
Identification and Functional Characterization of MdNRT1.1 in Nitrogen Utilization and Abiotic Stress Tolerance in Malus domestica


Abstract
1. Introduction
2. Results
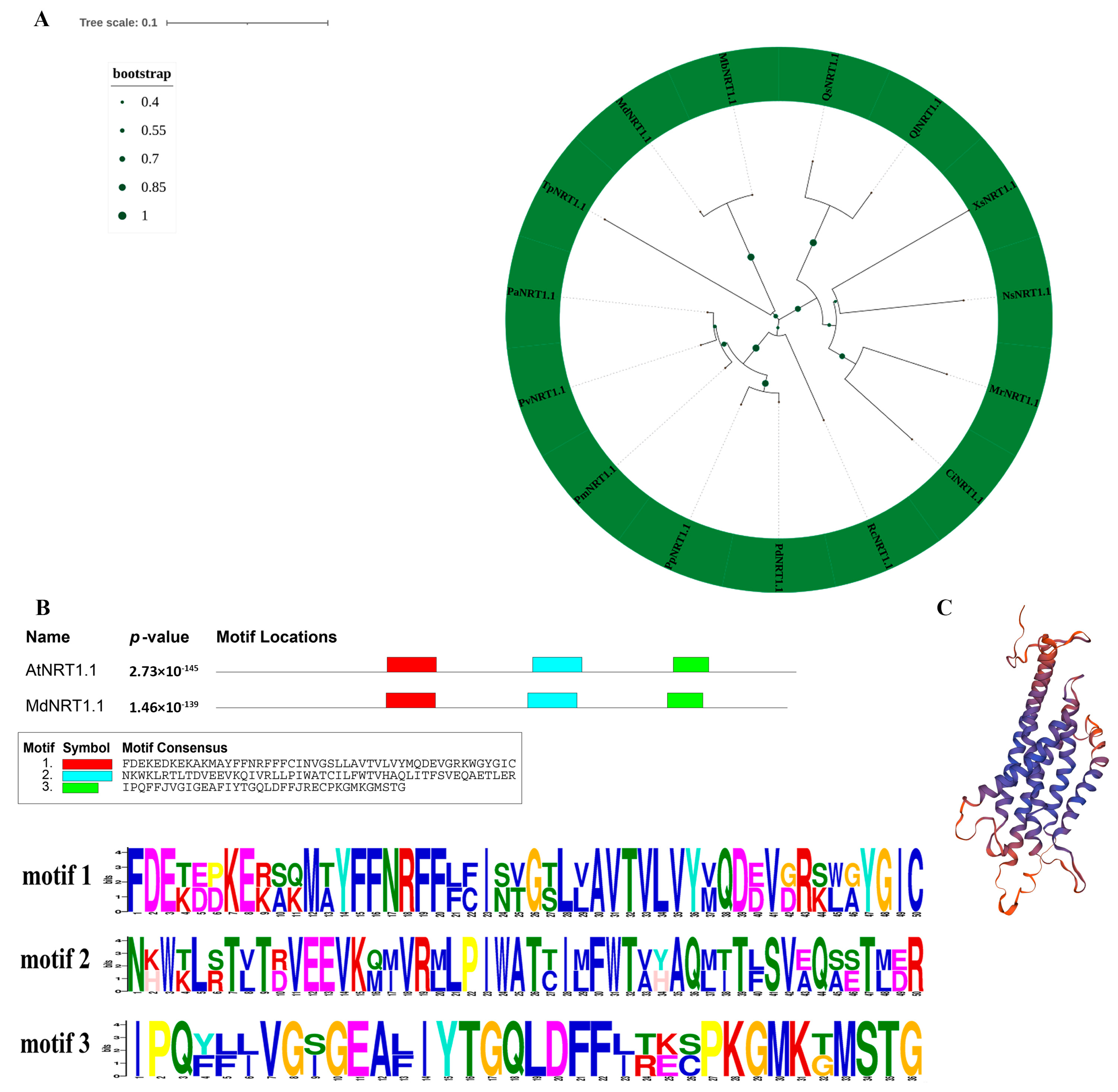
2.1. Phylogenetic Relationships and Multiple Sequence Alignment of MdNRT1.1
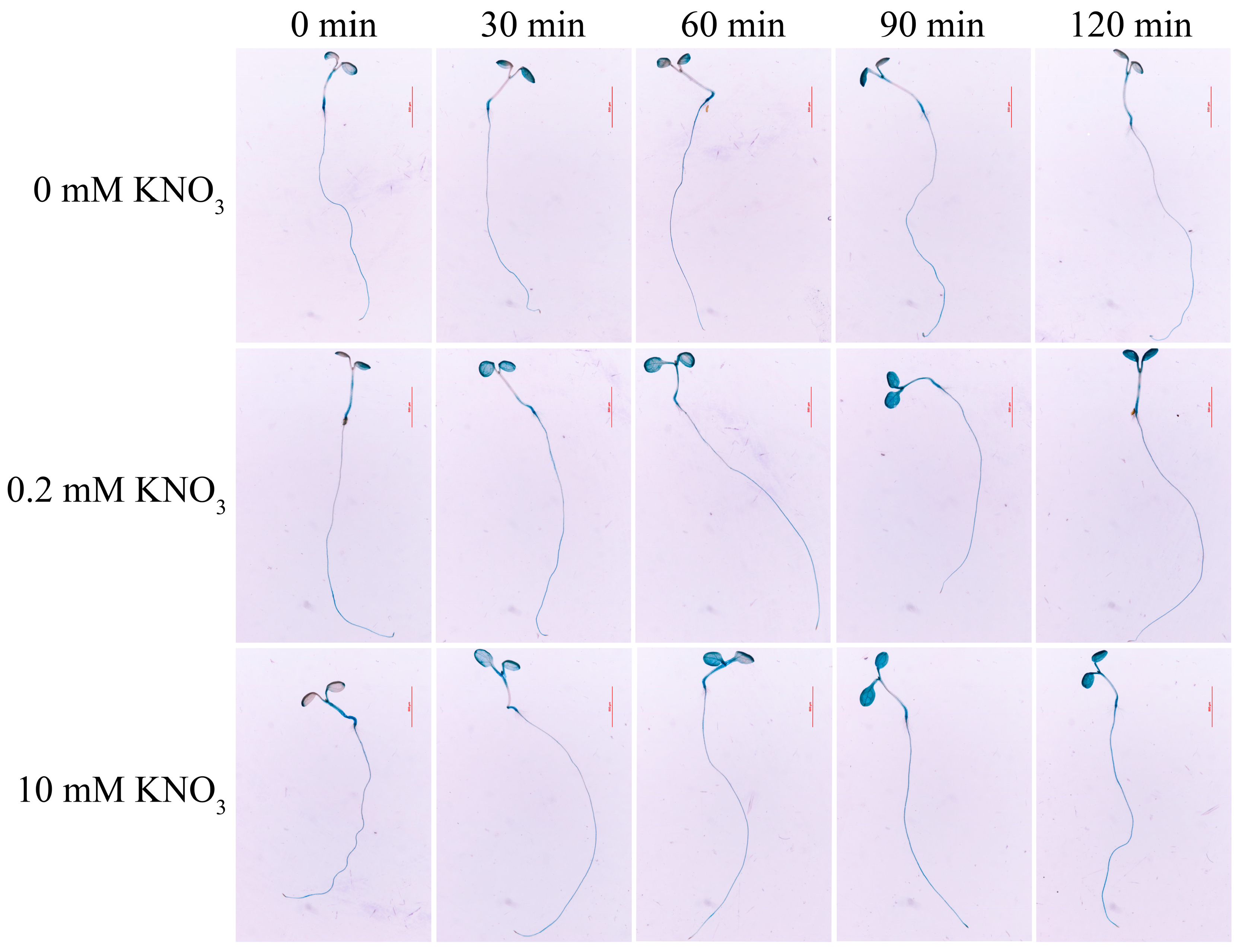
2.2. Nitrate and Abiotic Stresses Induce an Increased Transcript Level of MdNRT1.1
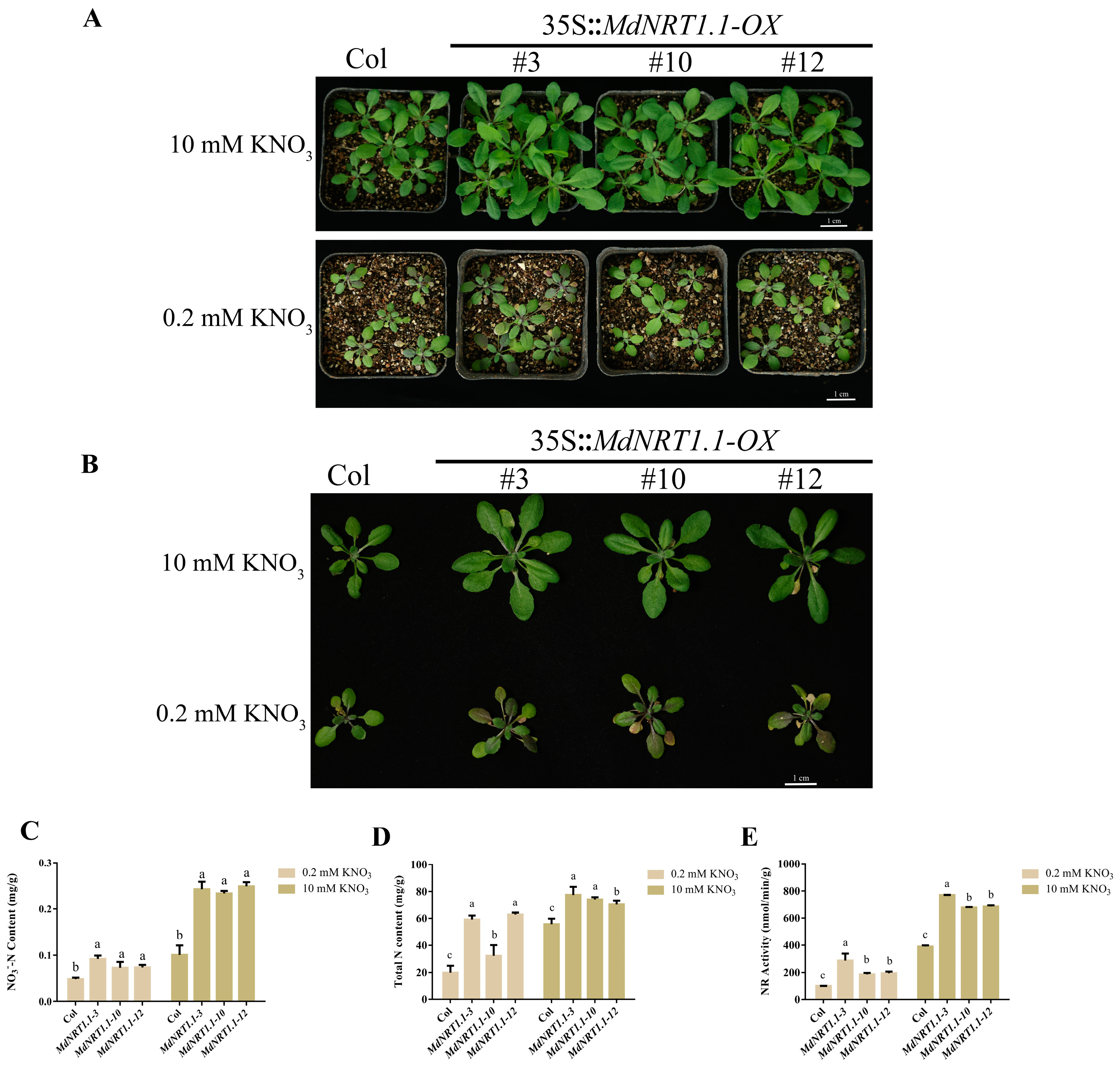
2.3. Overexpression of MdNRT1.1 Affects Nitrate Utilization
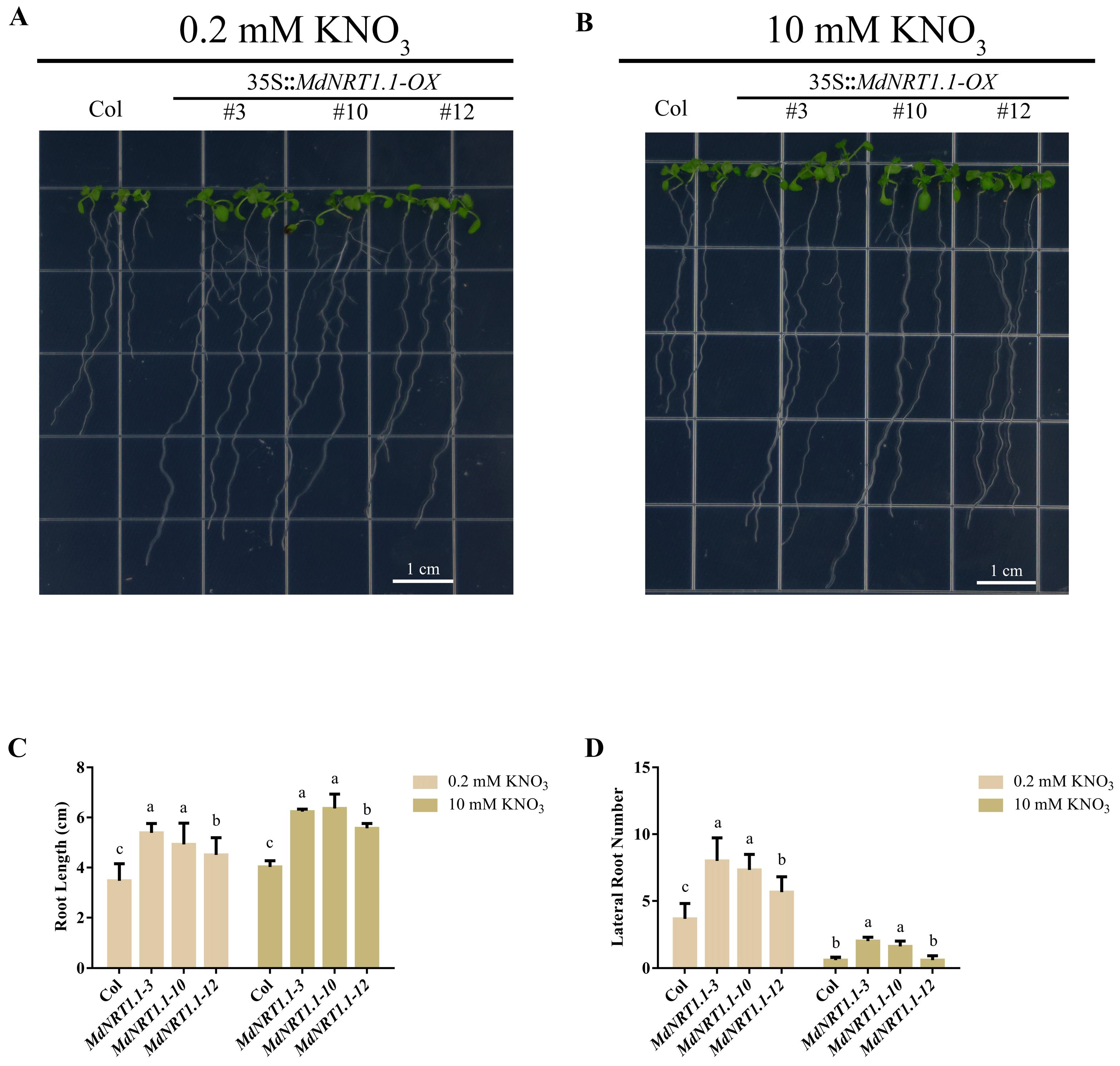
2.4. Overexpression of MdNRT1.1 Affects Root Development
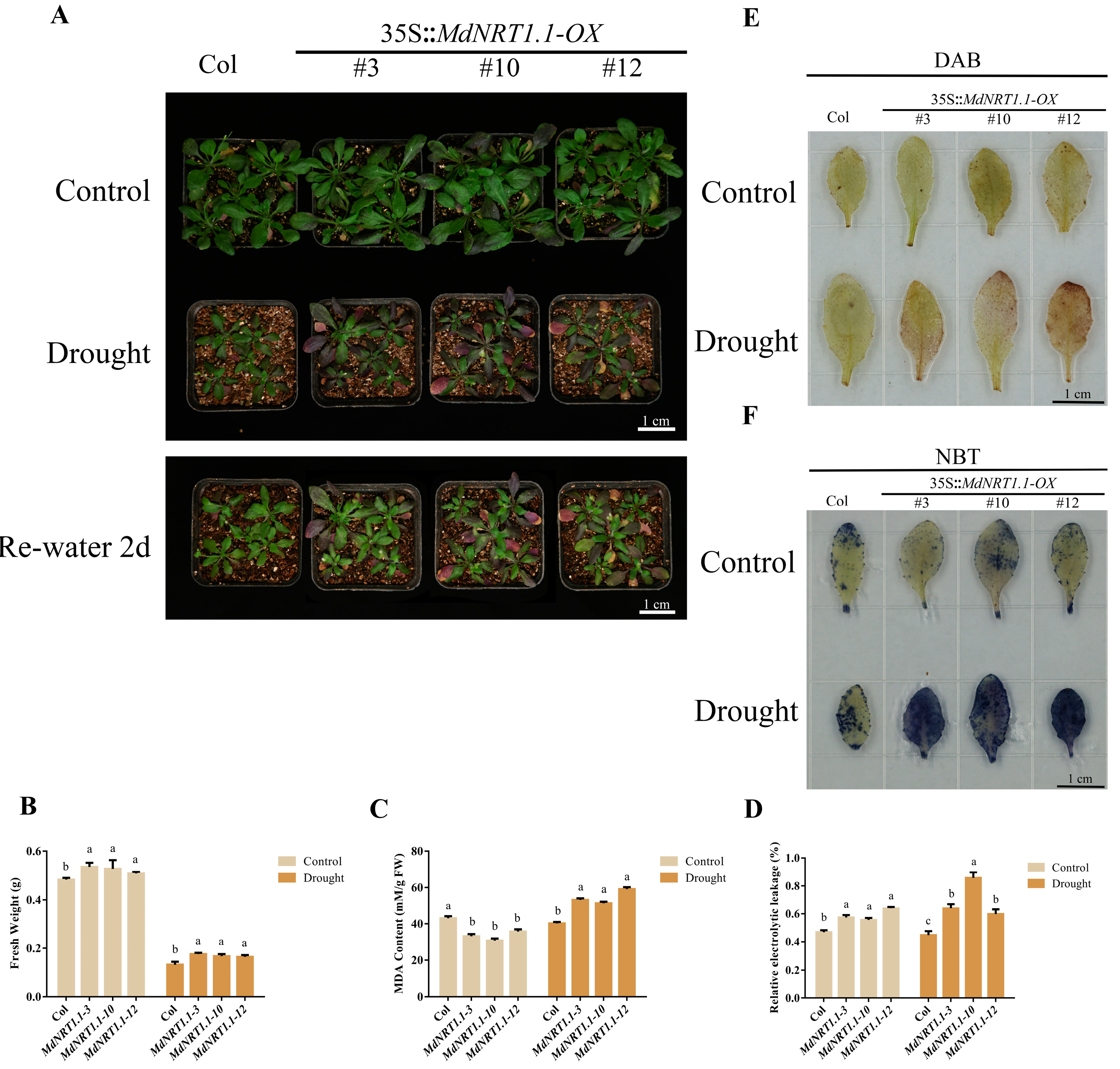
2.5. Overexpression of MdNRT1.1 Reduces Drought Tolerance
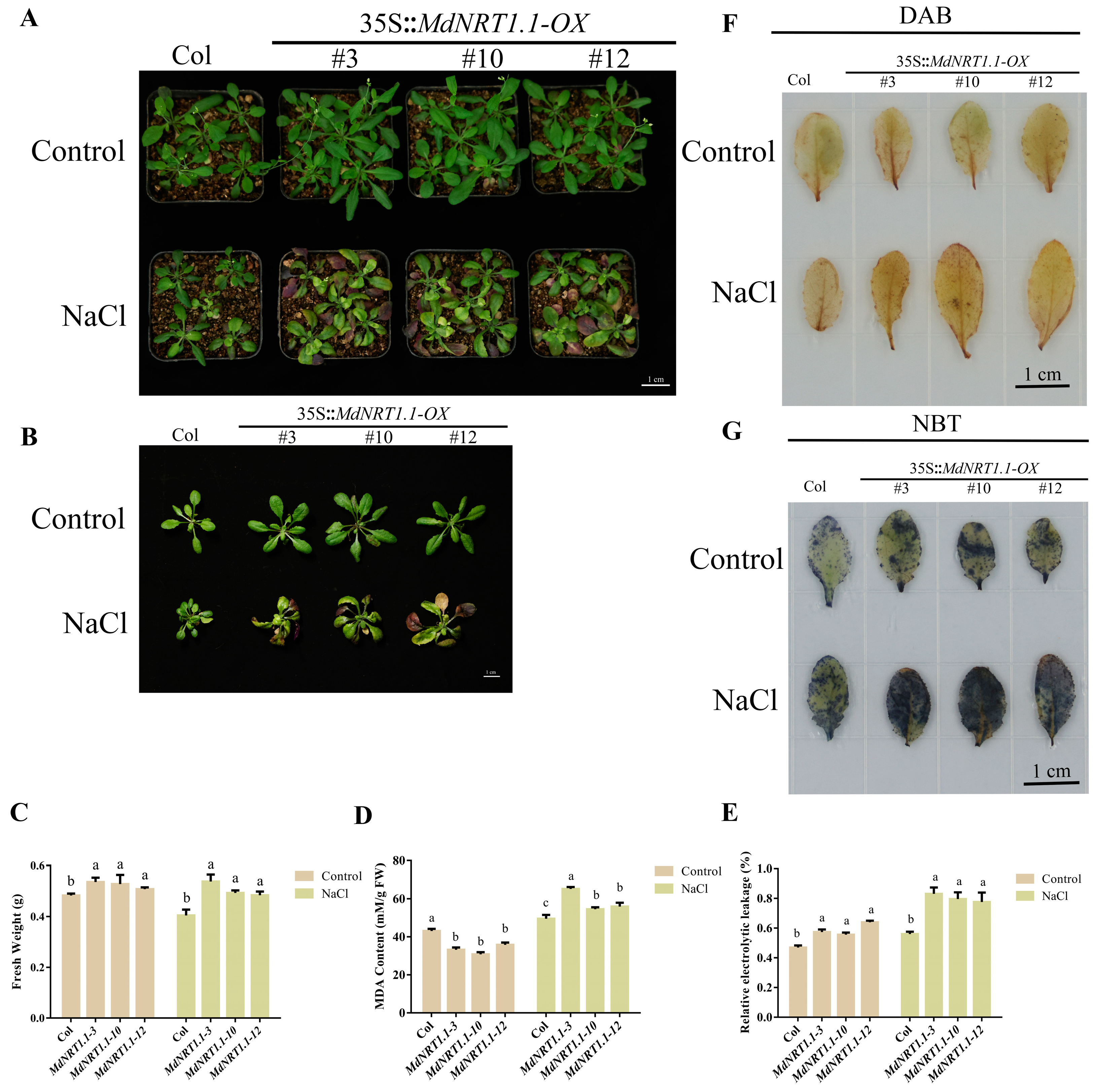
2.6. Overexpression of MdNRT1.1 Reduces Salt Stress Tolerance
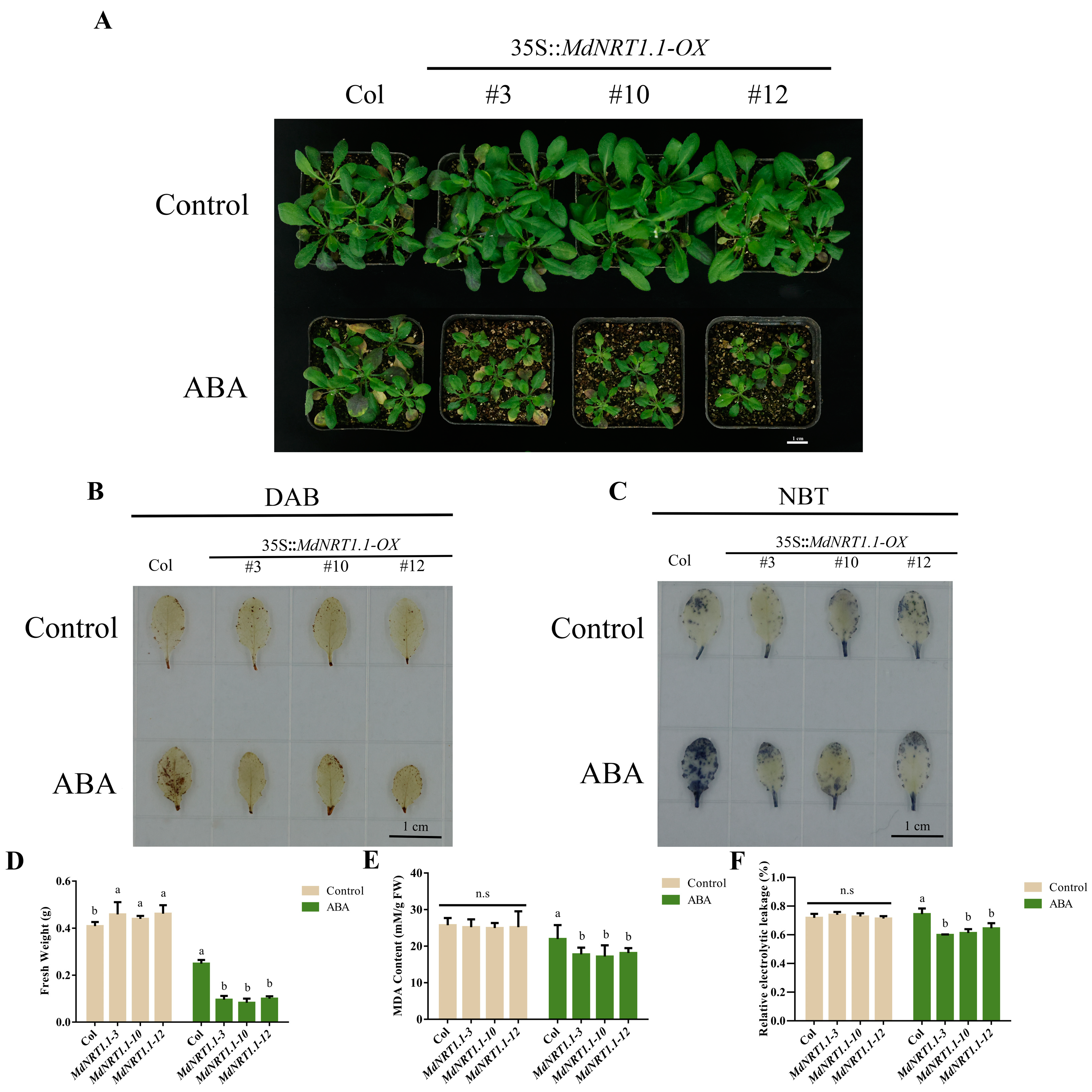
2.7. Overexpression of MdNRT1.1 Changes Response to Exogenous ABA
3. Discussion
4. Materials and Methods
4.1. Bioinformatics Analysis of the NRT1.1 Gene
4.2. Plant Materials and Cultivation Methods
4.3. Nitrate and Abiotic Stress Treatments
4.4. Generation of Transgenic Materials
4.5. GUS Staining
4.6. Extracting Plant Genomic DNA and RNA
4.7. Real-Time Quantitative Polymerase Chain Reaction (qPCR)
4.8. Determination of Nitrate, Nitrate Reductase, Total Nitrogen, Electrolyte Leakage, MDA, and O2−
4.9. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Orsel, M.; Filleur, S.; Fraisier, V.; Daniel-Vedele, F. Nitrate transport in plants: Which gene and which control? J. Exp. Bot. 2002, 53, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Mosier, A.R.; Syers, J.K.; Freney, J.R. Global assessment of nitrogen fertilizer: The SCOPE/IGBP nitrogen fertilizer rapid assessment project. Sci. China C Life Sci. 2005, 48, 759–766. [Google Scholar] [PubMed]
- Dluzniewska, P.; Gessler, A.; Kopriva, S.; Strnad, M.; Novak, O.; Dietrich, H.; Rennenberg, H. Exogenous supply of glutamine and active cytokinin to the roots reduces NO3- uptake rates in poplar. Plant Cell Environ. 2006, 29, 1284–1297. [Google Scholar] [CrossRef] [PubMed]
- Sakuraba, Y.; Chaganzhana, A.; Mabuchi, A.; Iba, K.; Yanagisawa, S. Enhanced NRT1.1/NPF6.3 expression in shoots improves growth under nitrogen deficiency stress in Arabidopsis. Commun. Biol. 2021, 4, 256. [Google Scholar] [CrossRef] [PubMed]
- Maghiaoui, A.; Gojon, A.; Bach, L. NRT1.1-centered nitrate signaling in plants. J. Exp. Bot. 2020, 71, 6226–6237. [Google Scholar] [CrossRef]
- Weil, S.; Barker, A.V.; Zandvakili, O.R.; Etemadi, F. Plant growth and calcium and potassium accumulation in lettuce under different nitrogen regimes of ammonium and nitrate nutrition. J. Plant Nutr. 2021, 44, 270–281. [Google Scholar] [CrossRef]
- de Souza, J.C.; Monteiro, F.A.; Xin, X.P.; He, Z.L. Nitrate Supply Affects Copper Nanoparticle Accumulation by Maize Plant and Availability of Nutrients in Rhizosphere and Bulk Soil. J. Soil Sci. Plant Nutr. 2022, 22, 3598–3610. [Google Scholar] [CrossRef]
- Kishchenko, O.; Stepanenko, A.; Straub, T.; Zhou, Y.; Neuhauser, B.; Borisjuk, N. Ammonium Uptake, Mediated by Ammonium Transporters, Mitigates Manganese Toxicity in Duckweed, Spirodela polyrhiza. Plants 2023, 12, 205. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Cheng, Y.H.; Chen, K.E.; Tsay, Y.F. Nitrate Transport, Signaling, and Use Efficiency. Annu. Rev. Plant Biol. 2018, 69, 85–122. [Google Scholar] [CrossRef]
- Kim, Y.J.; Choi, W.J.; Lim, S.S.; Kwak, J.H.; Chang, S.X.; Kim, H.Y.; Yoon, K.S.; Ro, H.M. Changes in nitrogen isotopic compositions during composting of cattle feedlot manure: Effects of bedding material type. Bioresour. Technol. 2008, 99, 5452–5458. [Google Scholar] [CrossRef]
- Bae, B.; Kim, M.; Kim, S.; Ro, H.S. Growth Characteristics of Polyporales Mushrooms for the Mycelial Mat Formation. Mycobiology 2021, 49, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, A.; Santini, G.; Piombino, P.; Pittari, E.; Sanmartin, C.; Moio, L.; Modesti, M.; Bellincontro, A.; Mencarelli, F. Nitrogen maceration of wine grape: An alternative and sustainable technique to carbonic maceration. Food Chem. 2023, 404, 134138. [Google Scholar] [CrossRef] [PubMed]
- Gorji, Z.E.; Khodadadi, A.A.; Riahi, S.; Repo, T.; Mortazavi, Y.; Kemell, M. Functionalization of nitrogen-doped graphene quantum dot: A sustainable carbon-based catalyst for the production of cyclic carbonate from epoxide and CO2. J. Environ. Sci. 2023, 126, 408–422. [Google Scholar] [CrossRef] [PubMed]
- James, S.N.; Vijayanandan, A. Recent advances in simultaneous nitrification and denitrification for nitrogen and micropollutant removal: A review. Biodegradation 2023, 34, 103–123. [Google Scholar] [CrossRef] [PubMed]
- Mahboob, W.; Yang, G.Z.; Irfan, M. Crop nitrogen (N) utilization mechanism and strategies to improve N use efficiency. Acta Physiol. Plant 2023, 45, 52. [Google Scholar] [CrossRef]
- Javed, T.; Indo, I.; Singhal, R.K.; Shabbir, R.; Shah, A.N.; Kumar, P.; Jinger, D.; Dharmappa, P.M.; Shad, M.A.; Saha, D.; et al. Recent Advances in Agronomic and Physio-Molecular Approaches for Improving Nitrogen Use Efficiency in Crop Plants. Front. Plant Sci. 2022, 13, 877544. [Google Scholar] [CrossRef]
- Medici, A.; Szponarski, W.; Dangeville, P.; Safi, A.; Dissanayake, I.M.; Saenchai, C.; Emanuel, A.; Rubio, V.; Lacombe, B.; Ruffel, S.; et al. Identification of Molecular Integrators Shows that Nitrogen Actively Controls the Phosphate Starvation Response in Plants. Plant Cell 2019, 31, 1171–1184. [Google Scholar] [CrossRef]
- Chai, S.; Li, E.; Zhang, Y.; Li, S. NRT1.1-Mediated Nitrate Suppression of Root Coiling Relies on PIN2- and AUX1-Mediated Auxin Transport. Front. Plant Sci. 2020, 11, 671. [Google Scholar] [CrossRef]
- Huang, N.C.; Chiang, C.S.; Crawford, N.M.; Tsay, Y.F. CHL1 encodes a component of the low-affinity nitrate uptake system in Arabidopsis and shows cell type-specific expression in roots. Plant Cell 1996, 8, 2183–2191. [Google Scholar]
- Kiba, T.; Krapp, A. Plant Nitrogen Acquisition Under Low Availability: Regulation of Uptake and Root Architecture. Plant Cell Physiol. 2016, 57, 707–714. [Google Scholar] [CrossRef]
- Chen, H.Y.; Lin, S.H.; Cheng, L.H.; Wu, J.J.; Lin, Y.C.; Tsay, Y.F. Potential transceptor AtNRT1.13 modulates shoot architecture and flowering time in a nitrate-dependent manner. Plant Cell 2021, 33, 1492–1505. [Google Scholar] [CrossRef] [PubMed]
- Mantelin, S.; Desbrosses, G.; Larcher, M.; Tranbarger, T.J.; Cleyet-Marel, J.C.; Touraine, B. Nitrate-dependent control of root architecture and N nutrition are altered by a plant growth-promoting Phyllobacterium sp. Planta 2006, 223, 591–603. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, B.J.; Boukcim, H.; Plassard, C. A comparison of ammonium, nitrate and proton net fluxes along seedling roots of Douglas-fir and lodgepole pine grown and measured with different inorganic nitrogen sources. Plant Cell Environ. 2008, 31, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, A.M.; Torres, M.P.N.; Jimenez, G.S.; Sosa, F.C. Changes in biomass allocation and phenolic compounds accumulation due to the effect of light and nitrate supply in Cecropia peltata plants. Acta Physiol. Plant 2011, 33, 2135–2147. [Google Scholar] [CrossRef]
- Kovacik, J.; Klejdus, B.; Stork, F.; Hedbavny, J. Nitrate Deficiency Reduces Cadmium and Nickel Accumulation in Chamomile Plants. J. Agric. Food Chem. 2011, 59, 5139–5149. [Google Scholar] [CrossRef]
- Tsay, Y.F.; Ho, C.H.; Chen, H.Y.; Lin, S.H. Integration of Nitrogen and Potassium Signaling. Annu. Rev. Plant Biol. 2011, 62, 207–226. [Google Scholar] [CrossRef]
- Ishikawa, K.; Ohmori, T.; Miyamoto, H.; Ito, T.; Kumagai, Y.; Sonoda, M.; Matsumoto, J.; Miyamoto, H.; Kodama, H. Denitrification in soil amended with thermophile-fermented compost suppresses nitrate accumulation in plants. Appl. Microbiol. Biot. 2013, 97, 1349–1359. [Google Scholar] [CrossRef]
- Crawford, N.M.; Glass, A.D.M. Molecular and physiological aspects of nitrate uptake in plants. Trends Plant Sci. 1998, 3, 389–395. [Google Scholar] [CrossRef]
- Krouk, G.; Tillard, P.; Gojon, A. Regulation of the high-affinity NO3− uptake system by NRT1.1-mediated NO3− demand signaling in Arabidopsis. Plant Physiol. 2006, 142, 1075–1086. [Google Scholar] [CrossRef]
- Leran, S.; Varala, K.; Boyer, J.C.; Chiurazzi, M.; Crawford, N.; Daniel-Vedele, F.; David, L.; Dickstein, R.; Fernandez, E.; Forde, B.; et al. A unified nomenclature of NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER family members in plants. Trends Plant Sci. 2014, 19, 5–9. [Google Scholar] [CrossRef]
- Chiba, Y.; Shimizu, T.; Miyakawa, S.; Kanno, Y.; Koshiba, T.; Kamiya, Y.; Seo, M. Identification of Arabidopsis thaliana NRT1/PTR FAMILY (NPF) proteins capable of transporting plant hormones. J. Plant Res. 2015, 128, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.Z.; Fang, S.Q.; Ye, Z.Q.; Liu, D.; Zhao, K.L.; Jin, C.W. NRT1.1 Dual-Affinity Nitrate Transport/Signalling and its Roles in Plant Abiotic Stress Resistance. Front. Plant Sci. 2021, 12, 715694. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Pan, L.J.; Yang, Z.; Su, M.W.; Xu, J.; Jiang, X.; Yin, X.Z.; Wang, T.; Wan, F.F.; Chi, X.Y. A MYB-related transcription factor from peanut, AhMYB30, improves freezing and salt stress tolerance in transgenic Arabidopsis through both DREB/CBF and ABA-signaling pathways. Front. Plant Sci. 2023, 14, 1136626. [Google Scholar] [CrossRef] [PubMed]
- Mozzanini, E.; Grella, M.; Marucco, P.; Balsari, P.; Gioelli, F. Characterization of irrigator emitter to be used as Solid Set Canopy Delivery System: Which is best for which role in the vineyard? Pest. Manag. Sci. 2023, 79, 584–597. [Google Scholar] [CrossRef]
- Guo, F.Q.; Young, J.; Crawford, N.M. The nitrate transporter AtNRT1.1 (CHL1) functions in stomatal opening and contributes to drought susceptibility in Arabidopsis. Plant Cell 2003, 15, 107–117. [Google Scholar] [CrossRef]
- Guo, F.Q.; Wang, R.; Chen, M.; Crawford, N.M. The Arabidopsis dual-affinity nitrate transporter gene AtNRT1.1 (CHL1) is activated and functions in nascent organ development during vegetative and reproductive growth. Plant Cell 2001, 13, 1761–1777. [Google Scholar] [CrossRef]
- Alvarez-Aragon, R.; Haro, R.; Benito, B.; Rodriguez-Navarro, A. Salt intolerance in Arabidopsis: Shoot and root sodium toxicity, and inhibition by sodium-plus-potassium overaccumulation. Planta 2016, 243, 97–114. [Google Scholar] [CrossRef]
- Alvarez-Aragon, R.; Rodriguez-Navarro, A. Nitrate-dependent shoot sodium accumulation and osmotic functions of sodium in Arabidopsis under saline conditions. Plant J. 2017, 91, 208–219. [Google Scholar] [CrossRef]
- Liu, Y.J.; Gao, N.; Ma, Q.J.; Zhang, J.C.; Wang, X.; Lu, J.; Hao, Y.J.; Wang, X.F.; You, C.X. The MdABI5 transcription factor interacts with the MdNRT1.5/MdNPF7.3 promoter to fine-tune nitrate transport from roots to shoots in apple. Hortic. Res. 2021, 8, 236. [Google Scholar] [CrossRef]
- Gong, X.Y.; Deng, W.P.; Wen, B.B.; Xiao, W.; Li, L. Genome-wide analysis of cysteine proteases (CP) in apple (Malus domestica) and functional characterization of MdCP37 in response to drought stress. Plant Cell Tissue Organ 2022, 151, 651–661. [Google Scholar] [CrossRef]
- Qiao, Z.W.; Wang, D.R.; Wang, X.; You, C.X.; Wang, X.F. Genome-wide identification and stress response analysis of cyclophilin gene family in apple (Malus × domestica). BMC Genom. 2022, 23, 806. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.L.; Zhang, Z.X.; Cheng, J.; Xian, X.L.; Li, C.L.; Wang, Y.X. Genome-wide identification of the CER1 gene family in apple and response of MdCER1-1 to drought stress. Funct. Integr. Genom. 2023, 23, 17. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.X.; Li, X.H.; Yin, B.Y.; Sun, Y.; Liang, B.W.; Li, Z.Y.; Zhang, X.Y.; Xu, J.Z.; Zhou, S.S. Tolerance of transgenic Arabidopsis thaliana overexpressing apple MdAGO4.1 gene to drought and salt stress. J. Appl. Bot. Food Qual. 2023, 96, 11–19. [Google Scholar]
- Liu, R.X.; Li, H.L.; Rui, L.; Liu, G.D.; Wang, T.; Wang, X.F.; Li, L.G.; Zhang, Z.L.; You, C.X. An apple NITRATE REDUCTASE 2 gene positively regulates nitrogen utilization and abiotic stress tolerance in Arabidopsis and apple callus. Plant Physiol. Biochem. 2023, 196, 23–32. [Google Scholar] [CrossRef] [PubMed]
- David, L.C.; Berquin, P.; Kanno, Y.; Seo, M.; Daniel-Vedele, F.; Ferrario-Mery, S. N availability modulates the role of NPF3.1, a gibberellin transporter, in GA-mediated phenotypes in Arabidopsis. Planta 2016, 244, 1315–1328. [Google Scholar] [CrossRef]
- Bakshi, A.; Shemansky, J.M.; Chang, C.R.; Binder, B.M. History of Research on the Plant Hormone Ethylene. J. Plant Growth Regul. 2015, 34, 809–827. [Google Scholar] [CrossRef]
- Ruiz Herrera, L.F.; Shane, M.W.; Lopez-Bucio, J. Nutritional regulation of root development. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 431–443. [Google Scholar] [CrossRef]
- Gaber, R.F.; Styles, C.A.; Fink, G.R. TRK1 encodes a plasma membrane protein required for high-affinity potassium transport in Saccharomyces cerevisiae. Mol. Cell Biol. 1988, 8, 2848–2859. [Google Scholar]
- Lagarde, D.; Basset, M.; Lepetit, M.; Conejero, G.; Gaymard, F.; Astruc, S.; Grignon, C. Tissue-specific expression of Arabidopsis AKT1 gene is consistent with a role in K+ nutrition. Plant J. 1996, 9, 195–203. [Google Scholar] [CrossRef]
- Ragel, P.; Rodenas, R.; Garcia-Martin, E.; Andres, Z.; Villalta, I.; Nieves-Cordones, M.; Rivero, R.M.; Martinez, V.; Pardo, J.M.; Quintero, F.J.; et al. The CBL-Interacting Protein Kinase CIPK23 Regulates HAK5-Mediated High-Affinity K+ Uptake in Arabidopsis Roots. Plant Physiol. 2015, 169, 2863–2873. [Google Scholar]
- Truong, H.A.; Lee, W.J.; Jeong, C.Y.; Trinh, C.S.; Lee, S.; Kang, C.S.; Cheong, Y.K.; Hong, S.W.; Lee, H. Enhanced anthocyanin accumulation confers increased growth performance in plants under low nitrate and high salt stress conditions owing to active modulation of nitrate metabolism. J. Plant Physiol. 2018, 231, 41–48. [Google Scholar] [CrossRef]
- Tada, Y.; Endo, C.; Katsuhara, M.; Horie, T.; Shibasaka, M.; Nakahara, Y.; Kurusu, T. High-Affinity K+ Transporters from a Halophyte, Sporobolus virginicus, Mediate Both K+ and Na+ Transport in Transgenic Arabidopsis, X. laevis Oocytes and Yeast. Plant Cell Physiol. 2019, 60, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Hrmova, M.; Gilliham, M. High affinity Na+ transport by wheat HKT1;5 is blocked by K+. Plant Direct 2020, 4, e00275. [Google Scholar] [CrossRef] [PubMed]
- Rodenas, R.; Ragel, P.; Nieves-Cordones, M.; Martinez-Martinez, A.; Amo, J.; Lara, A.; Martinez, V.; Quintero, F.J.; Pardo, J.M.; Rubio, F. Insights into the mechanisms of transport and regulation of the arabidopsis high-affinity K+ transporter HAK5. Plant Physiol. 2021, 185, 1860–1874. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.H.; Huang, C.Y.; Tsay, Y.F. CHL1 is a dual-affinity nitrate transporter of Arabidopsis involved in multiple phases of nitrate uptake. Plant Cell 1999, 11, 865–874. [Google Scholar] [CrossRef] [PubMed]
- Valliyodan, B.; Nguyen, H.T. Understanding regulatory networks and engineering for enhanced drought tolerance in plants. Curr. Opin. Plant Biol. 2006, 9, 189–195. [Google Scholar] [CrossRef]
- Qian, Y.; Zhang, T.; Yu, Y.; Gou, L.; Yang, J.; Xu, J.; Pi, E. Regulatory Mechanisms of bHLH Transcription Factors in Plant Adaptive Responses to Various Abiotic Stresses. Front. Plant Sci. 2021, 12, 677611. [Google Scholar] [CrossRef]
- Wang, D.R.; Yang, K.; Wang, X.; Lin, X.L.; Rui, L.; Liu, H.F.; Liu, D.D.; You, C.X. Overexpression of MdZAT5, an C2H2-Type Zinc Finger Protein, Regulates Anthocyanin Accumulation and Salt Stress Response in Apple Calli and Arabidopsis. Int. J. Mol. Sci. 2022, 23, 1897. [Google Scholar] [CrossRef]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.H.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef]
- Park, H.J.; Kim, W.Y.; Yun, D.J. A New Insight of Salt Stress Signaling in Plant. Mol. Cells 2016, 39, 447–459. [Google Scholar] [CrossRef]
- Mittler, R.; Blumwald, E. The roles of ROS and ABA in systemic acquired acclimation. Plant Cell 2015, 27, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.Y.; Zheng, P.F.; Ren, Y.R.; Yao, Y.X.; You, C.X.; Wang, X.F.; Hao, Y.J. Apple MdSAT1 encodes a bHLHm1 transcription factor involved in salinity and drought responses. Planta 2021, 253, 46. [Google Scholar] [CrossRef] [PubMed]
- An, J.P.; Zhang, X.W.; Liu, Y.J.; Wang, X.F.; You, C.X.; Hao, Y.J. ABI5 regulates ABA-induced anthocyanin biosynthesis by modulating the MYB1-bHLH3 complex in apple. J. Exp. Bot. 2021, 72, 1460–1472. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cis-Element Name | Cis-Element Sequence (5′-3′) | Function | Location |
|---|---|---|---|
| ARE | AAACCA | A cis-acting regulatory element essential for anaerobic induction | −15 |
| MBS | CAACTG | MYB binding site involved in drought-inducibility | −1683 |
| GT1-motif | GGTTAA | Light-responsive element | −591 |
| WUN-motif | AAATTACT | Wound-responsive element | +1196 |
| TCA-element | TCAGAAGAGG | Cis-acting element involved in salicylic acid responsiveness | +557 |
| LTR | CCGAAA | Cis-acting element involved in low-temperature responsiveness | +1265 |
| GARE-motif | TCTGTTG | Gibberellin-responsive element | −1905 |
| CAT-box | GCCACT | Cis-acting regulatory element related to meristem expression | −791 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, G.; Rui, L.; Yang, Y.; Liu, R.; Li, H.; Ye, F.; You, C.; Zhang, S. Identification and Functional Characterization of MdNRT1.1 in Nitrogen Utilization and Abiotic Stress Tolerance in Malus domestica. Int. J. Mol. Sci. 2023, 24, 9291. https://doi.org/10.3390/ijms24119291
Liu G, Rui L, Yang Y, Liu R, Li H, Ye F, You C, Zhang S. Identification and Functional Characterization of MdNRT1.1 in Nitrogen Utilization and Abiotic Stress Tolerance in Malus domestica. International Journal of Molecular Sciences. 2023; 24(11):9291. https://doi.org/10.3390/ijms24119291
Chicago/Turabian StyleLiu, Guodong, Lin Rui, Yuying Yang, Ranxin Liu, Hongliang Li, Fan Ye, Chunxiang You, and Shuai Zhang. 2023. "Identification and Functional Characterization of MdNRT1.1 in Nitrogen Utilization and Abiotic Stress Tolerance in Malus domestica" International Journal of Molecular Sciences 24, no. 11: 9291. https://doi.org/10.3390/ijms24119291
APA StyleLiu, G., Rui, L., Yang, Y., Liu, R., Li, H., Ye, F., You, C., & Zhang, S. (2023). Identification and Functional Characterization of MdNRT1.1 in Nitrogen Utilization and Abiotic Stress Tolerance in Malus domestica. International Journal of Molecular Sciences, 24(11), 9291. https://doi.org/10.3390/ijms24119291