 135–173. [Google Scholar]
- Zhao, Y.; Kong, H.; Guo, Y.; Zou, Z. Light-harvesting chlorophyll a/b-binding protein-coding genes in jatropha and the comparison with castor, cassava and Arabidopsis. PeerJ. 2020, 8, e8465. [Google Scholar] [CrossRef]
- Neilson, A.; Durnford, G. Structural and functional diversification of the light-harvesting complexes in photosynthetic eukaryotes. Photosynth Res. 2010, 106, 57–71. [Google Scholar] [CrossRef]
- Mogen, K.; Eide, J.; Duysen, M.; Eskins, K. Chloramphenicol Stimulates the Accumulation of Light-Harvesting Chlorophyll a/b Protein II by Affecting Posttranscriptional Events in the Chlorina CD3 Mutant Wheat. Plant Physiol. 1990, 92, 1233–1240. [Google Scholar] [CrossRef] [Green Version]
- Passarini, F.; Xu, P.; Caffarri, S.; Hille, J.; Croce, R. Towards in vivo mutation analysis: Knock-out of specific chlorophylls bound to the light-harvesting complexes of Arabidopsis thaliana-the case of CP24 (Lhcb6). Biochim. Biophys. Acta 2014, 1837, 1500–1506. [Google Scholar] [CrossRef]
- Xu, H.; Liu, R.; Yan, L.; Liu, Q.; Jiang, C.; Shen, Y.; Wang, F.; Zhang, P. Light-harvesting chlorophyll a/b-binding proteins are required for stomatal response to abscisic acid in Arabidopsis. J. Exp. Bot. 2012, 63, 1095–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.-S.; Song, Z.-B.; Sun, Z.; Zhang, J.; Mei, Y.; Nian, H.-J.; Li, K.-Z.; Chen, L.-M. Effects of Formaldehyde Stress on Physiological Characteristics and Gene Expression Associated with Photosynthesis in Arabidopsis thaliana. Plant Mol. Biol. 2012, 30, 1291–1302. [Google Scholar] [CrossRef]
- Peterson, R.B.; Schultes, N.P. Light-harvesting complex B7 shifts the irradiance response of photosynthetic light-harvesting regulation in leaves of Arabidopsis thaliana. J. Plant Physiol. 2014, 171, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, J.; Yu, Y.; Kou, X.; Periakaruppan, R.; Chen, X.; Li, X. STAY-GREEN and light-harvesting complex II chlorophyll a/b binding protein are involved in albinism of a novel albino tea germplasm ‘Huabai 1′. Sci. Hortic. 2022, 293, 110653. [Google Scholar] [CrossRef]
- De Bianchi, S.; Betterle, N.; Kouril, R.; Cazzaniga, S.; Boekema, E.; Bassi, R.; Dall’Osto, L. Arabidopsis mutants deleted in the light-harvesting protein Lhcb4 have a disrupted photosystem II macrostructure and are defective in photoprotection. Plant Cell 2011, 23, 2659–2679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Abid, M.; Tu, J.; Gao, P.; Wang, Z.; Huang, H. Genome-Wide Identification of the LHC Gene Family in Kiwifruit and Regulatory Role of AcLhcb3.1/3.2 for Chlorophyll a Content. Int. J. Mol. Sci. 2022, 23, 6528. [Google Scholar] [CrossRef] [PubMed]
- Castelletti, S.; Morosinotto, T.; Robert, B.; Caffarri, S.; Bassi, R.; Croce, R. Recombinant Lhca2 and Lhca3 subunits of the pho-tosystem I antenna system. Biochemistry 2003, 42, 4226–4234. [Google Scholar] [CrossRef]
- Deng, S.; Kong, Y.; Zhou, B.; Zhang, S.; Yue, M.; Meng, W. Heterology expression of the tomato LeLhcb2 gene confers elevated tolerance to chilling stress in transgenic tobacco. Plant Physiol. Biochem. 2014, 80, 318–327. [Google Scholar] [CrossRef]
- Zhang, Q.; Ma, C.; Wang, X.; Ma, Q.; Fan, S.; Zhang, C. Genome-wide identification of the light-harvesting chlorophyll a/b binding (Lhc) family in Gossypium hirsutum reveals the influence of GhLhcb2.3 on chlorophyll a synthesis. Plant Biol. 2021, 23, 831–842. [Google Scholar] [CrossRef]
- Umate, P. Genome-wide analysis of the family of light-harvesting chlorophyll a/b-binding proteins in Arabidopsis and rice. Plant Signal. Behav. 2010, 5, 1537–1542. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST plus: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Grimm, B. Connecting Chlorophyll Metabolism with Accumulation of the Photosynthetic Apparatus. Trends Plant Sci. 2021, 26, 484–495. [Google Scholar] [CrossRef] [PubMed]
- De Bianchi, S.; Dall’Osto, L.; Tognon, G.; Morosinotto, T.; Bassi, R. Minor antenna proteins CP24 and CP26 affect the interactions between photosystem II subunits and the electron transport rate in grana membranes of Arabidopsis. Plant Cell. 2008, 20, 1012–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.E.; Ma, J.; Wu, N.; Su, Y.Q.; Zhang, Z.W.; Yuan, M.; Zhang, H.Y.; Zeng, X.Y.; Yuan, S. The roles of Arabidopsis proteins of Lhcb4, Lhcb5 and Lhcb6 in oxidative stress under natural light conditions. Plant Physiol. Biochem. 2018, 130, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Li, M.; Jia, R.; Zhao, H.; He, P.; Zhang, Y.; Guo, A. Genes encoding light-harvesting chlorophyll a/b-binding proteins in papaya (Carica papaya L.) and insight into lineage-specific evolution in Brassicaceae. Gene 2020, 748, 144685. [Google Scholar] [CrossRef]
- Pan, Z.; Chen, L.; Wang, F.; Song, W.; Cao, A.; Xie, S.; Chen, X.; Jin, X.; Li, H. Genome-wide identification and expression analysis of the ascorbate oxidase gene family in Gossypium hirsutum reveals the critical role of GhAO1A in delaying dark-induced leaf senescence. Int. J. Mol. Sci. 2019, 20, 6167. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, H.; Wang, J.; Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.H.; Bancroft, I.; Cheng, F.; et al. The Genome of the Mesopolyploid Crop Species Brassica Rapa. Nat. Genet. 2011, 43, 1035–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, F.; Mandáková, T.; Wu, J.; Xie, Q.; Lysak, M.A.; Wang, X. Deciphering the Diploid Ancestral Genome of the Mesohexaploid Brassica Rapa. Plant Cell. 2013, 25, 1541–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Ma, B.; Shen, J.; Zhao, S.; Ma, X.; Wang, Z.; Fan, Y.; Tang, Q.; Wei, D. The Evolution of the Expansin Gene Family in Brassica Species. Plant Physiol. Biochem. 2021, 167, 630–638. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, S.; He, F.; Zhu, J.; Hu, S.; Yu, J. How do variable substitution rates influence Ka and Ks calculations? Genom. Proteom. Bioinform. 2009, 7, 116–127. [Google Scholar] [CrossRef]
- Yu, F.; Tang, C.Q.; Xin, Y.Y.; Peng, D.C.; Xu, Y.N.; Li, L.B.; Kuang, T.Y. Progress in the structural and functional study of photosystem I. Chin. Bull. Bot. 2001, 18, 266–275. [Google Scholar]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bateman, A.; Coin, L.; Durbin, R.; Finn, R.D.; Hollich, V.; Grifths-Jones, S.; Khanna, A.; Marshall, M.; Moxon, S.; Sonnhammer, E.L.L.; et al. The Pfam protein families database. Nucleic Acids Res. 2004, 32 (Suppl. S1), D138–D141. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Doerks, T.; Bork, P. SMART: Recent updates, new developments and status in 2015. Nucleic Acids Res. 2015, 43, D257–D260. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2002, 32, 1792-7. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Feng, K.; Wang, M.; Wang, M.; Deng, P.; Song, W.; Nie, X. Genome-wide identifcation, phylogeny and expression analysis of AP2/ERF tran scription factors family in Brachypodium distachyon. BMC Genom. 2016, 17, 636. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Conery, J.S. The evolutionary fate and consequences of duplicate genes. Science 2000, 290, 1151–1155. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Species | Total | Lhca/b | Lil | PsbS | FCII | |||
|---|---|---|---|---|---|---|---|---|---|
| OHP | SEP | ELIP | Psb33 | ||||||
| Eudictos | Arabidopsis thaliana | 32 | 22 | 2 | 2 | 2 | 1 | 1 | 2 |
| Brassica rapa | 44 | 29 | 4 | 3 | 4 | 0 | 2 | 2 | |
| Gossypium raimondii | 44 | 24 | 5 | 2 | 3 | 1 | 2 | 7 | |
| Theobroma cacao | 25 | 15 | 2 | 2 | 1 | 2 | 1 | 2 | |
| Eucalyptus grandis | 30 | 17 | 5 | 3 | 2 | 0 | 1 | 2 | |
| Malus domestica | 42 | 26 | 4 | 2 | 3 | 1 | 2 | 4 | |
| Glycine max | 62 | 34 | 7 | 7 | 3 | 1 | 3 | 7 | |
| Populus trichocarpa | 36 | 22 | 3 | 3 | 3 | 1 | 1 | 3 | |
| Vitis vinifera | 31 | 20 | 2 | 2 | 3 | 1 | 1 | 2 | |
| Solanum lycopersicum | 40 | 24 | 3 | 2 | 4 | 1 | 1 | 5 | |
| Chenopodium quinoa | 49 | 29 | 4 | 4 | 2 | 2 | 2 | 6 | |
| Monocots | Zostera marina | 32 | 22 | 1 | 2 | 2 | 1 | 1 | 3 |
| Phoenix dactylifera | 33 | 21 | 3 | 3 | 2 | 0 | 1 | 3 | |
| Triticum aestivum | 58 | 31 | 6 | 3 | 3 | 6 | 3 | 6 | |
| Hordeum vulgare | 30 | 22 | 2 | 1 | 1 | 1 | 1 | 2 | |
| Zea mays | 37 | 25 | 3 | 2 | 2 | 1 | 1 | 3 | |
| Sorghum bicolor | 29 | 17 | 3 | 2 | 1 | 1 | 1 | 4 | |
| Oryza sativa | 25 | 14 | 3 | 1 | 1 | 2 | 2 | 2 | |
| Basal angiosperms | Amborella trichopoda | 27 | 15 | 2 | 3 | 3 | 0 | 1 | 3 |
| Ferns | Adiantum capillusveneris | 42 | 35 | 2 | 0 | 1 | 0 | 1 | 3 |
| Ceratopteris richardii | 54 | 40 | 4 | 1 | 2 | 0 | 2 | 5 | |
| Lycophytes | Selaginella moellendorffii | 25 | 16 | 1 | 0 | 2 | 0 | 2 | 4 |
| Mosses | Sphagnum magellanicum | 53 | 37 | 5 | 0 | 4 | 0 | 4 | 3 |
| Ceratodon purpureus | 41 | 34 | 2 | 0 | 3 | 0 | 1 | 1 | |
| Liverworts | Marchantia polymorpha | 54 | 45 | 2 | 0 | 2 | 0 | 3 | 2 |
| Physcomitrium patens | 51 | 40 | 2 | 0 | 4 | 0 | 1 | 4 | |
| Streptophyta | Marchantia paleacea | 56 | 47 | 2 | 0 | 2 | 1 | 3 | 1 |
| Chara braunii | 16 | 12 | 1 | 0 | 0 | 0 | 2 | 1 | |
| Chlorophyta | Chlamydomonas reinhardtii | 28 | 23 | 1 | 0 | 1 | 0 | 2 | 1 |
| Volvox cartei | 30 | 27 | 0 | 1 | 0 | 0 | 1 | 1 | |
| Ostreococcus sp | 11 | 9 | 0 | 0 | 1 | 0 | 0 | 1 | |
| Coccomyxa subellipsoidea | 22 | 20 | 0 | 0 | 1 | 0 | 0 | 1 | |
| Dunaliella salina | 16 | 13 | 0 | 0 | 1 | 0 | 1 | 1 | |
| Rhodophyta | Cyanidioschyzon merolae | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Ectocarpus siliculosus | 9 | 8 | 0 | 0 | 0 | 0 | 0 | 1 | |
| Bacteria | Bathycoccus prasinos | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Panttoea ananatis | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | |
| Erwinia gerundensis | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | |
| Halomonas sulfidaeris | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | |
| Cronobacter sakazakii | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | |
| Geobacter sulfurreducens | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | |
| Gloeobacter violaceus | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lan, Y.; Song, Y.; Zhao, F.; Cao, Y.; Luo, D.; Qiao, D.; Cao, Y.; Xu, H. Phylogenetic, Structural and Functional Evolution of the LHC Gene Family in Plant Species. Int. J. Mol. Sci. 2023, 24, 488. https://doi.org/10.3390/ijms24010488
Lan Y, Song Y, Zhao F, Cao Y, Luo D, Qiao D, Cao Y, Xu H. Phylogenetic, Structural and Functional Evolution of the LHC Gene Family in Plant Species. International Journal of Molecular Sciences. 2023; 24(1):488. https://doi.org/10.3390/ijms24010488
Chicago/Turabian StyleLan, Yanhong, Yao Song, Fei Zhao, Yu Cao, Dening Luo, Dairong Qiao, Yi Cao, and Hui Xu. 2023. "Phylogenetic, Structural and Functional Evolution of the LHC Gene Family in Plant Species" International Journal of Molecular Sciences 24, no. 1: 488. https://doi.org/10.3390/ijms24010488