
Role and Dysregulation of miRNA in Patients with Parkinson’s Disease

 ,
,  , and
, and
Abstract
:1. Introduction
microRNAs and Parkinson’s Disease
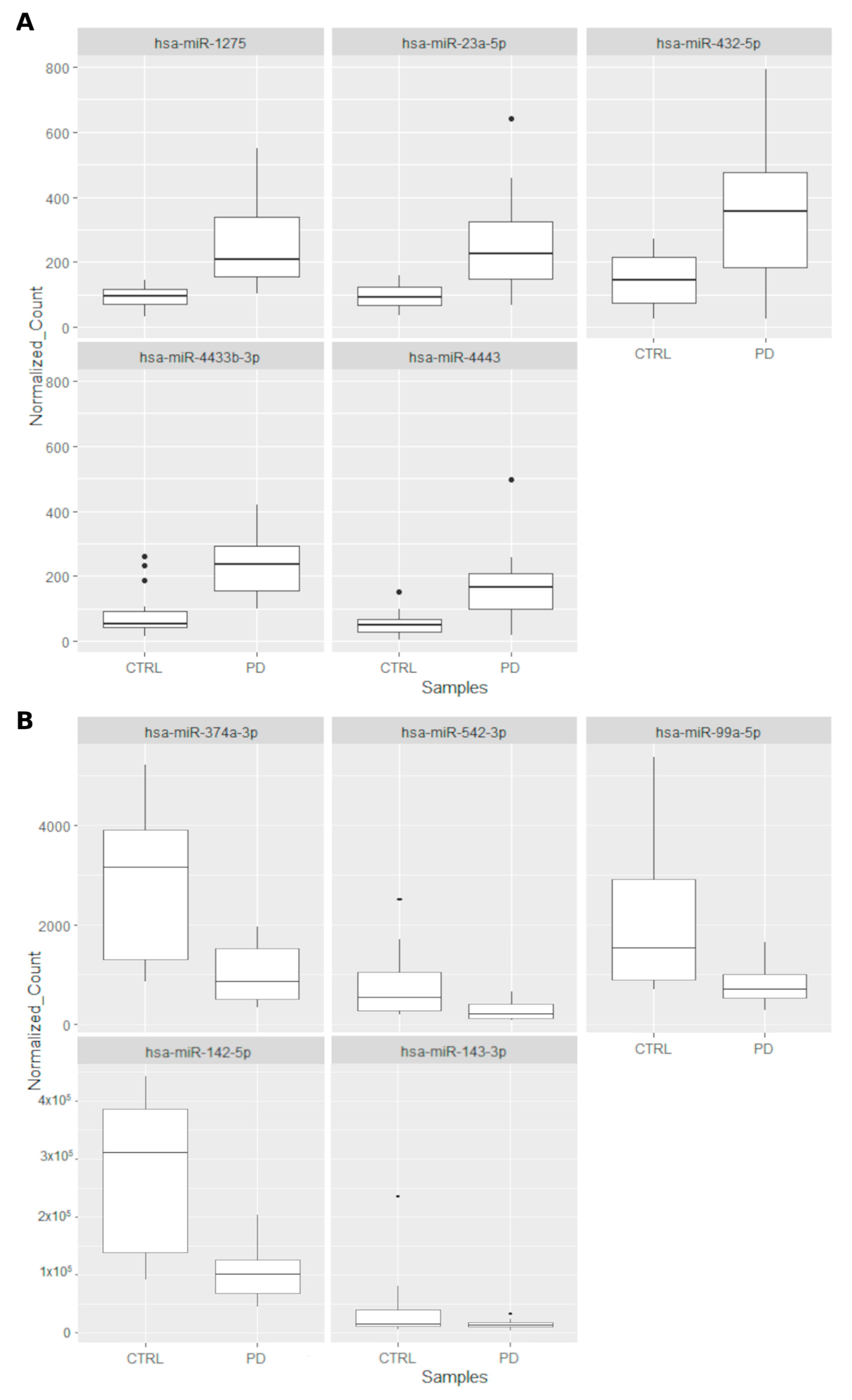
2. Results
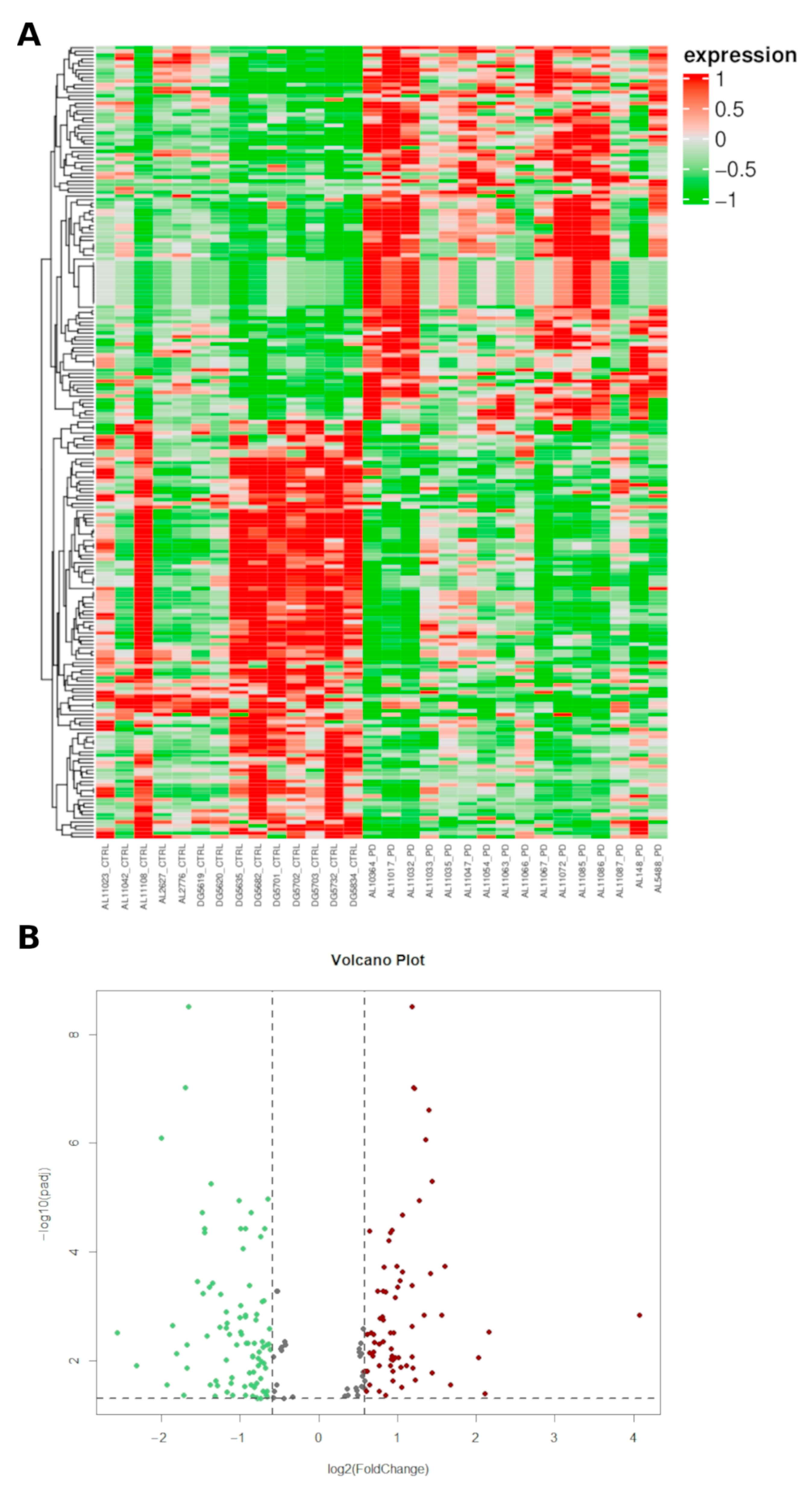
2.1. Characterization of Small Non Coding RNAs Deregulation in PD Patients
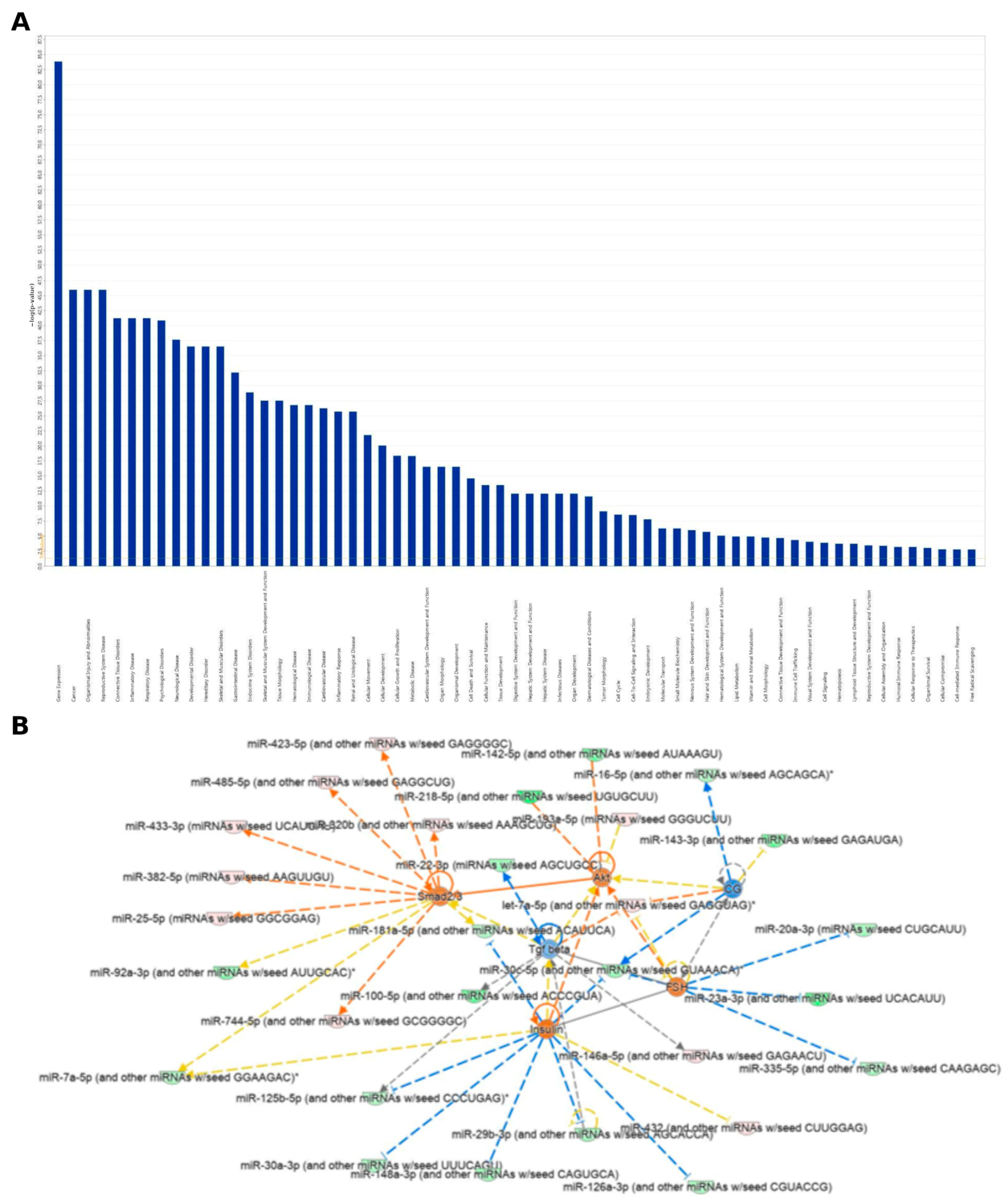
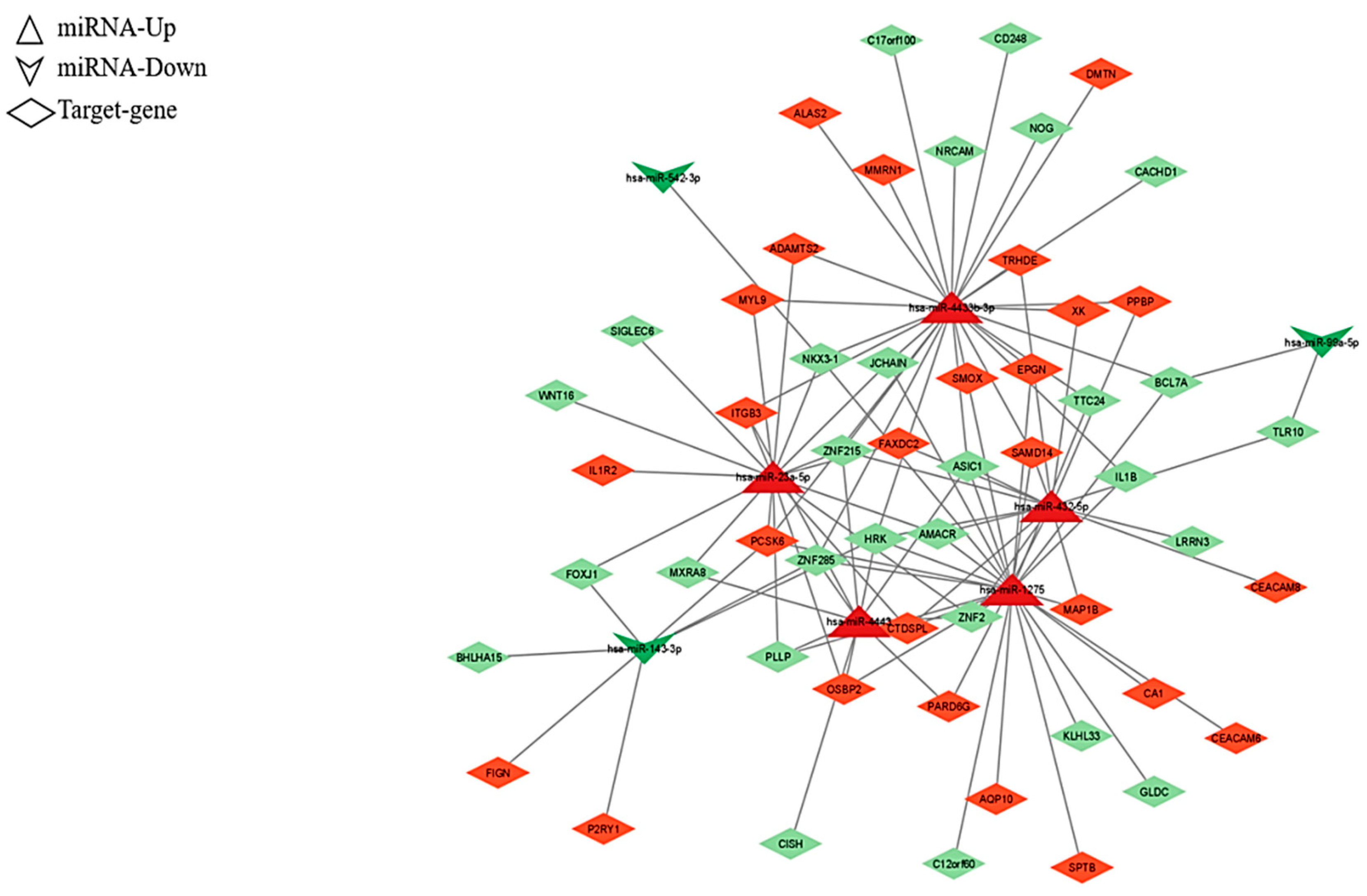
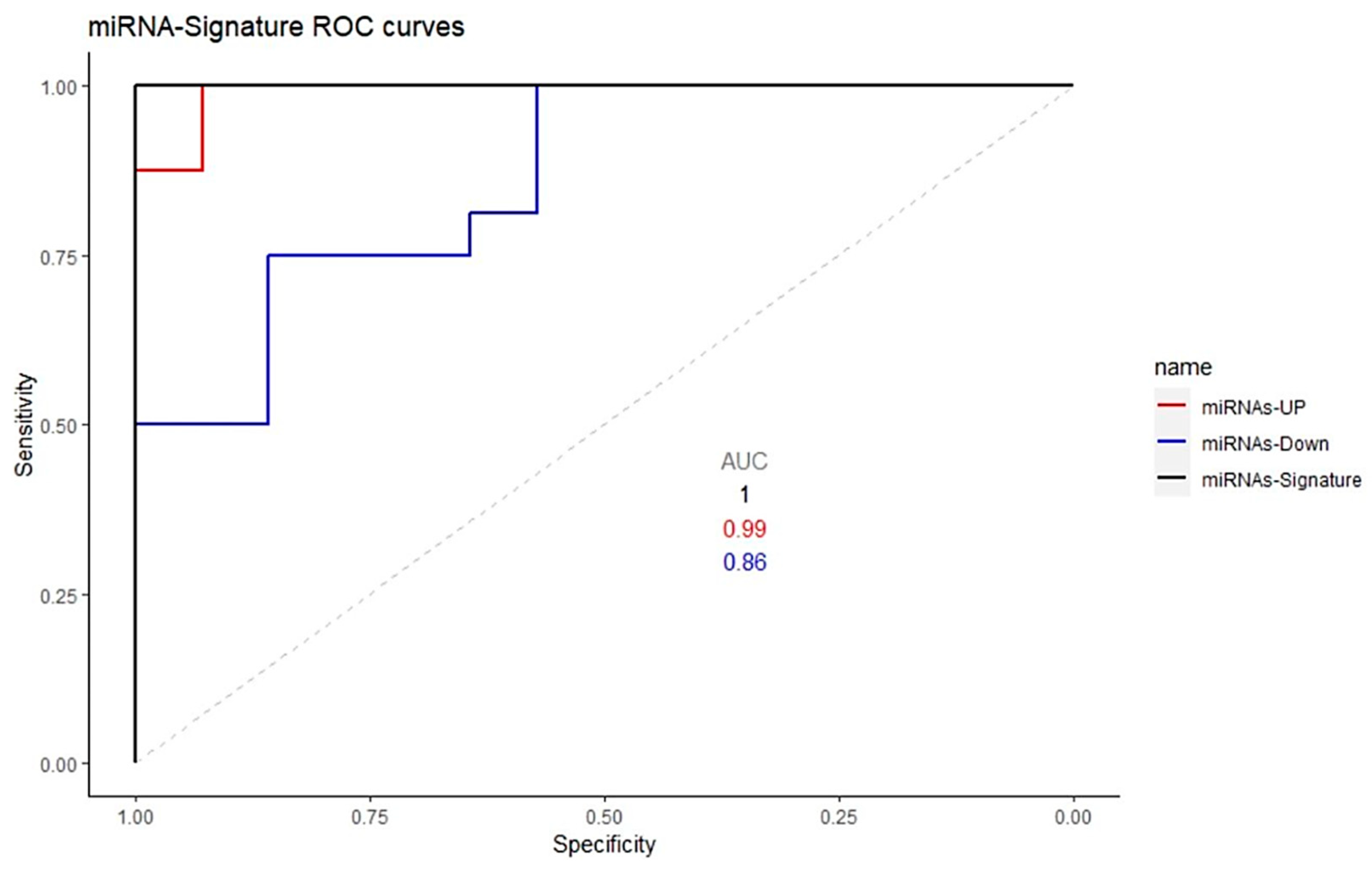
2.2. Analysis of the Differentially Expressed miRNAs’ Target Genes
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. miRNA Extraction
4.3. miRNA Sequencing and Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.E.; Lang, A.E. Parkinson disease. Nat. Rev. Dis. Prim. 2017, 3, 17013. [Google Scholar] [CrossRef] [PubMed]
- Leggio, L.; Vivarelli, S.; L’Episcopo, F.; Tirolo, C.; Caniglia, S.; Testa, N.; Marchetti, B.; Iraci, N. microRNAs in Parkinson’s Disease: From Pathogenesis to Novel Diagnostic and Therapeutic Approaches. Int. J. Mol. Sci. 2017, 18, 2698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, J.; Takousis, P.; Wohlers, I.; Itua, I.O.G.; Dobricic, V.; Rucker, G.; Binder, H.; Middleton, L.; Ioannidis, J.P.A.; Perneczky, R.; et al. Meta-analyses identify differentially expressed micrornas in Parkinson’s disease. Ann. Neurol. 2019, 85, 835–851. [Google Scholar] [CrossRef] [PubMed]
- Santos-Lobato, B.L.; Vidal, A.F.; Ribeiro-Dos-Santos, A. Regulatory miRNA-mRNA Networks in Parkinson’s Disease. Cells 2021, 10, 1410. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef] [Green Version]
- Ruby, J.G.; Jan, C.H.; Bartel, D.P. Intronic microRNA precursors that bypass Drosha processing. Nature 2007, 448, 83–86. [Google Scholar] [CrossRef] [Green Version]
- Ruby, J.G.; Stark, A.; Johnston, W.K.; Kellis, M.; Bartel, D.P.; Lai, E.C. Evolution, biogenesis, expression, and target predictions of a substantially expanded set of Drosophila microRNAs. Genome Res. 2007, 17, 1850–1864. [Google Scholar] [CrossRef] [Green Version]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef]
- Lu, J.; Xu, Y.; Quan, Z.; Chen, Z.; Sun, Z.; Qing, H. Dysregulated microRNAs in neural system: Implication in pathogenesis and biomarker development in Parkinson’s disease. Neuroscience 2017, 365, 70–82. [Google Scholar] [CrossRef]
- Lagos-Quintana, M.; Rauhut, R.; Yalcin, A.; Meyer, J.; Lendeckel, W.; Tuschl, T. Identification of tissue-specific microRNAs from mouse. Curr. Biol. CB 2002, 12, 735–739. [Google Scholar] [CrossRef]
- Rajman, M.; Schratt, G. MicroRNAs in neural development: From master regulators to fine-tuners. Development 2017, 144, 2310–2322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schratt, G. microRNAs at the synapse. Nat. Rev. Neurosci. 2009, 10, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Antoniou, A.; Khudayberdiev, S.; Idziak, A.; Bicker, S.; Jacob, R.; Schratt, G. The dynamic recruitment of TRBP to neuronal membranes mediates dendritogenesis during development. EMBO Rep. 2018, 19, e44853. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, S.R.; Dionisio, P.A.; Correia Guedes, L.; Goncalves, N.; Coelho, M.; Rosa, M.M.; Amaral, J.D.; Ferreira, J.J.; Rodrigues, C.M.P. Circulating Inflammatory miRNAs Associated with Parkinson’s Disease Pathophysiology. Biomolecules 2020, 10, 945. [Google Scholar] [CrossRef]
- Danborg, P.B.; Simonsen, A.H.; Waldemar, G.; Heegaard, N.H. The potential of microRNAs as biofluid markers of neurodegenerative diseases--a systematic review. Biomark. Biochem. Indic. Expo. Response Susceptibility Chem. 2014, 19, 259–268. [Google Scholar] [CrossRef]
- Van den Hove, D.L.; Kompotis, K.; Lardenoije, R.; Kenis, G.; Mill, J.; Steinbusch, H.W.; Lesch, K.P.; Fitzsimons, C.P.; De Strooper, B.; Rutten, B.P. Epigenetically regulated microRNAs in Alzheimer’s disease. Neurobiol. Aging 2014, 35, 731–745. [Google Scholar] [CrossRef]
- Hoye, M.L.; Koval, E.D.; Wegener, A.J.; Hyman, T.S.; Yang, C.; O’Brien, D.R.; Miller, R.L.; Cole, T.; Schoch, K.M.; Shen, T.; et al. MicroRNA Profiling Reveals Marker of Motor Neuron Disease in ALS Models. J. Neurosci. Off. J. Soc. Neurosci. 2017, 37, 5574–5586. [Google Scholar] [CrossRef] [Green Version]
- Butovsky, O.; Siddiqui, S.; Gabriely, G.; Lanser, A.J.; Dake, B.; Murugaiyan, G.; Doykan, C.E.; Wu, P.M.; Gali, R.R.; Iyer, L.K.; et al. Modulating inflammatory monocytes with a unique microRNA gene signature ameliorates murine ALS. J. Clin. Investig. 2012, 122, 3063–3087. [Google Scholar] [CrossRef] [Green Version]
- Junker, A.; Krumbholz, M.; Eisele, S.; Mohan, H.; Augstein, F.; Bittner, R.; Lassmann, H.; Wekerle, H.; Hohlfeld, R.; Meinl, E. MicroRNA profiling of multiple sclerosis lesions identifies modulators of the regulatory protein CD47. Brain J. Neurol. 2009, 132, 3342–3352. [Google Scholar] [CrossRef] [Green Version]
- Moore, C.S.; Rao, V.T.; Durafourt, B.A.; Bedell, B.J.; Ludwin, S.K.; Bar-Or, A.; Antel, J.P. miR-155 as a multiple sclerosis-relevant regulator of myeloid cell polarization. Ann. Neurol. 2013, 74, 709–720. [Google Scholar] [CrossRef]
- Goh, S.Y.; Chao, Y.X.; Dheen, S.T.; Tan, E.K.; Tay, S.S. Role of MicroRNAs in Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20, 5649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gentile, G.; Morello, G.; La Cognata, V.; Guarnaccia, M.; Conforti, F.L.; Cavallaro, S. Dysregulated miRNAs as Biomarkers and Therapeutical Targets in Neurodegenerative Diseases. J. Pers. Med. 2022, 12, 770. [Google Scholar] [CrossRef] [PubMed]
- Thomas, K.T.; Gross, C.; Bassell, G.J. microRNAs Sculpt Neuronal Communication in a Tight Balance That Is Lost in Neurological Disease. Front. Mol. Neurosci. 2018, 11, 455. [Google Scholar] [CrossRef] [PubMed]
- Konovalova, J.; Gerasymchuk, D.; Parkkinen, I.; Chmielarz, P.; Domanskyi, A. Interplay between MicroRNAs and Oxidative Stress in Neurodegenerative Diseases. Int. J. Mol. Sci. 2019, 20, 6055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.P.; Yu, S.H.; Zhang, G.H.; Hou, Y.B.; Gu, X.J.; Ou, R.W.; Shen, Y.; Song, W.; Chen, X.P.; Zhao, B.; et al. The mutation spectrum of Parkinson-disease-related genes in early-onset Parkinson’s disease in ethnic Chinese. Eur. J. Neurol. 2022, 29, 3218–3228. [Google Scholar] [CrossRef]
- Emekli, I.; Tepgec, F.; Samanci, B.; Toksoy, G.; Hasanogullari Kina, G.; Tufekcioglu, Z.; Basaran, S.; Bilgic, B.; Gurvit, I.H.; Emre, M.; et al. Clinical and molecular genetic findings of hereditary Parkinson’s patients from Turkey. Park. Relat. Disord. 2021, 93, 35–39. [Google Scholar] [CrossRef]
- Book, A.; Guella, I.; Candido, T.; Brice, A.; Hattori, N.; Jeon, B.; Farrer, M.J.; Consortium, S.M.I.o.t.G. A Meta-Analysis of alpha-Synuclein Multiplication in Familial Parkinsonism. Front. Neurol. 2018, 9, 1021. [Google Scholar] [CrossRef]
- Catala-Lopez, F.; Suarez-Pinilla, M.; Suarez-Pinilla, P.; Valderas, J.M.; Gomez-Beneyto, M.; Martinez, S.; Balanza-Martinez, V.; Climent, J.; Valencia, A.; McGrath, J.; et al. Inverse and direct cancer comorbidity in people with central nervous system disorders: A meta-analysis of cancer incidence in 577,013 participants of 50 observational studies. Psychother. Psychosom. 2014, 83, 89–105. [Google Scholar] [CrossRef]
- Ibanez, K.; Boullosa, C.; Tabares-Seisdedos, R.; Baudot, A.; Valencia, A. Molecular evidence for the inverse comorbidity between central nervous system disorders and cancers detected by transcriptomic meta-analyses. PLoS Genet. 2014, 10, e1004173. [Google Scholar] [CrossRef]
- Salemi, M.; Mogavero, M.P.; Lanza, G.; Mongioi, L.M.; Calogero, A.E.; Ferri, R. Examples of Inverse Comorbidity between Cancer and Neurodegenerative Diseases: A Possible Role for Noncoding RNA. Cells 2022, 11, 1930. [Google Scholar] [CrossRef]
- Houck, A.L.; Seddighi, S.; Driver, J.A. At the Crossroads Between Neurodegeneration and Cancer: A Review of Overlapping Biology and Its Implications. Curr. Aging Sci. 2018, 11, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Pepe, P.; Vatrano, S.; Cannarella, R.; Calogero, A.E.; Marchese, G.; Ravo, M.; Fraggetta, F.; Pepe, L.; Pennisi, M.; Romano, C.; et al. A study of gene expression by RNA-seq in patients with prostate cancer and in patients with Parkinson disease: An example of inverse comorbidity. Mol. Biol. Rep. 2021, 48, 7627–7631. [Google Scholar] [CrossRef] [PubMed]
- Mogavero, M.P.; Silvani, A.; DelRosso, L.M.; Salemi, M.; Ferri, R. Focus on the Complex Interconnection between Cancer, Narcolepsy and Other Neurodegenerative Diseases: A Possible Case of Orexin-Dependent Inverse Comorbidity. Cancers 2021, 13, 2612. [Google Scholar] [CrossRef] [PubMed]
- Fores-Martos, J.; Boullosa, C.; Rodrigo-Dominguez, D.; Sanchez-Valle, J.; Suay-Garcia, B.; Climent, J.; Falco, A.; Valencia, A.; Puig-Butille, J.A.; Puig, S.; et al. Transcriptomic and Genetic Associations between Alzheimer’s Disease, Parkinson’s Disease, and Cancer. Cancers 2021, 13, 2990. [Google Scholar] [CrossRef] [PubMed]
- Monsivais, D.; Peng, J.; Kang, Y.; Matzuk, M.M. Activin-like kinase 5 (ALK5) inactivation in the mouse uterus results in metastatic endometrial carcinoma. Proc. Natl. Acad. Sci. USA 2019, 116, 3883–3892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.L.; He, J.P.; Zhu, C.; Cheng, H.Z. Endometrial carcinoma may favor partial, but not complete, loss of the TGF-β signaling pathway. Proc. Natl. Acad. Sci. USA 2019, 116, 9164–9165. [Google Scholar] [CrossRef] [Green Version]
- Seoane, J.; Gomis, R.R. TGF-β Family Signaling in Tumor Suppression and Cancer Progression. Cold Spring Harbor Perspect. Biol. 2017, 9, a022277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drabsch, Y.; Dijke, P. TGF-β signalling and its role in cancer progression and metastasis. Cancer Metastasis Rev. 2012, 31, 553–568. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Lloret, S.; Cardinali, D.P. Melatonin as a Chronobiotic and Cytoprotective Agent in Parkinson’s Disease. Front. Pharmacol. 2021, 12, 650597. [Google Scholar] [CrossRef]
- Figorilli, M.; Lanza, G.; Congiu, P.; Lecca, R.; Casaglia, E.; Mogavero, M.P.; Puligheddu, M.; Ferri, R. Neurophysiological Aspects of REM Sleep Behavior Disorder (RBD): A Narrative Review. Brain Sci. 2021, 11, 1588. [Google Scholar] [CrossRef]
- Chen, D.; Zhang, T.; Lee, T.H. Cellular Mechanisms of Melatonin: Insight from Neurodegenerative Diseases. Biomolecules 2020, 10, 1158. [Google Scholar] [CrossRef] [PubMed]
- Luo, F.; Sandhu, A.F.; Rungratanawanich, W.; Williams, G.E.; Akbar, M.; Zhou, S.; Song, B.J.; Wang, X. Melatonin and Autophagy in Aging-Related Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 7174. [Google Scholar] [CrossRef] [PubMed]
- Tamtaji, O.R.; Reiter, R.J.; Alipoor, R.; Dadgostar, E.; Kouchaki, E.; Asemi, Z. Melatonin and Parkinson Disease: Current Status and Future Perspectives for Molecular Mechanisms. Cell. Mol. Neurobiol. 2020, 40, 15–23. [Google Scholar] [CrossRef] [PubMed]
- De Araújo, D.P.; Lobato, R.D.E.F.; Cavalcanti, J.R.; Sampaio, L.R.; Araújo, P.V.; Silva, M.C.; Neves, K.R.; Fonteles, M.M.; Sousa, F.C.; Vasconcelos, S.M. The contributions of antioxidant activity of lipoic acid in reducing n eurogenerative progression of Parkinson’s disease: A review. Int. J. Neurosci. 2011, 121, 51–57. [Google Scholar] [CrossRef] [PubMed]
- D’Mello, F.; Braidy, N.; Marçal, H.; Guillemin, G.; Rossi, F.; Chinian, M.; Laurent, D.; Teo, C.; Neilan, B.A. Cytotoxic Effects of Environmental Toxins on Human Glial Cells. Neurotox. Res. 2017, 31, 245–258. [Google Scholar] [CrossRef]
- Gao, A.; McCoy, H.M.; Zaman, V.; Shields, D.C.; Banik, N.L.; Haque, A. Calpain activation and progression of inflammatory cycles in Parkinson’s disease. Front. Biosci. 2022, 27, 20. [Google Scholar] [CrossRef]
- Shams, R.; Banik, N.L.; Haque, A. Calpain in the cleavage of alpha-synuclein and the pathogenesis of Par kinson’s disease. Prog. Mol. Biol. Transl. Sci. 2019, 167, 107–124. [Google Scholar]
- Donkor, I.O. An update on the therapeutic potential of calpain inhibitors: A patent review. Expert Opin. Ther. Pat. 2020, 30, 659–675. [Google Scholar] [CrossRef]
- Mo, M.; Xiao, Y.; Huang, S.; Cen, L.; Chen, X.; Zhang, L.; Luo, Q.; Li, S.; Yang, X.; Lin, X.; et al. MicroRNA expressing profiles in A53T mutant alpha-synuclein transgenic mice and Parkinsonian. Oncotarget 2017, 8, 15–28. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Su, Y.; Li, N.; Zhang, F.R.; Zhang, N. Induced Neuronal Injury by Regulating LINC00943/miR-142-5p/KPNA4/NF-κ B Pathway in SK-N-SH Cells. Neurochem. Res. 2021, 46, 3286–3300. [Google Scholar] [CrossRef]
- Chen, J.; Jiang, C.; Du, J.; Xie, C.L. MiR-142-5p Protects Against 6-OHDA-Induced SH-SY5Y Cell Injury by Down regulating BECN1 and Autophagy. Dose Response 2020, 18, 1559325820907016. [Google Scholar] [CrossRef] [PubMed]
- Ke, L.; Chen, Y.; Li, Y.; Chen, Z.; He, Y.; Liu, J.; Zhuang, Y. miR-142-5p promotes cervical cancer progression by targeting LMX1A thr ough Wnt/β-catenin pathway. Open Med. 2021, 16, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Majed, S.O.; Mustafa, S.A. MACE-Seq-based coding RNA and TrueQuant-based small RNA profile in bre ast cancer: Tumor-suppressive miRNA-1275 identified as a novel marker. BMC Cancer 2021, 21, 473. [Google Scholar] [CrossRef] [PubMed]
- Mai, J.; Gu, J.; Liu, Y.; Liu, X.; Sai, K.; Chen, Z.; Lu, W.; Yang, X.; Wang, J.; Guo, C.; et al. Negative regulation of miR-1275 by H3K27me3 is critical for glial indu ction of glioblastoma cells. Mol. Oncol. 2019, 13, 1589–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Lu, C.; Wang, H.; Lin, Q.; Cai, L.; Meng, F.; Tesfaye, E.B.; Lai, H.C.; Tzeng, C.M. Identification of hsa-miR-1275 as a novel biomarker targeting MECP2 for human epilepsy of unknown etiology. Mol. Ther. Methods Clin. Dev. 2020, 19, 398–410. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhao, X.; Wu, Z.; Ma, Y.; Li, H. LncRNA FLVCR1-AS1 mediates miR-23a-5p/SLC7A11 axis to promote malignan t behavior of cervical cancer cells. Bioengineered 2022, 13, 10454–10466. [Google Scholar] [CrossRef]
- Gao, X.; Cao, Y.; Li, J.; Wang, C.; He, H. LncRNA TPT1-AS1 Sponges miR-23a-5p in Glioblastoma to Promote Cancer C ell Proliferation. Cancer Biother. Radiopharm. 2021, 36, 549–555. [Google Scholar]
- Roncon, P.; Soukupovà, M.; Binaschi, A.; Falcicchia, C.; Zucchini, S.; Ferracin, M.; Langley, S.R.; Petretto, E.; Johnson, M.R.; Marucci, G.; et al. MicroRNA profiles in hippocampal granule cells and plasma of rats with pilocarpine-induced epilepsy–comparison with human epileptic samples. Sci. Rep. 2015, 5, 14143. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Lin, M.; Chen, J.; Pedrosa, E.; Hrabovsky, A.; Fourcade, H.M.; Zheng, D.; Lachman, H.M. MicroRNA Profiling of Neurons Generated Using Induced Pluripotent Stem Cells Derived from Patients with Schizophrenia and Schizoaffective Di sorder, and 22q11.2 Del. PLoS One 2015, 10, e0132387. [Google Scholar]
- Luo, M.; Hu, Z.; Kong, Y.; Li, L. MicroRNA-432-5p inhibits cell migration and invasion by targeting CXCL 5 in colorectal cancer. Exp. Ther. Med. 2021, 21, 301. [Google Scholar] [CrossRef]
- Xu, T.; Lei, T.; Li, S.Q.; Mai, E.H.; Ding, F.H.; Niu, B. DNAH17-AS1 promotes pancreatic carcinoma by increasing PPME1 expression via inhibition of miR-432-5p. World J. Gastroenterol. 2020, 26, 1745–1757. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Lu, T.; Li, X.; Jiang, M.; Jia, M.; Liu, J.; Zhang, D.; Li, J.; Wang, L. Altered Expression of Brain-specific Autism-Associated miRNAs in the H an Chinese Population. Front. Genet. 2022, 13, 865881. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, L.; Teuber-Hanselmann, S.; Soub, D.; Harnisch, K.; Mairinger, F.; Junker, A. MicroRNA profiles of MS gray matter lesions identify modulators of the synaptic protein synaptotagmin-7. Brain Pathol. 2020, 30, 524–540. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.J.; Ban, H.J.; Park, S.M.; Lee, B.; Choi, Y.; Baek, Y.; Lee, S.; Cha, S. Circulating microRNAs as Potential Diagnostic Biomarkers for Poor Slee p Quality. Nat. Sci. Sleep 2021, 13, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, P.M.M.; Vieira, E.; Lemos, D.S.; Souza, I.L.M.; Zanata, S.M.; Pankievicz, V.C.; Tuleski, T.R.; Souza, E.M.; Wowk, P.F.; Urban, C.A.; et al. Identification of miRNAs Enriched in Extracellular Vesicles Derived fr om Serum Samples of Breast Cancer Patients. Biomolecules 2020, 10, 150. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Zhang, X.; Ding, X.; Zheng, Y.; Du, H.; Li, H.; Ji, H.; Wang, Z.; Jiao, P.; Song, X.; et al. Long Noncoding RNA LINC00460 Promotes Cell Progression by Sponging miR -4443 in Head and Neck Squamous Cell Carcinoma. Cell Transplant. 2020, 29, 963689720927405. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Xu, J.; Lin, J.; Lin, R.; Chen, K.; Kong, J.; Shui, X. Long Noncoding RNA FEZF1-AS1 Promotes Osteosarcoma Progression by Regu lating the miR-4443/NUPR1 Axis. Oncol. Res. 2018, 26, 1335–1343. [Google Scholar] [CrossRef]
- Gong, J.; Wang, J.; Liu, T.; Hu, J.; Zheng, J. lncRNA FEZF1-AS1 contributes to cell proliferation, migration and inva sion by sponging miR-4443 in hepatocellular carcinoma. Mol. Med. Rep. 2018, 18, 5614–5620. [Google Scholar]
- Ge, X.; Yao, T.; Zhang, C.; Wang, Q.; Wang, X.; Xu, L.C. Human microRNA-4433 (hsa-miR-4443) Targets 18 Genes to be a Risk Facto r of Neurodegenerative Diseases. Curr. Alzheimer Res. 2022, 19, 511–522. [Google Scholar]
- Li, Z.; Yao, H.; Wang, S.; Li, G.; Gu, X. CircTADA2A suppresses the progression of colorectal cancer via miR-374 a-3p/KLF14 axis. J. Exp. Clin. Cancer Res. 2020, 39, 160. [Google Scholar] [CrossRef]
- Zhu, J.; Huang, Y.; Zhang, Y.; Huang, R.; Huang, C. KCNMB2-AS1 Promotes Bladder Cancer Progression Through Sponging miR-37 4a-3p to Upregulate S100A10. Front. Genet. 2021, 12, 655569. [Google Scholar] [CrossRef] [PubMed]
- Feng, B.; Meng, X.; Zhou, H.; Chen, L.; Zou, C.; Liang, L.; Meng, Y.; Xu, N.; Wang, H.; Zou, D. Identification of Dysregulated Mechanisms and Potential Biomarkers in Ischemic Stroke Onset. Int. J. Gen. Med. 2021, 14, 4731–4744. [Google Scholar] [CrossRef]
- Xiong, D.D.; Xu, W.Q.; He, R.Q.; Dang, Y.W.; Chen, G.; Luo, D.Z. In silico analysis identified miRNA-based therapeutic agents against g lioblastoma multiforme. Oncol. Rep. 2019, 41, 2194–2208. [Google Scholar] [PubMed] [Green Version]
- Wei, Q.; Guo, Z.; Chen, D.; Jia, X. MiR-542-3p Suppresses Neuroblastoma Cell Proliferation and Invasion by Downregulation of KDM1A and ZNF346. Open Life Sci. 2020, 15, 173–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, D.; Yang, S.; Wang, X. Long non-coding RNA SNHG16 regulates cell behaviors through miR-542-3p /HNF4α axis via RAS/RAF/MEK/ERK signaling pathway in pediatric neurobl astoma cells. Biosci. Rep. 2020, 40, BSR20200723. [Google Scholar] [CrossRef]
- Wen, Y.; Gong, X.; Dong, Y.; Tang, C. Long Non Coding RNA SNHG16 Facilitates Proliferation, Migration, Invas ion and Autophagy of Neuroblastoma Cells via Sponging miR-542-3p and U pregulating ATG5 Expression. OncoTargets Ther. 2020, 13, 263–275. [Google Scholar] [CrossRef] [Green Version]
- Althoff, K.; Lindner, S.; Odersky, A.; Mestdagh, P.; Beckers, A.; Karczewski, S.; Molenaar, J.J.; Bohrer, A.; Knauer, S.; Speleman, F.; et al. miR-542-3p exerts tumor suppressive functions in neuroblastoma by down regulating Survivin. Int. J. Cancer 2015, 136, 1308–1320. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Zhao, J.; Zhang, N.; Xu, X.; Li, R.; Yi, Y.; Fang, L.; Zhang, L.; Li, M.; Wu, J.; et al. MicroRNA-542-3p Suppresses Tumor Cell Invasion via Targeting AKT Pathw ay in Human Astrocytoma. J. Biol. Chem. 2015, 290, 24678–24688. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, A.; Sawada, K.; Nakamura, K.; Kinose, Y.; Nakatsuka, E.; Kobayashi, M.; Miyamoto, M.; Ishida, K.; Matsumoto, Y.; Kodama, M.; et al. Exosomal miR-99a-5p is elevated in sera of ovarian cancer patients and promotes cancer cell invasion by increasing fibronectin and vitronect in expression in neighboring peritoneal mesothelial cells. BMC Cancer 2018, 18, 1065. [Google Scholar] [CrossRef]
- Chen, Y.T.; Yao, J.N.; Qin, Y.T.; Hu, K.; Wu, F.; Fang, Y.Y. Biological role and clinical value of miR-99a-5p in head and neck squa mous cell carcinoma (HNSCC): A bioinformatics-based study. FEBS Open Bio 2018, 8, 1280–1298. [Google Scholar] [CrossRef]
- Wang, G.; Lu, Y.; Di, S.; Xie, M.; Jing, F.; Dai, X. miR-99a-5p inhibits glycolysis and induces cell apoptosis in cervical cancer by targeting RRAGD. Oncol. Lett. 2022, 24, 228. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.; Belmonte, K.C.; Kasten, T.; Bateman, R.; Kim, J. Intra- and Inter-individual Variability of microRNA Levels in Human Ce rebrospinal Fluid: Critical Implications for Biomarker Discovery. Sci. Rep. 2017, 7, 12720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Deng, Q.; Lv, Z.; Ling, Y.; Hou, X.; Chen, Z.; Dinglin, X.; Ma, S.; Li, D.; Wu, Y.; et al. N6-methyladenosine induced miR-143-3p promotes the brain metastasis of lung cancer via regulation of VASH1. Mol. Cancer 2019, 18, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, X.; Shao, Y.; Teng, Y.; Liu, S.; Li, W.; Xue, L.; Cao, Y.; Sun, C.; Zhang, J.; Han, J.; et al. The Cancer-Testis Long Non-coding RNA PCAT6 Facilitates the Malignant Phenotype of Ovarian Cancer by Sponging miR-143-3p. Front. Cell Dev. Biol. 2021, 9, 593677. [Google Scholar] [CrossRef]
- Wang, L.; Shui, X.; Mei, Y.; Xia, Y.; Lan, G.; Hu, L.; Zhang, M.; Gan, C.L.; Li, R.; Tian, Y.; et al. miR-143-3p Inhibits Aberrant Tau Phosphorylation and Amyloidogenic Pro cessing of APP by Directly Targeting DAPK1 in Alzheimer’s Disease. Int. J. Mol. Sci. 2022, 23, 7992. [Google Scholar] [CrossRef]
- Jiang, F.; Wu, Q.; Sun, S.; Bi, G.; Guo, L. Identification of potential diagnostic biomarkers for Parkinson’s dise ase. FEBS Open Bio 2019, 9, 1460–1468. [Google Scholar] [CrossRef] [Green Version]
- Salemi, M.; Lanza, G. A Transcriptome Analysis of mRNAs and Long Non-Coding RNAs in Patients with Parkinson’s Disease. Int. J. Mol. Sci. 2022, 23, 1535. [Google Scholar] [CrossRef]
- Li, C.S.; Chae, S.C.; Lee, J.H.; Zhang, Q.; Chung, H.T. Identification of single nucleotide polymorphisms in FOXJ1 and their association with allergic rhinitis. J. Hum. Genet. 2006, 51, 292–297. [Google Scholar] [CrossRef]
- Li, C.S.; Zhang, Q.; Lim, M.K.; Sheen, D.H.; Shim, S.C.; Kim, J.Y.; Lee, S.S.; Yun, K.J.; Moon, H.B.; Chung, H.T.; et al. Association of FOXJ1 polymorphisms with systemic lupus erythematosus and rheumatoid arthritis in Korean population. Exp. Mol. Med. 2007, 39, 805–811. [Google Scholar] [CrossRef]
- Sun, H.; Li, M.; Ji, Y.; Zhu, J.; Chen, Z.; Zhang, L.; Deng, C.; Cheng, Q.; Wang, W.; Shen, Y.; et al. Identification of Regulatory Factors and Prognostic Markers in Amyotro phic Lateral Sclerosis. Antioxidants 2022, 11, 303. [Google Scholar] [CrossRef]
- Weishaupt, C.; Mastrofrancesco, A.; Metze, D.; Kemper, B.; Stegemann, A.; Picardo, M.; Klein-Szanto, A.J.P.; Bohm, M. Paired Basic Amino Acid-cleaving Enzyme 4 (PCSK6): An Emerging New Target Molecule in Human Melanoma. Acta Derm.-Venereol. 2020, 100, adv00157. [Google Scholar] [CrossRef] [PubMed]
- Postuma, R.B.; Berg, D.; Stern, M.; Poewe, W.; Olanow, C.W.; Oertel, W.; Obeso, J.; Marek, K.; Litvan, I.; Lang, A.E.; et al. MDS clinical diagnostic criteria for Parkinson’s disease. Mov. Disord. Off. J. Mov. Disord. Soc. 2015, 30, 1591–1601. [Google Scholar] [CrossRef] [PubMed]
- Aparicio-Puerta, E.; Lebrón, R.; Rueda, A.; Gómez-Martín, C.; Giannoukakos, S.; Jaspez, D.; Medina, J.M.; Zubkovic, A.; Jurak, I.; Fromm, B.; et al. sRNAbench and sRNAtoolbox 2019: Intuitive fast small RNA profiling and differential expression. Nucleic Acids Res. 2019, 47, W530–W535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing da ta. Nucleic Acids Res. 2014, 42, 68–73. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Sticht, C.; De La Torre, C.; Parveen, A.; Gretz, N. miRWalk: An online resource for prediction of microRNA binding sites. PLoS ONE 2018, 13, e0206239. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level of Expression Found | miRNA | Association with PD Models or Patients (References) | Association with Other Neurological, Neuropsychiatric, or Neuro-Oncological Disorders (References) |
|---|---|---|---|
| Up | miR-1275 | No | Multiple sclerosis [PMID: 35273684]; Human epilepsy of unknown etiology [PMID: 33251277]; Atypical meningioma [PMID: 32426270]; Ischemic stroke [PMID: 31935511]; Glioblastoma [PMID: 31162799; PMID: 25129238; PMID: 22736761]; Neural tube defects [PMID: 22642222]; Atherosclerosis [PMID: 31935511; PMID: 31506771] |
| Up | miR-4433b-3p | No | Poor sleep quality [PMID: 34234603]; Prediction for stroke [PMID: 31136284] |
| Up | miR-23a-5p | No | Cerebral ischemia in mice [PMID: 35547763]; Glioblastoma [PMID: 32783743]; Multiple sclerosis [PMID: 32432792]; Atherosclerosis [PMID: 30227118]; Epilepsy [PMID: 26382856]; Schizophrenia and schizoaffective disorder [PMID: 26173148] |
| Up | miR-4443 | No | Glioblastoma [PMID: 35992881; PMID: 35300350; PMID: 29678219; PMID: 29643013]; Alzheimer’s disease and Huntington’s disease [PMID: 35929619]; Sporadic amyotrophic lateral sclerosis [PMID: 34776863]; Stroke-induced immunosuppression [PMID: 32337817] |
| Up | miR-432-5p | No | Autism [PMID: 35342389]; Glioma [PMID: 33915163; PMID: 33220929; PMID: 32629066; PMID: 31504797; PMID: 31246330]; Neuroblastoma [PMID: 33837793; PMID: 25762502]; Multiple sclerosis [PMID: 31663645; PMID: 29084979]; Schizophrenia [PMID: 31297041] |
| Down | miR-374a-3p | No | Ischemic stroke [PMID: 34456585] |
| Down | miR-542-3p | Yes [PMID: 27965467] | Neuroblastoma [PMID: 33987474; PMID: 32412051; PMID: 32021273; PMID: 25046253]; Glioma [PMID: 33922649]; Glioblastoma [PMID: 30816530]; Astrocytoma [PMID: 26286747]; Cerebral infarction prevention [PMID: 33407827]; Ischemic stroke [PMID: 27151415]; Epilepsy [PMID: 31702493]; Neointima formation [PMID: 26026397] |
| Down | miR-142-5p | Yes [PMID: 34427876] Yes [PMID: 32127787] | Spinal muscular atrophy [PMID: 35584175]; Intracranial germ cell tumors [PMID: 35171328]; Cognitive impairment [PMID: 34302879]; Epilepsy [PMID: 33041753; PMID: 32800995; PMID: 32439493; PMID: 24454901]; Alzheimer’s disease and other dementia [PMID: 32251633; PMID: 26981236]; Glioma [PMID: 32238705]; Multiple sclerosis [PMID: 30175165; PMID: 28302134; PMID: 28114622] |
| Down | miR-143-3p | No | Acute ischemia/stroke [PMID: 35571371; PMID: 35401659; PMID: 33038923; PMID: 28724745]; Alzheimer’s disease [PMID: 34775974; PMID: 32337953]; Rare muscular dystrophy [PMID: 35393236]; Epilepsy [PMID: 35055144]; Amyotrophic lateral sclerosis [PMID: 34454204]; Intracranial Aneurysmal Tissues [PMID: 34185228]; Atherosclerosis [PMID: 33603842]; Peripheral nerve tumors [PMID: 32076030]; Multiple Sclerosis [PMID: 28114622] |
| Down | miR-99a-5p | No | Cerebro-spinal fluid biomarkers [PMID: 28983117]; Ischemic stroke [PMID: 30276300]; Neurotoxicity to PM2.5 [PMID: 27539004] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salemi, M.; Marchese, G.; Lanza, G.; Cosentino, F.I.I.; Salluzzo, M.G.; Schillaci, F.A.; Ventola, G.M.; Cordella, A.; Ravo, M.; Ferri, R. Role and Dysregulation of miRNA in Patients with Parkinson’s Disease. Int. J. Mol. Sci. 2023, 24, 712. https://doi.org/10.3390/ijms24010712
Salemi M, Marchese G, Lanza G, Cosentino FII, Salluzzo MG, Schillaci FA, Ventola GM, Cordella A, Ravo M, Ferri R. Role and Dysregulation of miRNA in Patients with Parkinson’s Disease. International Journal of Molecular Sciences. 2023; 24(1):712. https://doi.org/10.3390/ijms24010712
Chicago/Turabian StyleSalemi, Michele, Giovanna Marchese, Giuseppe Lanza, Filomena I. I. Cosentino, Maria Grazia Salluzzo, Francesca A. Schillaci, Giovanna Maria Ventola, Angela Cordella, Maria Ravo, and Raffaele Ferri. 2023. "Role and Dysregulation of miRNA in Patients with Parkinson’s Disease" International Journal of Molecular Sciences 24, no. 1: 712. https://doi.org/10.3390/ijms24010712
APA StyleSalemi, M., Marchese, G., Lanza, G., Cosentino, F. I. I., Salluzzo, M. G., Schillaci, F. A., Ventola, G. M., Cordella, A., Ravo, M., & Ferri, R. (2023). Role and Dysregulation of miRNA in Patients with Parkinson’s Disease. International Journal of Molecular Sciences, 24(1), 712. https://doi.org/10.3390/ijms24010712