
Changes in the Expression and Functional Activities of C-X-C Motif Chemokine Ligand 13 (CXCL13) in Hyperplastic Prostate
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
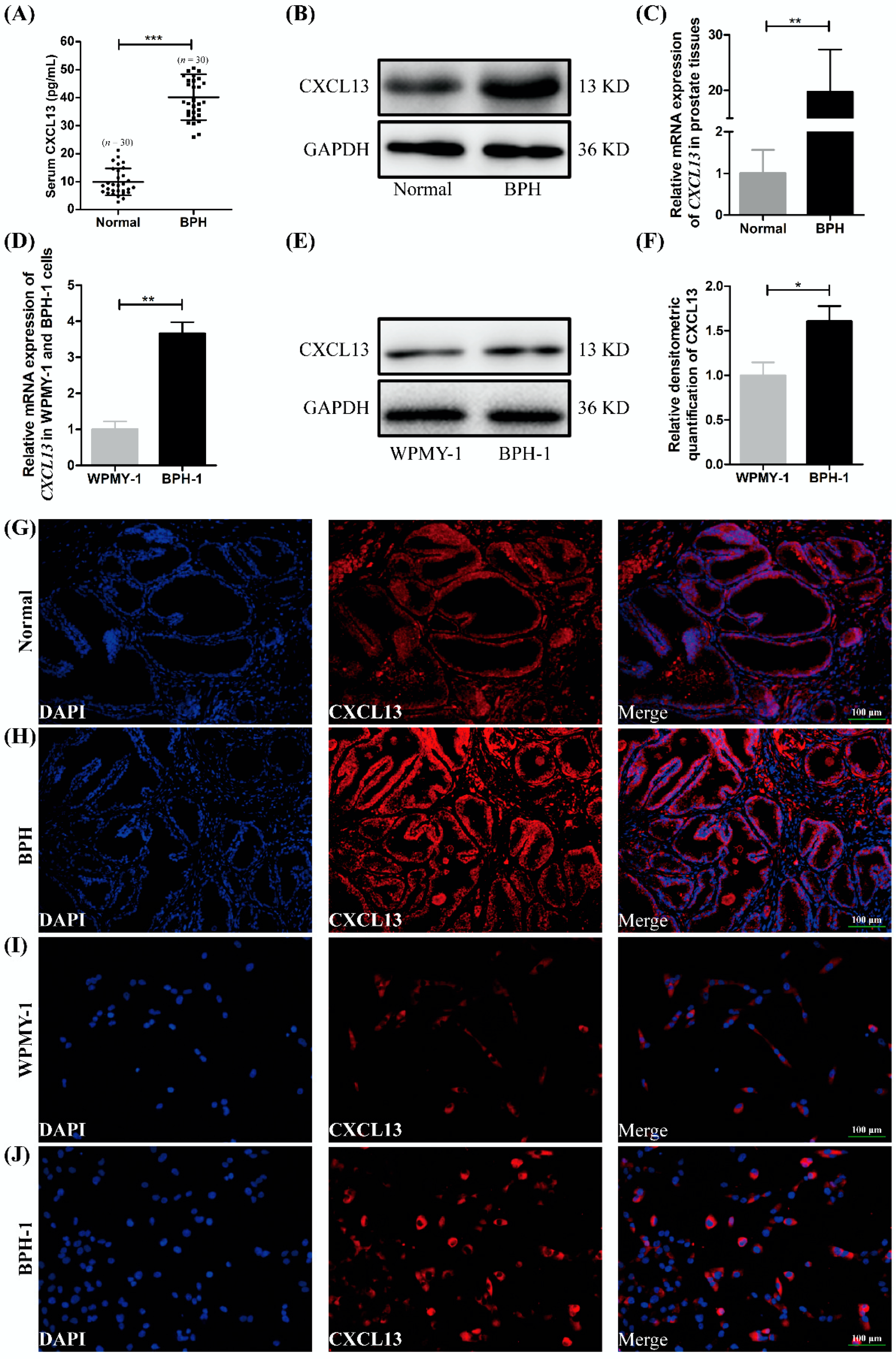
2.1. The Expression and Localization of CXCL13 in Human Prostate Tissues and Cell Lines
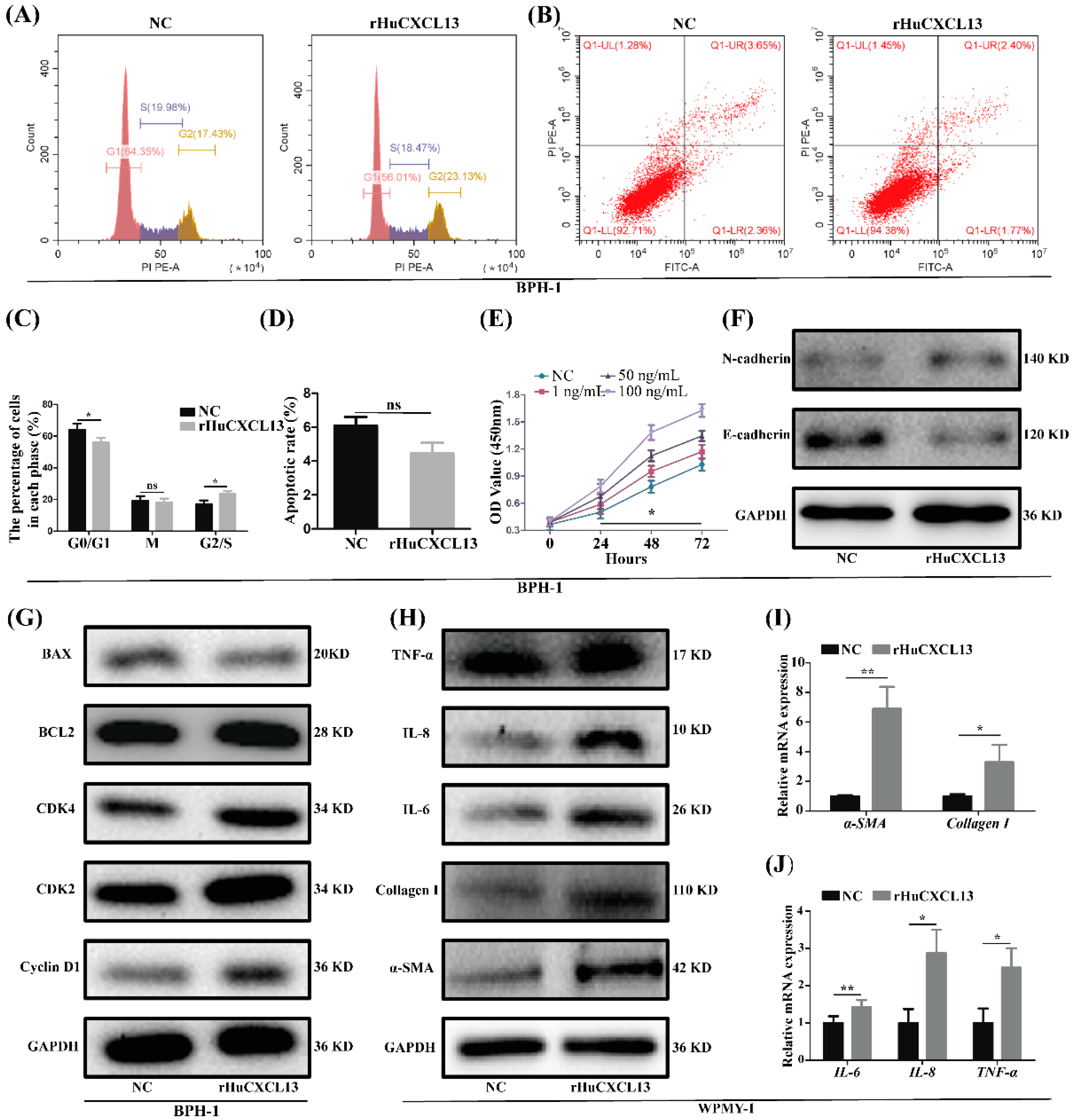
2.2. rHuCXCL13 Treatment Promoted Proliferation and EMT of BPH-1 Cells, and Enhanced Inflammation and Fibrosis of WPMY-1 Cells
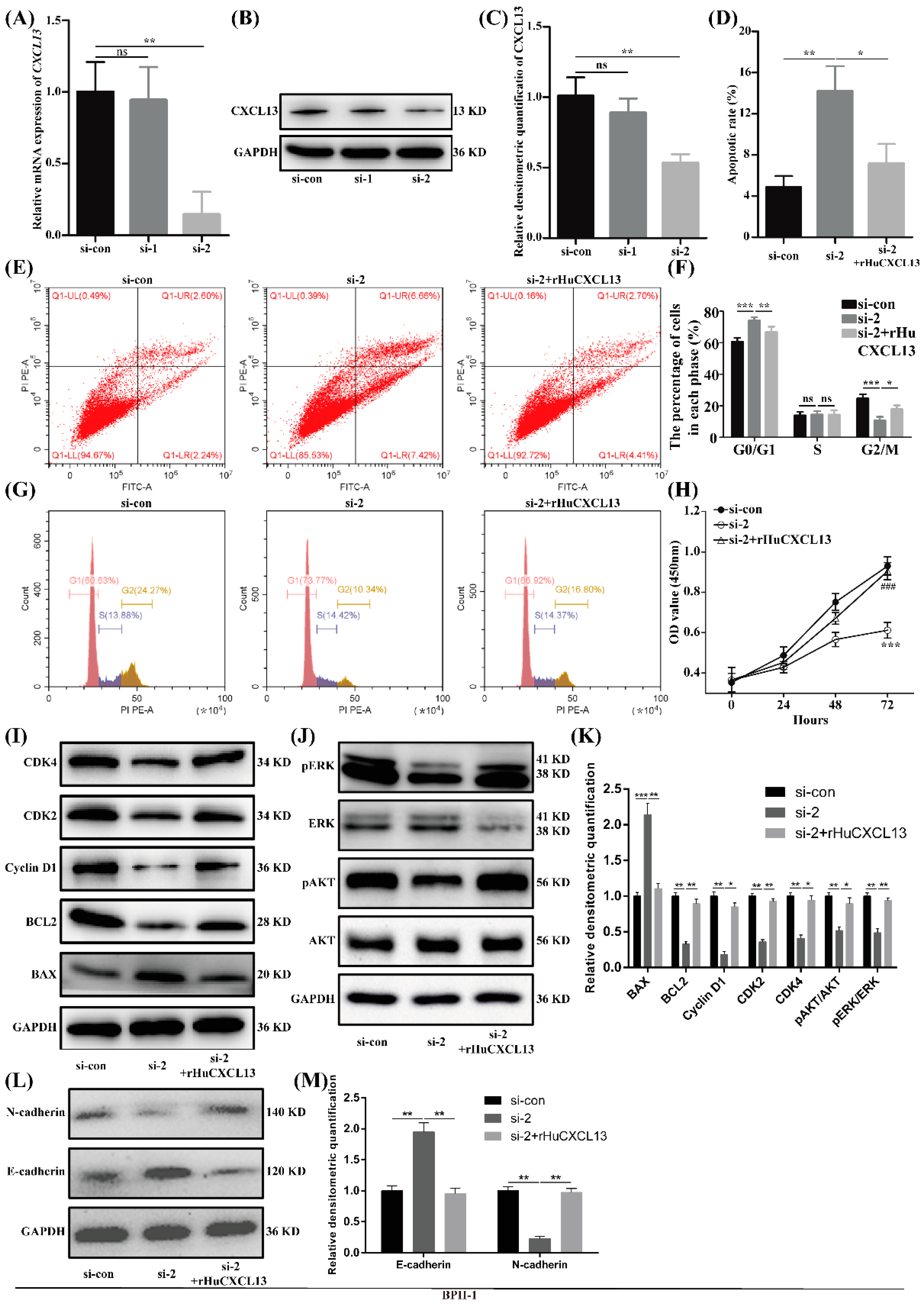
2.3. Knockdown of CXCL13 Inhibited Proliferation, EMT and Promoted Apoptosis in BPH-1 Cells via ERK1/2 and AKT Pathway
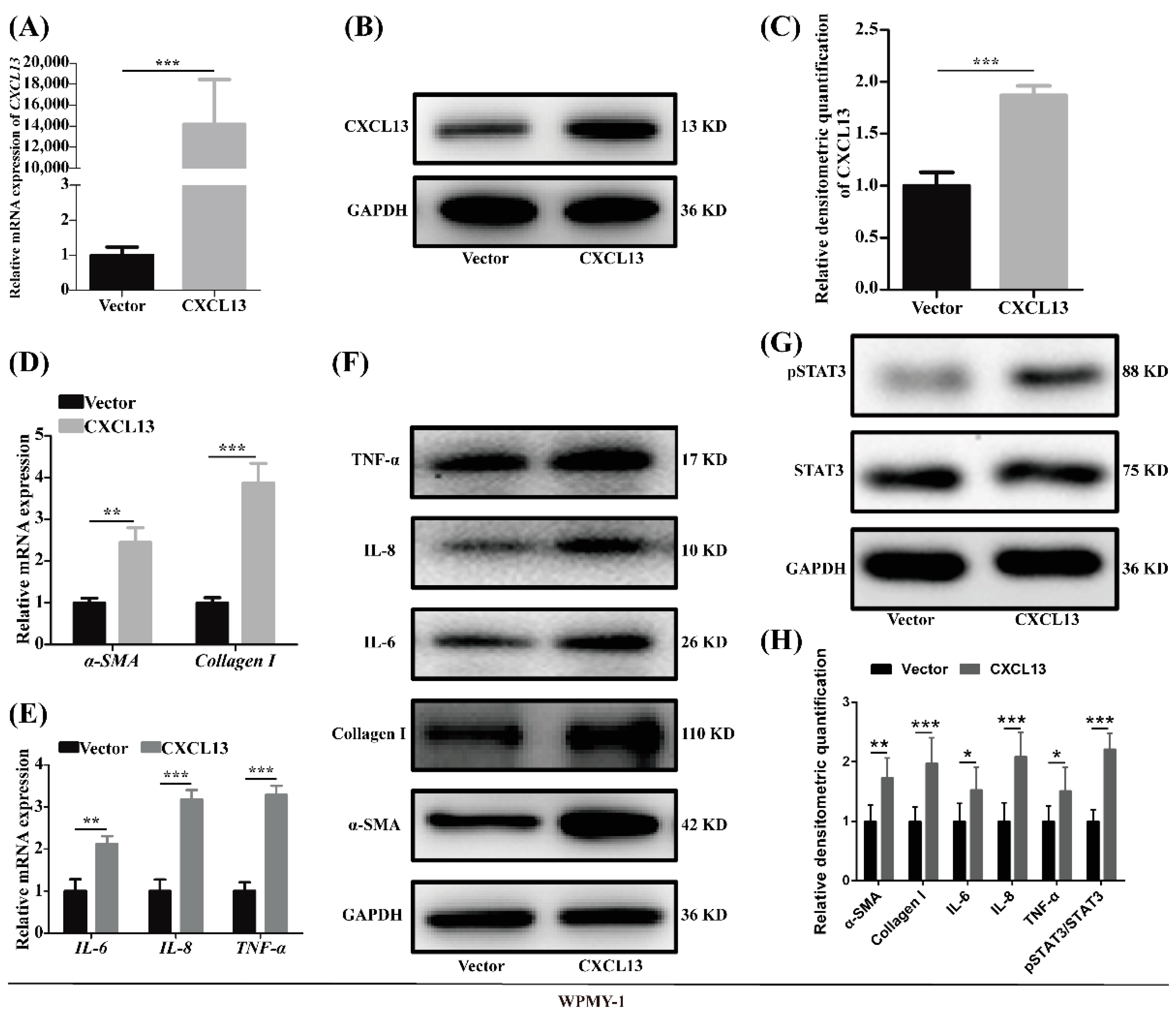
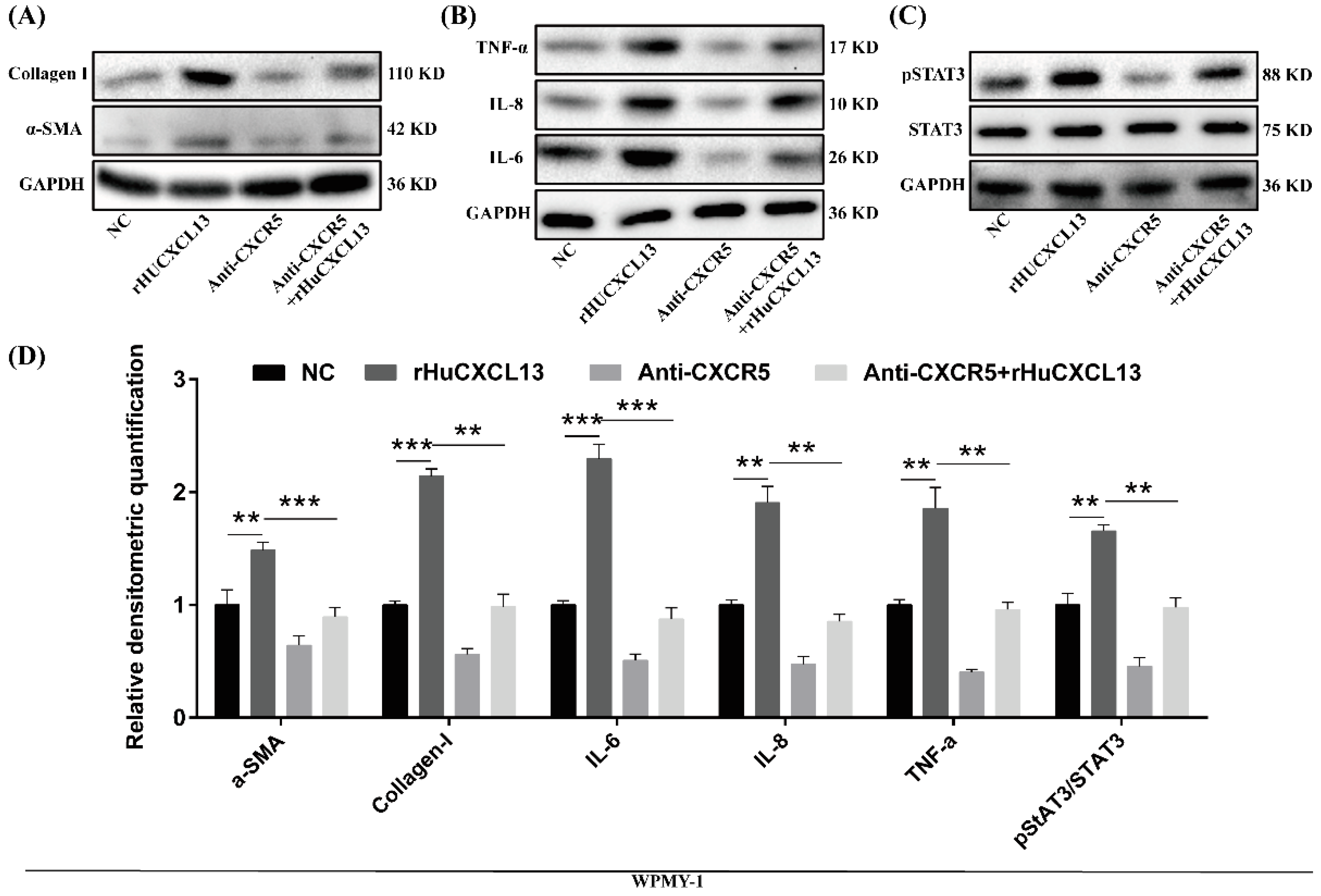
2.4. Overexpression of CXCL13 Upregulated Inflammation and Fibrosis in WPMY-1 Cells through STAT3 Pathway
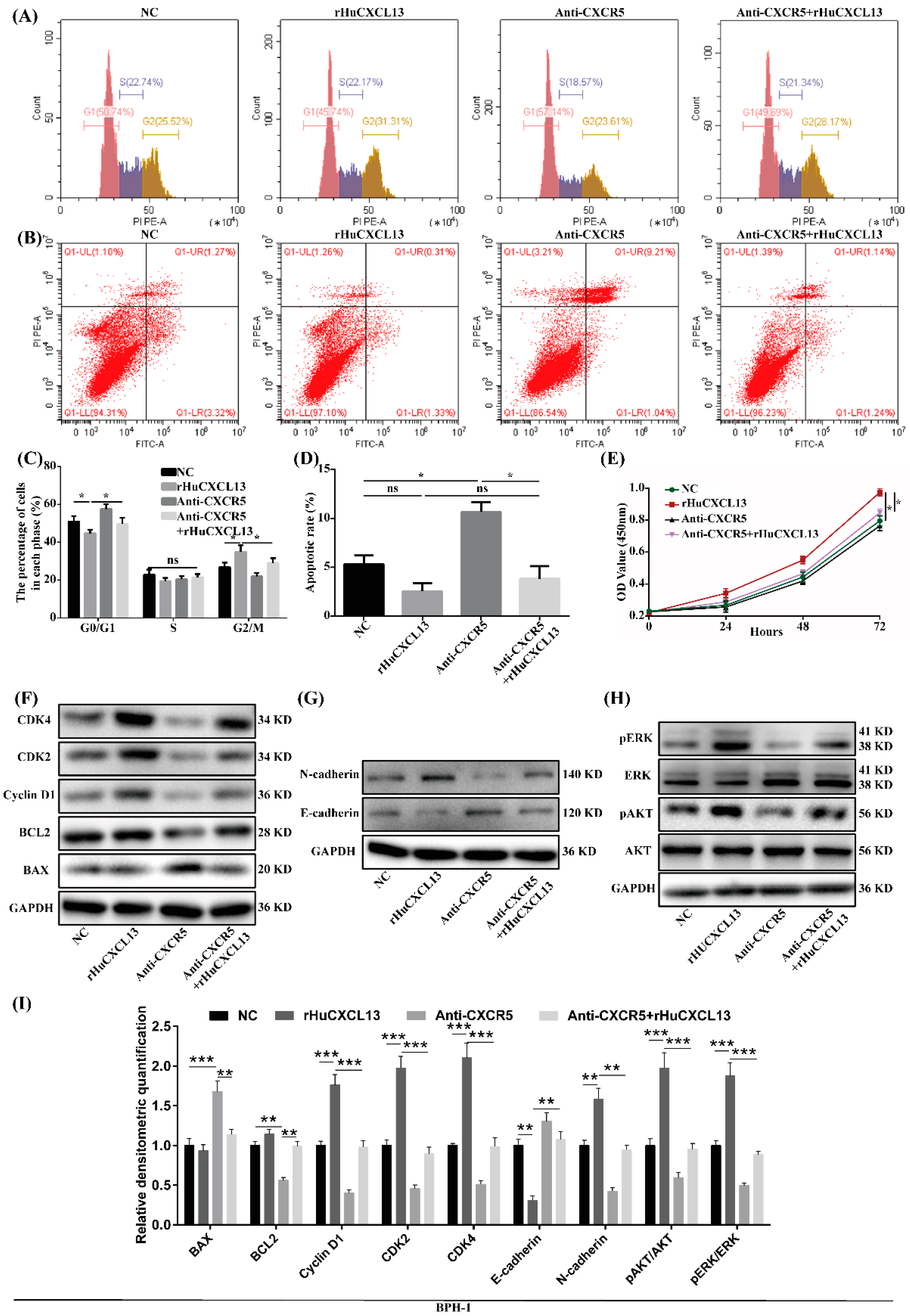
2.5. Anti-CXCR5 Could Rescue the Alternations Induced by rHuCXCL13
2.6. rHuCXCL13 Induces the Development of BPH In Vivo
2.7. TMA Showed the Correlations of CXCL13 with Clinical Parameters
3. Discussion
4. Materials and Methods
4.1. Animals, Tissues and Serum
4.2. TMA Construction and Immunohistochemistry Analysis
4.3. Cell Culture
4.4. Addition of rHuCXCL13 in BPH-1 and WPMY-1 Cells
4.5. Addition of Anti-CXCR5 in BPH-1 and WPMY-1 Cells
4.6. Knockdown of CXCL13 in BPH-1 Cells
4.7. Overexpression of CXCL13 in WPMY-1 Cells
4.8. Total RNA Extraction, RNA Reverse Transcription and qRT-PCR Analysis
4.9. Western Blotting Analysis
4.10. Flow Cytometry Analysis
4.11. Cell Counting Kit-8 (CCK8) Assay
4.12. Immunofluorescent Staining for Tissues
4.13. Immunofluorescent Staining for Cells
4.14. Hematoxylin and Eosin (H&E) Staining
4.15. Masson’s Trichrome Staining
4.16. Enzyme-Linked Immune Sorbent Assay
4.17. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roehrborn, C. Benign prostatic hyperplasia: Etiology, pathophysiology, epidemiology and natural history. In Campbell-Walsh Urology; Partin, A.W., Dmochowski, R.R., Kavoussi, L.R., Partin, A.W., Peters, C.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 15079–15247. [Google Scholar]
- Luo, J.; Dunn, T.; Ewing, C.; Sauvageot, J.; Chen, Y.; Trent, J.; Isaacs, W. Gene expression signature of benign prostatic hyperplasia revealed by cDNA microarray analysis. Prostate 2002, 51, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Middleton, L.W.; Shen, Z.; Varma, S.; Pollack, A.S.; Gong, X.; Zhu, S.; Zhu, C.; Foley, J.W.; Vennam, S.; Sweeney, R.T.; et al. Genomic analysis of benign prostatic hyperplasia implicates cellular re-landscaping in disease pathogenesis. JCI Insight 2019, 5, e129749. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Jiang, Y.; He, W.; Xu, D.; Chen, P.; Liu, D.; Liu, J.; Wang, X.; DiSanto, M.E.; Zhang, X. Identification and functional activity of matrix-remodeling associated 5 (MXRA5) in benign hyperplastic prostate. Aging 2020, 12, 8605–8621. [Google Scholar] [CrossRef] [PubMed]
- Alturaiki, W. Elevated Plasma Levels of CXCL13 Chemokine in Saudi Patients With Asthma Exacerbation. Cureus 2022, 14, e21142. [Google Scholar] [CrossRef] [PubMed]
- Heinrichs, M.; Ashour, D.; Siegel, J.; Büchner, L.; Wedekind, G.; Heinze, K.G.; Arampatzi, P.; Saliba, A.E.; Cochain, C.; Hofmann, U.; et al. The healing myocardium mobilizes a distinct B-cell subset through a CXCL13-CXCR5-dependent mechanism. Cardiovasc. Res. 2021, 117, 2664–2676. [Google Scholar] [CrossRef]
- Zanetti, C.; Kumar, R.; Ender, J.; Godavarthy, P.S.; Hartmann, M.; Hey, J.; Breuer, K.; Weissenberger, E.S.; Minciacchi, V.R.; Karantanou, C.; et al. The age of the bone marrow microenvironment influences B-cell acute lymphoblastic leukemia progression via CXCR5-CXCL13. Blood 2021, 138, 1870–1884. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, T.; Miyagawa, T.; Toyama, S.; Yamashita, T.; Nakamura, K.; Saigusa, R.; Ichimura, Y.; Takahashi, T.; Toyama, T.; Yoshizaki, A.; et al. CXCL13 produced by macrophages due to Fli1 deficiency may contribute to the development of tissue fibrosis, vasculopathy and immune activation in systemic sclerosis. Exp. Dermatol. 2018, 27, 1030–1037. [Google Scholar] [CrossRef]
- Perreau, M.; Suffiotti, M.; Marques-Vidal, P.; Wiedemann, A.; Levy, Y.; Laouénan, C.; Ghosn, J.; Fenwick, C.; Comte, D.; Roger, T.; et al. The cytokines HGF and CXCL13 predict the severity and the mortality in COVID-19 patients. Nat. Commun. 2021, 12, 4888. [Google Scholar] [CrossRef]
- Kramer, G.; Mitteregger, D.; Marberger, M. Is benign prostatic hyperplasia (BPH) an immune inflammatory disease? Eur. Urol. 2007, 51, 1202–1216. [Google Scholar] [CrossRef]
- Wang, L.; Yang, J.R.; Yang, L.Y.; Liu, Z.T. Chronic inflammation in benign prostatic hyperplasia: Implications for therapy. Med. Hypotheses 2008, 70, 1021–1023. [Google Scholar] [CrossRef]
- Gandaglia, G.; Briganti, A.; Gontero, P.; Mondaini, N.; Novara, G.; Salonia, A.; Sciarra, A.; Montorsi, F. The role of chronic prostatic inflammation in the pathogenesis and progression of benign prostatic hyperplasia (BPH). BJU Int. 2013, 112, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Penna, G.; Fibbi, B.; Amuchastegui, S.; Cossetti, C.; Aquilano, F.; Laverny, G.; Gacci, M.; Crescioli, C.; Maggi, M.; Adorini, L. Human benign prostatic hyperplasia stromal cells as inducers and targets of chronic immuno-mediated inflammation. J. Immunol. 2009, 182, 4056–4064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fibbi, B.; Penna, G.; Morelli, A.; Adorini, L.; Maggi, M. Chronic inflammation in the pathogenesis of benign prostatic hyperplasia. Int. J. Androl. 2010, 33, 475–488. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, J.T.; Coffey, D.S. Etiology and disease process of benign prostatic hyperplasia. Prostate Suppl. 1989, 2, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, L.; Yao, L.; Sun, S.; Shen, L. CXCL13 promotes proliferation and autophagy of human mesenchymal stem cells through MAPK signaling pathway under hypoxic condition. Chin. J. Cell. Mol. Immunol. 2019, 35, 211–216. [Google Scholar]
- Da, Z.; Li, L.; Zhu, J.; Gu, Z.; You, B.; Shan, Y.; Shi, S. CXCL13 Promotes Proliferation of Mesangial Cells by Combination with CXCR5 in SLE. J. Immunol. Res. 2016, 2016, 2063985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lisignoli, G.; Toneguzzi, S.; Piacentini, A.; Cristino, S.; Grassi, F.; Cavallo, C.; Facchini, A. CXCL12 (SDF-1) and CXCL13 (BCA-1) chemokines significantly induce proliferation and collagen type I expression in osteoblasts from osteoarthritis patients. J. Cell. Physiol. 2006, 206, 78–85. [Google Scholar] [CrossRef]
- Liu, G.; Wang, Y.; Jiang, S.; Sui, M.; Wang, C.; Kang, L.; Sun, Y.; Jiang, Y. Suppression of lymphocyte apoptosis in spleen by CXCL13 after porcine circovirus type 2 infection and regulatory mechanism of CXCL13 expression in pigs. Vet. Res. 2019, 50, 17. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.J.; Jiang, L.; Tong, D.Y.; Ren, Y.N.; Sheng, M.F.; Liu, H.C. CXCL13 inhibition induce the apoptosis of MDA-MB-231 breast cancer cells through blocking CXCR5/ERK signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 8755–8762. [Google Scholar] [CrossRef]
- De Nunzio, C.; Kramer, G.; Marberger, M.; Montironi, R.; Nelson, W.; Schröder, F.; Sciarra, A.; Tubaro, A. The controversial relationship between benign prostatic hyperplasia and prostate cancer: The role of inflammation. Eur. Urol. 2011, 60, 106–117. [Google Scholar] [CrossRef]
- Wynn, T.A. Cellular and molecular mechanisms of fibrosis. J. Pathol. 2008, 214, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Gharaee-Kermani, M.; Kunju, L.; Hollingsworth, J.M.; Adler, J.; Arruda, E.M.; Macoska, J.A. Prostatic fibrosis is associated with lower urinary tract symptoms. J. Urol. 2012, 188, 1375–1381. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Nieves, J.A.; Macoska, J.A. Prostatic fibrosis, lower urinary tract symptoms, and BPH. Nat. Rev. Urol. 2013, 10, 546–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bushman, W.A.; Jerde, T.J. The role of prostate inflammation and fibrosis in lower urinary tract symptoms. Am. J. Physiol. Ren. Physiol. 2016, 311, F817–F821. [Google Scholar] [CrossRef] [Green Version]
- Biswas, S.; Sengupta, S.; Roy Chowdhury, S.; Jana, S.; Mandal, G.; Mandal, P.K.; Saha, N.; Malhotra, V.; Gupta, A.; Kuprash, D.V.; et al. CXCL13-CXCR5 co-expression regulates epithelial to mesenchymal transition of breast cancer cells during lymph node metastasis. Breast Cancer Res. Treat. 2014, 143, 265–276. [Google Scholar] [CrossRef]
- Xie, Y.; Chen, Z.; Zhong, Q.; Zheng, Z.; Chen, Y.; Shangguan, W.; Zhang, Y.; Yang, J.; Zhu, D.; Xie, W. M2 macrophages secrete CXCL13 to promote renal cell carcinoma migration, invasion, and EMT. Cancer Cell Int. 2021, 21, 677. [Google Scholar] [CrossRef]
- Ammirante, M.; Shalapour, S.; Kang, Y.; Jamieson, C.A.; Karin, M. Tissue injury and hypoxia promote malignant progression of prostate cancer by inducing CXCL13 expression in tumor myofibroblasts. Proc. Natl. Acad. Sci. USA 2014, 111, 14776–14781. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Singh, R.; Sharma, P.K.; Singh, U.P.; Rai, S.N.; Chung, L.W.; Cooper, C.R.; Novakovic, K.R.; Grizzle, W.E.; Lillard, J.W., Jr. Serum CXCL13 positively correlates with prostatic disease, prostate-specific antigen and mediates prostate cancer cell invasion, integrin clustering and cell adhesion. Cancer Lett. 2009, 283, 29–35. [Google Scholar] [CrossRef] [Green Version]
- El-Haibi, C.P.; Singh, R.; Gupta, P.; Sharma, P.K.; Greenleaf, K.N.; Singh, S.; Lillard, J.W., Jr. Antibody Microarray Analysis of Signaling Networks Regulated by Cxcl13 and Cxcr5 in Prostate Cancer. J. Proteom. Bioinform. 2012, 5, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Pascal, L.E.; Stolz, D.B.; Wang, K.; Zhou, Y.; Chen, W.; Xu, Y.; Chen, Y.; Dhir, R.; Parwani, A.V.; et al. E-cadherin is downregulated in benign prostatic hyperplasia and required for tight junction formation and permeability barrier in the prostatic epithelial cell monolayer. Prostate 2019, 79, 1226–1237. [Google Scholar] [CrossRef]
- Chen, W.; Pascal, L.E.; Wang, K.; Dhir, R.; Sims, A.M.; Campbell, R.; Gasper, G.; DeFranco, D.B.; Yoshimura, N.; Wang, Z. Differential impact of paired patient-derived BPH and normal adjacent stromal cells on benign prostatic epithelial cell growth in 3D culture. Prostate 2020, 80, 1177–1187. [Google Scholar] [CrossRef] [PubMed]
- Fraga, C.H.; True, L.D.; Kirk, D. Enhanced expression of the mesenchymal marker, vimentin, in hyperplastic versus normal human prostatic epithelium. J. Urol. 1998, 159, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Magdalena, P.; Brössner, C.; Reiner, A.; Cheng, G.; Sugiyama, N.; Warner, M.; Gustafsson, J.A. A role for epithelial-mesenchymal transition in the etiology of benign prostatic hyperplasia. Proc. Natl. Acad. Sci. USA 2009, 106, 2859–2863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slabáková, E.; Pernicová, Z.; Slavíčková, E.; Staršíchová, A.; Kozubík, A.; Souček, K. TGF-β1-induced EMT of non-transformed prostate hyperplasia cells is characterized by early induction of SNAI2/Slug. Prostate 2011, 71, 1332–1343. [Google Scholar] [CrossRef]
- Hu, S.; Yu, W.; Lv, T.J.; Chang, C.S.; Li, X.; Jin, J. Evidence of TGF-β1 mediated epithelial-mesenchymal transition in immortalized benign prostatic hyperplasia cells. Mol. Membr. Biol. 2014, 31, 103–110. [Google Scholar] [CrossRef]
- Broster, S.A.; Kyprianou, N. Epithelial-mesenchymal transition in prostatic disease. Future Oncol. 2015, 11, 3197–3206. [Google Scholar] [CrossRef]
- Zhao, Q.; Guo, J.; Wang, G.; Bi, Y.; Cheng, X.; Liao, Y.; Jin, S.; Li, L.; Guo, Y.; Pan, L.; et al. CXCL13 promotes intestinal tumorigenesis through the activation of epithelial AKT signaling. Cancer Lett. 2021, 511, 1–14. [Google Scholar] [CrossRef]
- Zheng, Z.; Cai, Y.; Chen, H.; Chen, Z.; Zhu, D.; Zhong, Q.; Xie, W. CXCL13/CXCR5 Axis Predicts Poor Prognosis and Promotes Progression Through PI3K/AKT/mTOR Pathway in Clear Cell Renal Cell Carcinoma. Front. Oncol. 2018, 8, 682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Chen, S.; Yang, C.; Zhou, M.; Yang, T.; Sun, B.; Zhu, J.; Zhang, H.; Lu, Q.; Li, L.; et al. Activation of CXCL13/CXCR5 axis aggravates experimental autoimmune cystitis and interstitial cystitis/bladder pain syndrome. Biochem. Pharmacol. 2022, 200, 115047. [Google Scholar] [CrossRef]
- Alvarez, E.; Piccio, L.; Mikesell, R.J.; Klawiter, E.C.; Parks, B.J.; Naismith, R.T.; Cross, A.H. CXCL13 is a biomarker of inflammation in multiple sclerosis, neuromyelitis optica, and other neurological conditions. Mult. Scler. 2013, 19, 1204–1208. [Google Scholar] [CrossRef] [Green Version]
- Bugatti, S.; Manzo, A.; Vitolo, B.; Benaglio, F.; Binda, E.; Scarabelli, M.; Humby, F.; Caporali, R.; Pitzalis, C.; Montecucco, C. High expression levels of the B cell chemoattractant CXCL13 in rheumatoid synovium are a marker of severe disease. Rheumatology 2014, 53, 1886–1895. [Google Scholar] [CrossRef] [PubMed]
- Mack, M. Inflammation and fibrosis. Matrix Biol. J. Int. Soc. Matrix Biol. 2018, 68–69, 106–121. [Google Scholar] [CrossRef] [PubMed]
- Vuga, L.J.; Tedrow, J.R.; Pandit, K.V.; Tan, J.; Kass, D.J.; Xue, J.; Chandra, D.; Leader, J.K.; Gibson, K.F.; Kaminski, N.; et al. C-X-C motif chemokine 13 (CXCL13) is a prognostic biomarker of idiopathic pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 2014, 189, 966–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djavan, B.; Lin, V.; Seitz, C.; Kramer, G.; Kaplan, P.; Richier, J.; Marberger, M.; McConnell, J.D. Elastin gene expression in benign prostatic hyperplasia. Prostate 1999, 40, 242–247. [Google Scholar] [CrossRef]
- Cantiello, F.; Cicione, A.; Salonia, A.; Autorino, R.; Tucci, L.; Madeo, I.; Damiano, R. Periurethral fibrosis secondary to prostatic inflammation causing lower urinary tract symptoms: A prospective cohort study. Urology 2013, 81, 1018–1023. [Google Scholar] [CrossRef]
- Bauman, T.M.; Nicholson, T.M.; Abler, L.L.; Eliceiri, K.W.; Huang, W.; Vezina, C.M.; Ricke, W.A. Characterization of fibrillar collagens and extracellular matrix of glandular benign prostatic hyperplasia nodules. PLoS ONE 2014, 9, e109102. [Google Scholar] [CrossRef] [Green Version]
- Pinsky, P.F.; Kramer, B.S.; Crawford, E.D.; Grubb, R.L.; Urban, D.A.; Andriole, G.L.; Chia, D.; Levin, D.L.; Gohagan, J.K. Prostate volume and prostate-specific antigen levels in men enrolled in a large screening trial. Urology 2006, 68, 352–356. [Google Scholar] [CrossRef]
- Mochtar, C.A.; Kiemeney, L.A.; van Riemsdijk, M.M.; Barnett, G.S.; Laguna, M.P.; Debruyne, F.M.; de la Rosette, J.J. Prostate-specific antigen as an estimator of prostate volume in the management of patients with symptomatic benign prostatic hyperplasia. Eur. Urol. 2003, 44, 695–700. [Google Scholar] [CrossRef]
- Bosch, J.L.; Bohnen, A.M.; Groeneveld, F.P. Validity of digital rectal examination and serum prostate specific antigen in the estimation of prostate volume in community-based men aged 50 to 78 years: The Krimpen Study. Eur. Urol. 2004, 46, 753–759. [Google Scholar] [CrossRef]
- Putra, I.B.; Hamid, A.R.; Mochtar, C.A.; Umbas, R. Relationship of age, prostate-specific antigen, and prostate volume in Indonesian men with benign prostatic hyperplasia. Prostate Int. 2016, 4, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Jensen, E.C. Quantitative analysis of histological staining and fluorescence using ImageJ. Anat. Rec. 2013, 296, 378–381. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Body Weight (g) | Ventral Prostate Weight (mg) | Prostate Index | |
|---|---|---|---|---|
| Initial | Final | |||
| sham | 337.6 (17.9) | 473.2 (37.8) | 598.7 (81.1) | 1.3(0.3) |
| rHuCXCL13 | 339.1 (8.1) | 468.3 (38.2) | 759.8 (83.1) * | 1.7 (0.2) * |
| Mean | SD | |
|---|---|---|
| CXCL13 score | 0.6 | 0.4 |
| Age (years) | 70.1 | 7.5 |
| BMI (kg/m2) | 22.8 | 2.8 |
| PV (cm3) | 60.8 | 36.7 |
| tPSA (ng/mL) | 7.0 | 5.9 |
| fPSA (ng/mL) | 1.6 | 1.4 |
| Qmax (mL/s) | 10.0 | 5.9 |
| RUV (mL) | 160.7 | 114.5 |
| IPSS | 21.7 | 7.4 |
| nocturnal enuresis | 3.1 | 2.0 |
| Correlation Coefficient | p Value | |
|---|---|---|
| Age | −0.150 | 0.131 |
| BMI | 0.124 | 0.211 |
| PV | 0.230 | 0.028 * |
| tPSA | 0.235 | 0.027 * |
| fPSA | 0.149 | 0.167 |
| Qmax | 0.112 | 0.502 |
| RUV | -0.060 | 0.687 |
| IPSS | 0.074 | 0.457 |
| nocturnal enuresis | 0.040 | 0.687 |
| Target Gene | Human (5′ to 3′) | |
|---|---|---|
| CXCL13 | Forward | GCTTGAGGTGTAGATGTGTCC |
| Reverse | CCCACGGGGCAAGATTTGAA | |
| IL-6 | Forward | GAACTCCTTCTCCACAAGCG |
| Reverse | GCCTCTTTGCTGCTTTCACA | |
| IL-8 | Forward | TCCAAACCTTTCCACCCCAAA |
| Reverse | TTTCTGTGTTGGCGCAGTGT | |
| TNF-α | Forward | GGTCCTCTTCAAGGGCCAAG |
| Reverse | TCACAGGGCAATGATCCCAA | |
| α-SMA | Forward | GGCATTCACGAGACCACCTAC |
| Reverse | CGACATGACGTTGTTGGCATAC | |
| Collagen Ⅰ | Forward | GAGGGCCAAGACGAAGACATC |
| Reverse | CAGATCACGTCATCGCACAAC | |
| GAPDH | Forward | ATCCCATCACCATCTTCCAGGAG |
| Reverse | CCTGCTTCACCACCTTCTTGATG | |
| Antigens | Species | Dilution (IF) | Dilution (IHC) | Dilution (WB) | Supplier |
|---|---|---|---|---|---|
| CDK2 | monoclonal | 1:1000 | CST, USA, Cat. 2546 | ||
| CDK4 | monoclonal | 1:1000 | CST, USA, Cat. 12790 | ||
| Cyclin D1 | monoclonal | 1:1000 | CST, USA, Cat. 2978 | ||
| BAX | monoclonal | 1:1000 | Abclonal, CHN, Cat. A19684 | ||
| BCL-2 | monoclonal | 1:1000 | Abclonal, CHN, Cat. A19693 | ||
| E-cadherin | polyclonal | 1:1000 | Abclonal, CHN, Cat. A3044 | ||
| N-cadherin | monoclonal | 1:1000 | Abclonal, CHN, Cat. A19083 | ||
| α-SMA | monoclonal | 1:100 | 1:1000 | Abclonal, CHN, Cat. A17910 | |
| Collagen Ⅰ | polyclonal | 1:100 | 1:1000 | Abclonal, CHN, Cat. A1352 | |
| TNF-α | polyclonal | 1:1000 | Abclonal, CHN, Cat. A0277 | ||
| IL-6 | polyclonal | 1:1000 | Abclonal, CHN, Cat. A0286 | ||
| IL-8 | polyclonal | 1:1000 | Abclonal, CHN, Cat. A2541 | ||
| CXCL13 | monoclonal | 1:500 | 1:500 | 1:1000 | Abcam, UK, Cat.ab112521 |
| pAKT | monoclonal | 1:1000 | Abclonal, CHN, Cat. AP0637 | ||
| AKT | monoclonal | 1:1000 | Abclonal, CHN, Cat. A17909 | ||
| pERK1/2 | monoclonal | 1:1000 | Abclonal, CHN, Cat. AP0472 | ||
| ERK1/2 | monoclonal | 1:1000 | Abclonal, CHN, Cat. A4782 | ||
| pSTAT3 | monoclonal | 1:1000 | Abclonal, CHN, Cat. AP0715 | ||
| STAT3 | polyclonal | 1:1000 | Abclonal, CHN, Cat. A1192 | ||
| GAPDH | monoclonal | 1:1000 | Abclonal, CHN, Cat. AC001 |
| Secondary Detection System Used | Host | Dilution Used | Supplier |
|---|---|---|---|
| Anti-Rabbit-IgG (H + L)-HRP | Goat | 1:10000 (WB) | Sungene Biotech, Tianjin, China, Cat. #LK2001 |
| 1:1000 (IHC) | |||
| Anti-Rabbit IgG (H + L), F(ab’)2 fragment (Alexa Fluor® (Irving, USA) 488 Conjugate) | Goat | 1:50 (IF) | Cell Signaling Technology, Boston, USA, Cat. 4412 |
| Hoechst 33342 (1 mg/mL) nucleic acid staining (DAPI) | - | 1:750 (IF) | Molecular Probes/Invitrogen, Carlsbad, USA, Cat. A11007 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, D.; Li, M.; Fu, X.; Yang, S.; Wang, Z.; Liu, J.; Li, Y.; Zhou, Y.; Ren, P.; Guo, Y.; et al. Changes in the Expression and Functional Activities of C-X-C Motif Chemokine Ligand 13 (CXCL13) in Hyperplastic Prostate. Int. J. Mol. Sci. 2023, 24, 56. https://doi.org/10.3390/ijms24010056
Liu D, Li M, Fu X, Yang S, Wang Z, Liu J, Li Y, Zhou Y, Ren P, Guo Y, et al. Changes in the Expression and Functional Activities of C-X-C Motif Chemokine Ligand 13 (CXCL13) in Hyperplastic Prostate. International Journal of Molecular Sciences. 2023; 24(1):56. https://doi.org/10.3390/ijms24010056
Chicago/Turabian StyleLiu, Daoquan, Mingzhou Li, Xun Fu, Shu Yang, Zhen Wang, Jianmin Liu, Yan Li, Yongying Zhou, Pengfei Ren, Yuhang Guo, and et al. 2023. "Changes in the Expression and Functional Activities of C-X-C Motif Chemokine Ligand 13 (CXCL13) in Hyperplastic Prostate" International Journal of Molecular Sciences 24, no. 1: 56. https://doi.org/10.3390/ijms24010056