
Deletion of the foxO4 Gene Increases Hypoxia Tolerance in Zebrafish

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
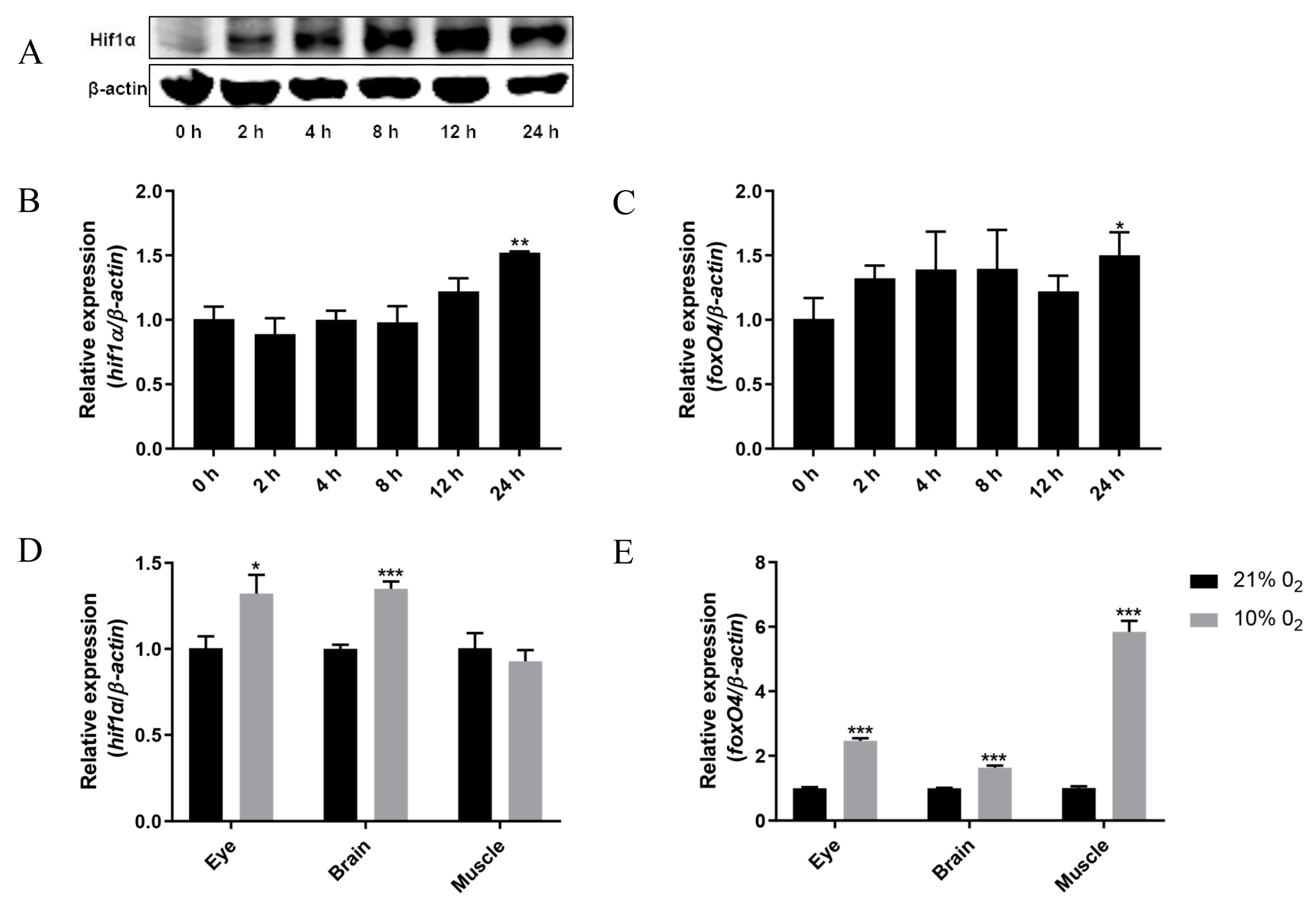
2.1. Hypoxia Leads to Up-Regulation of foxO4 mRNA Expression
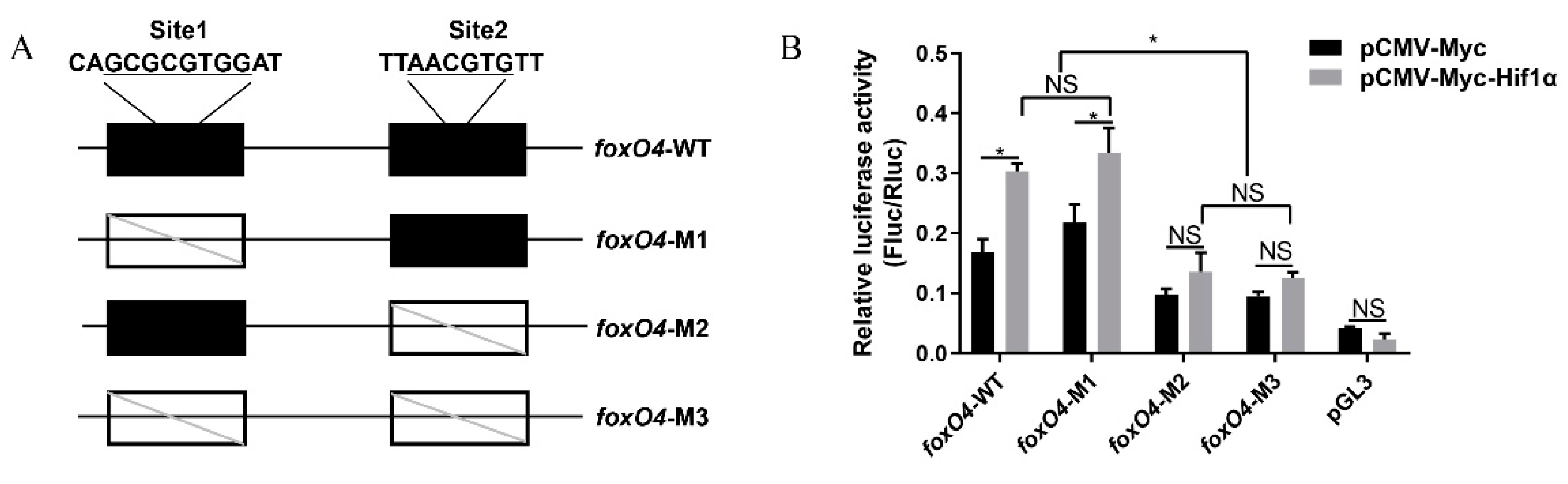
2.2. Hif1α Directly Targets the foxO4 Promoter under Hypoxia
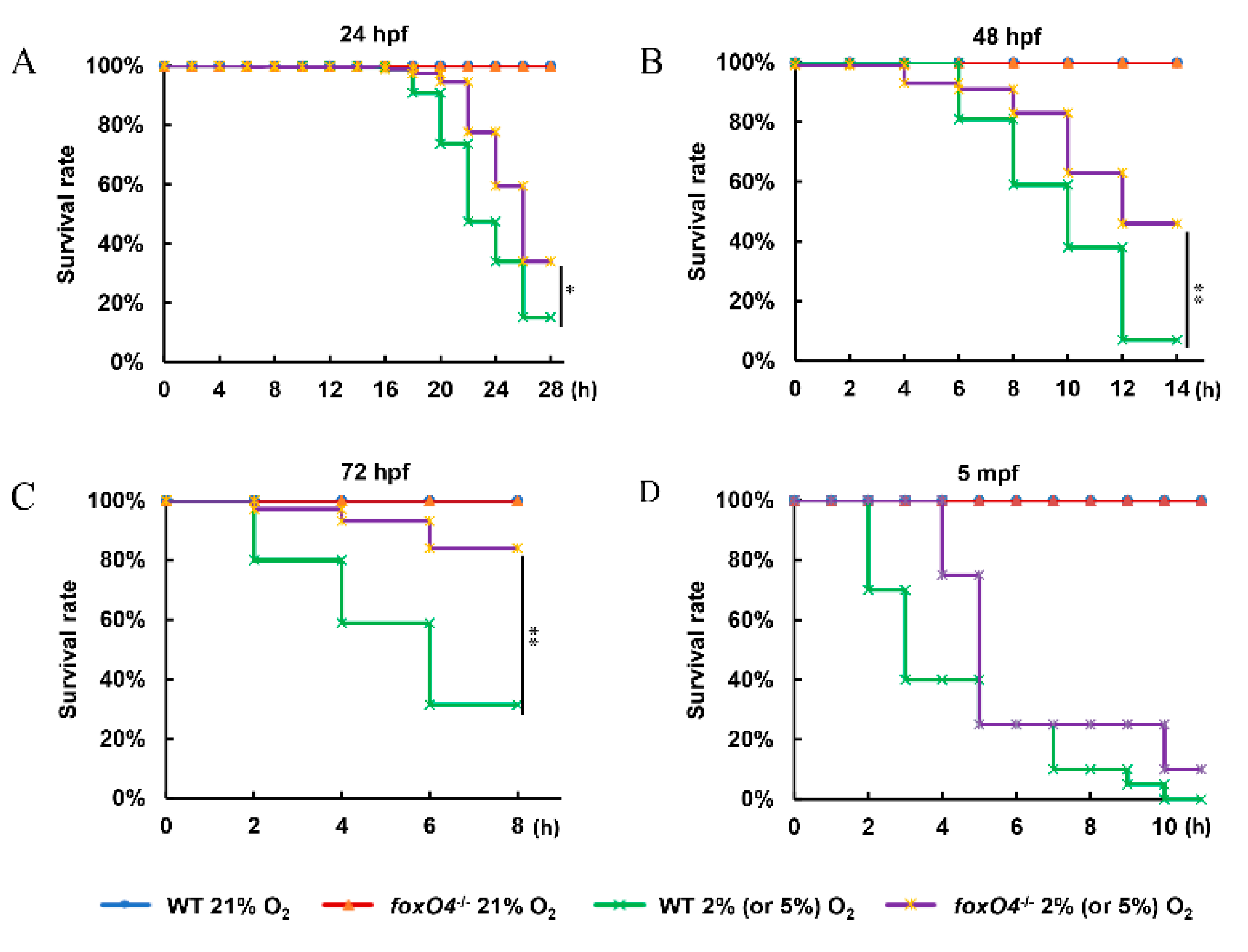
2.3. Disruption of Zebrafish foxO4 Increases Hypoxia Tolerance
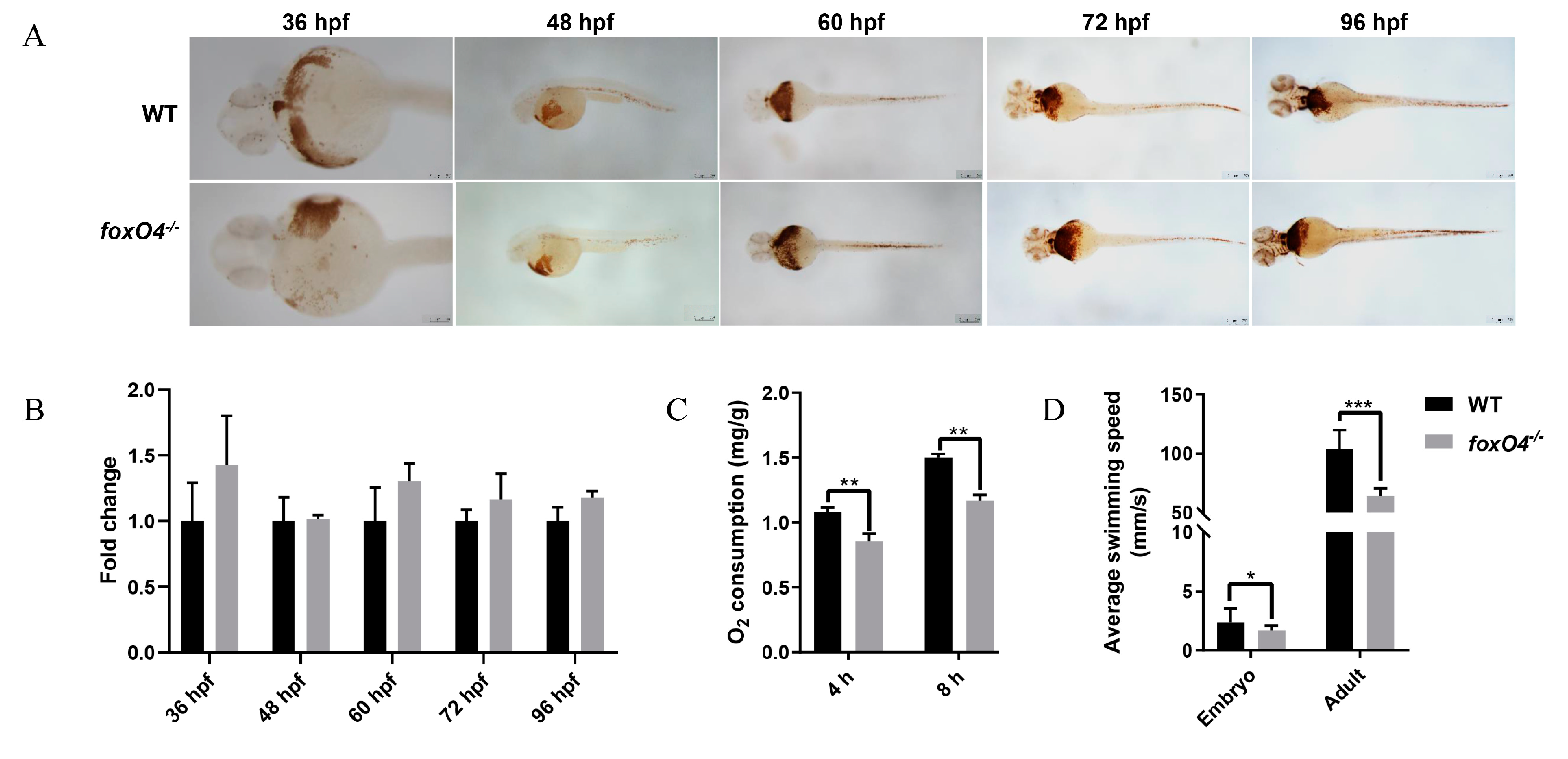
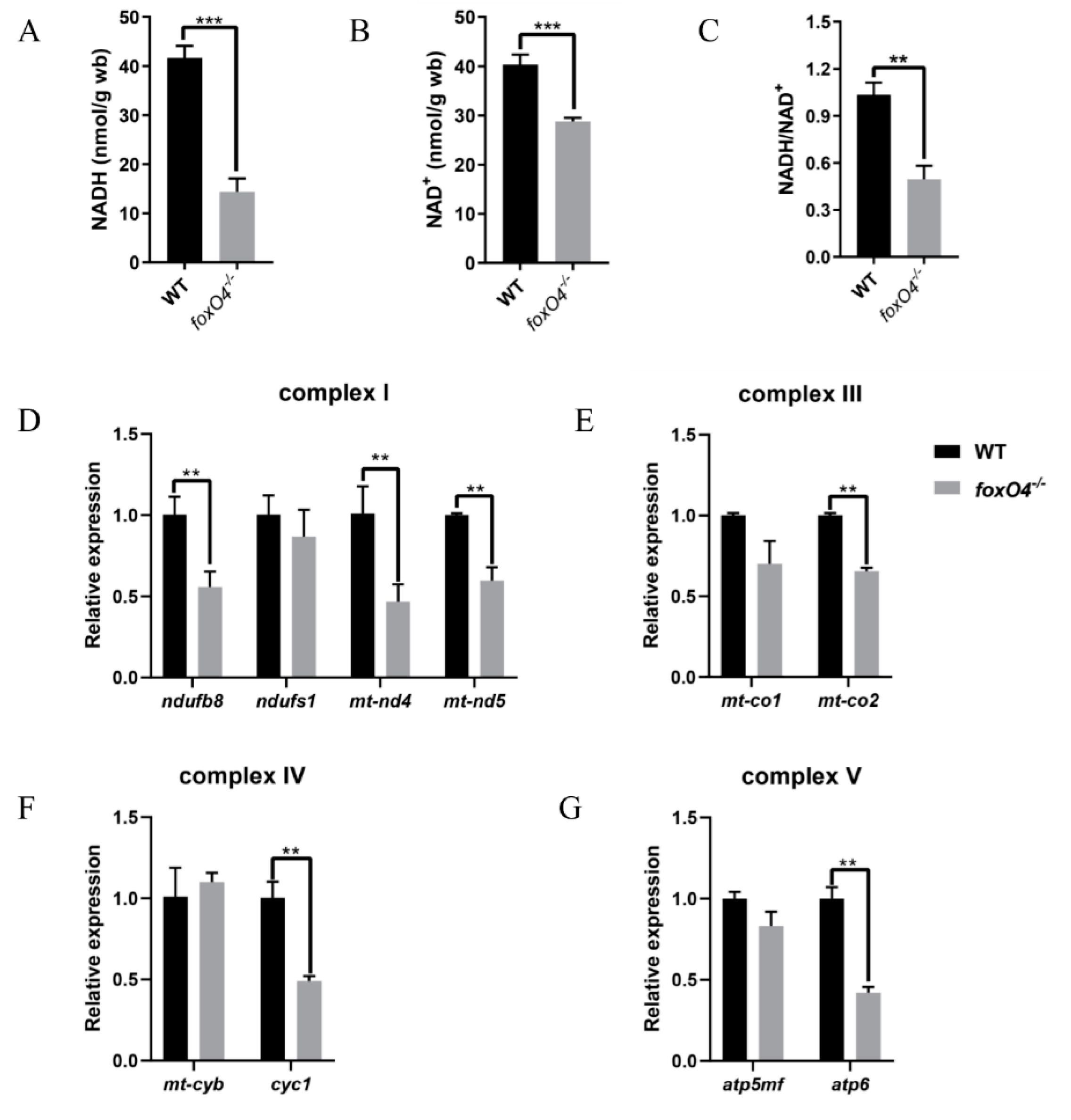
2.4. Disruption of Zebrafish foxO4 Reduces Oxygen Demand
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Hypoxia Treatment
4.3. Plasmid Constructs
4.4. Cell Transfection and Luciferase Reporter Assays
4.5. Generation of foxO4 Mutant Zebrafish
4.6. Oxygen Consumption and Locomotor Activity
4.7. O-Dianisidine Staining
4.8. NAD(H) Content
4.9. Quantitative Real-Time PCR (qRT-PCR)
4.10. Western Blotting
4.11. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zimna, A.; Kurpisz, M. Hypoxia-inducible factor-1 in physiological and pathophysiological angiogenesis: Applications and therapies. Biomed Res. Int. 2015, 2015, 549412. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.Z.; Jiang, S.; Zhang, L.; Yu, Z.B. Mitochondrial electron transport chain, ROS generation and uncoupling (Review). Int. J. Mol. Med. 2019, 44, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Loscalzo, J. The cellular response to hypoxia: Tuning the system with microRNAs. J. Clin. Investig. 2010, 120, 3815–3817. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.T.; Lasky, L.A. The forkhead transcription factor FOXO4 induces the down-regulation of hypoxia-inducible factor 1 alpha by a von Hippel-Lindau protein-independent mechanism. J. Biol. Chem. 2003, 278, 30125–30135. [Google Scholar] [CrossRef]
- Lee, J.W.; Bae, S.H.; Jeong, J.W.; Kim, S.H.; Kim, K.W. Ypoxia-inducible factor (HIF-1)alpha: Its protein stability and biological functions. Exp. Mol. Med. 2004, 36, 1–12. [Google Scholar] [CrossRef]
- Rademakers, S.E.; Span, P.N.; Kaanders, J.H.; Sweep, F.C.; van der Kogel, A.J.; Bussink, J. Molecular aspects of tumour hypoxia. Mol. Oncol. 2008, 2, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Cai, X.; Hu, B.; Mei, Z.; Zhang, D.; Ouyang, G.; Wang, J.; Zhang, W.; Xiao, W. Forkhead Transcription Factor 3a (FOXO3a) Modulates hypoxia signaling via up-regulation of the von hippel-lindau gene (VHL). J. Biol. Chem. 2016, 291, 25692–25705. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Pang, M.; Yu, X.; Wang, J.; Tong, J. Transcriptome sequencing provides insights into the mechanism of hypoxia adaption in bighead carp (Hypophthalmichthys nobilis). Comp. Biochem. Physiol. Part D Genom. Proteom. 2021, 40, 100891. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, F.; Zou, G.; Feng, C.; Sha, H.; Liu, S.; Liang, H. Physiological responses and molecular strategies in heart of silver carp (Hypophthalmichthys molitrix) under hypoxia and reoxygenation. Comp. Biochem. Physiol. Part D Genom. Proteom. 2021, 40, 100908. [Google Scholar] [CrossRef]
- Mendenhall, A.R.; LaRue, B.; Padilla, P.A. Glyceraldehyde-3-phosphate dehydrogenase mediates anoxia response and survival in Caenorhabditis elegans. Genetics 2006, 174, 1173–1187. [Google Scholar] [CrossRef]
- Liu, W.; Li, Y.; Luo, B. Current perspective on the regulation of FOXO4 and its role in disease progression. Cell. Mol. Life Sci. 2020, 77, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Yuan, Z.; Zhou, T.; Yang, Y.; Gao, D.; Dunham, R.; Liu, Z. FOXO genes in channel catfish and their response after bacterial infection. Dev. Comp. Immunol. 2019, 97, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.X.; Huang, Y.; Duan, X.K.; Zhang, M.; Tu, J.P.; Liu, J.X.; Liu, H.; Chen, T.S.; Wang, W.M.; Wang, H.L. Zebrafish miR-462-731 regulates hematopoietic specification and pu.1-dependent primitive myelopoiesis. Cell Death Differ. 2019, 26, 1531–1544. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.T.; Xie, J.T.; Xu, G.F.; Su, Y.Y.; Li, J.Z.; Zhu, M.; Wang, M.D. Hypoxia regulates reactive oxygen species levels in SHG-44 glioma cells. Neural Regen Res. 2013, 8, 554–560. [Google Scholar]
- Chen, C.; Mao, W.F.; Wu, Y.F. The Effects of hypoxia-reoxygenation in mouse digital flexor tendon-derived cells. Oxid Med. Cell. Longev. 2020, 2020, 7305392. [Google Scholar] [CrossRef]
- Jiang, S.; Yang, Z.; Di, S.; Hu, W.; Ma, Z.; Chen, F.; Yang, Y. Novel role of forkhead box O 4 transcription factor in cancer: Bringing out the good or the bad. Semin. Cancer Biol. 2018, 50, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, L.; Xu, X.; Han, P.; Wu, J.; Tian, X.; Li, M. FOXO4 Inhibits the Migration and Metastasis of Colorectal Cancer by Regulating the APC2/beta-Catenin Axis. Front Cell Dev. Biol. 2021, 9, 659731. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.H.; Jiang, Z.H.; Yang, H.M.; Zhang, Y.; Xu, L.H. Hypoxia-induced FOXO4/LDHA axis modulates gastric cancer cell glycolysis and progression. Clin. Transl. Med. 2021, 11, e279. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Jiang, X.; Chehab, F.F. FoxO4 interacts with the sterol regulatory factor SREBP2 and the hypoxia inducible factor HIF2alpha at the CYP51 promoter. J. Lipid Res. 2014, 55, 431–442. [Google Scholar] [CrossRef]
- Tothova, Z.; Kollipara, R.; Huntly, B.J.; Lee, B.H.; Castrillon, D.H.; Cullen, D.E.; McDowell, E.P.; Lazo-Kallanian, S.; Williams, I.R.; Sears, C.; et al. FoxOs are critical mediators of hematopoietic stem cell resistance to physiologic oxidative stress. Cell 2007, 128, 325–339. [Google Scholar] [CrossRef]
- Paik, J.H.; Ding, Z.; Narurkar, R.; Ramkissoon, S.; Muller, F.; Kamoun, W.S.; Chae, S.S.; Zheng, H.; Ying, H.; Mahoney, J. FoxOs cooperatively regulate diverse pathways governing neural stem cell homeostasis. Cell Stem Cell 2009, 5, 540–553. [Google Scholar] [CrossRef]
- Kim-Muller, J.Y.; Zhao, S.; Srivastava, S.; Mugabo, Y.; Noh, H.L.; Kim, Y.R.; Madiraju, S.R.; Ferrante, A.W.; Skolnik, E.Y.; Prentki, M. Metabolic inflexibility impairs insulin secretion and results in MODY-like diabetes in triple FoxO-deficient mice. Cell Metab. 2014, 20, 593–602. [Google Scholar] [CrossRef]
- Tsuchiya, K.; Tanaka, J.; Shuiqing, Y.; Welch, C.L.; DePinho, R.A.; Tabas, I.; Tall, A.R.; Goldberg, I.J.; Accili, D. FoxOs integrate pleiotropic actions of insulin in vascular endothelium to protect mice from atherosclerosis. Cell Metab. 2012, 15, 372–381. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, K.; Westerterp, M.; Murphy, A.J.; Subramanian, V.; Ferrante, A.W.; Tall, A.R.; Accili, D. Expanded granulocyte/monocyte compartment in myeloid-specific triple FoxO knockout increases oxidative stress and accelerates atherosclerosis in mice. Circ. Res. 2013, 112, 992–1003. [Google Scholar] [CrossRef]
- Hosaka, T.; Biggs, W.H.; Tieu, D.; Boyer, A.D.; Varki, N.M.; Cavenee, W.K.; Arden, K.C. Disruption of forkhead transcription factor (FOXO) family members in mice reveals their functional diversification. Proc. Natl. Acad. Sci. USA 2004, 101, 2975–2980. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.T.; Dowbenko, D.; Jackson, A.; Toney, L.; Lewin, D.A.; Dent, A.L.; Lasky, L.A. The forkhead transcription factor AFX activates apoptosis by induction of the BCL-6 transcriptional repressor. J. Biol. Chem. 2002, 277, 14255–14265. [Google Scholar] [CrossRef]
- Huang, J.; Liu, Y.; Wang, M.; Wang, R.; Ling, H.; Yang, Y. FoxO4 negatively modulates USP10 transcription to aggravate the apoptosis and oxidative stress of hypoxia/reoxygenation-induced cardiomyocytes by regulating the Hippo/YAP pathway. J. Bioenerg. Biomembr. 2021, 53, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Zhang, W.; Huang, C.; Liang, Q.; Bao, H.; Gong, Z.; Xu, M.; Wang, Z.; Wen, M.; Cheng, X. FoxO4 promotes myocardial ischemia-reperfusion injury: The role of oxidative stress-induced apoptosis. Am. J. Transl. Res. 2018, 10, 2890–2900. [Google Scholar]
- Liu, H.; Wang, L.; Weng, X.; Chen, H.; Du, Y.; Diao, C.; Chen, Z.; Liu, X. Inhibition of Brd4 alleviates renal ischemia/reperfusion injury-induced apoptosis and endoplasmic reticulum stress by blocking FoxO4-mediated oxidative stress. Redox Biol. 2019, 24, 101195. [Google Scholar] [CrossRef]
- Storey, K.B. Anoxia tolerance in turtles: Metabolic regulation and gene expression. Comp. Biochem. Physiol. A Mol Integr. Physiol. 2007, 147, 263–276. [Google Scholar] [CrossRef]
- Xiao, W.; Loscalzo, J. Metabolic responses to reductive stress. Antioxid. Redox Signal. 2020, 32, 1330–1347. [Google Scholar] [CrossRef]
- Huang, C.X.; Chen, N.; Wu, X.J.; Huang, C.H.; He, Y.; Tang, R.; Wang, W.M.; Wang, H.L. The zebrafish miR-462/miR-731 cluster is induced under hypoxic stress via hypoxia-inducible factor 1alpha and functions in cellular adaptations. FASEB J. 2015, 29, 4901–4913. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Wang, J.; Zhou, C.; Xiao, W. Zebrafish akt2 is essential for survival, growth, bone development, and glucose homeostasis. Mech. Dev. 2017, 143, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, D.; Du, J.; Zhou, C.; Li, Z.; Liu, X.; Ouyang, G.; Xiao, W. Tet1 facilitates hypoxia tolerance by stabilizing the HIF-alpha proteins independent of its methylcytosine dioxygenase activity. Nucleic. Acids Res. 2017, 45, 12700–12714. [Google Scholar] [CrossRef]
- Detrich, H.W.; Kieran, M.W.; Chan, F.Y.; Barone, L.M.; Yee, K.; Rundstadler, J.A.; Pratt, S.; Ransom, D.; Zon, L.I. Intraembryonic hematopoietic cell migration during vertebrate development. Proc. Natl. Acad. Sci. USA. 1995, 92, 10713–10717. [Google Scholar] [CrossRef]
- Shi, L.L.; Han, S.X.; Fei, J.M.; Zhang, L.; Ray, J.W.; Wang, W.M.; Li, Y.H. Molecular characterization and functional study of insulin-like androgenic gland hormone gene in the red swamp crayfish, Procambarus clarkia. Genes 2019, 10, 645. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, L.; Zhang, A.; Liu, H.; Wang, H. Deletion of the foxO4 Gene Increases Hypoxia Tolerance in Zebrafish. Int. J. Mol. Sci. 2023, 24, 8942. https://doi.org/10.3390/ijms24108942
Shi L, Zhang A, Liu H, Wang H. Deletion of the foxO4 Gene Increases Hypoxia Tolerance in Zebrafish. International Journal of Molecular Sciences. 2023; 24(10):8942. https://doi.org/10.3390/ijms24108942
Chicago/Turabian StyleShi, Linlin, Axin Zhang, Hong Liu, and Huanling Wang. 2023. "Deletion of the foxO4 Gene Increases Hypoxia Tolerance in Zebrafish" International Journal of Molecular Sciences 24, no. 10: 8942. https://doi.org/10.3390/ijms24108942