
A Targeted Mass Spectrometry Approach to Identify Peripheral Changes in Metabolic Pathways of Patients with Alzheimer’s Disease

 , ,
, ,  , , , , and
, , , , and
Abstract
1. Introduction
2. Results
2.1. Demographics of Participants
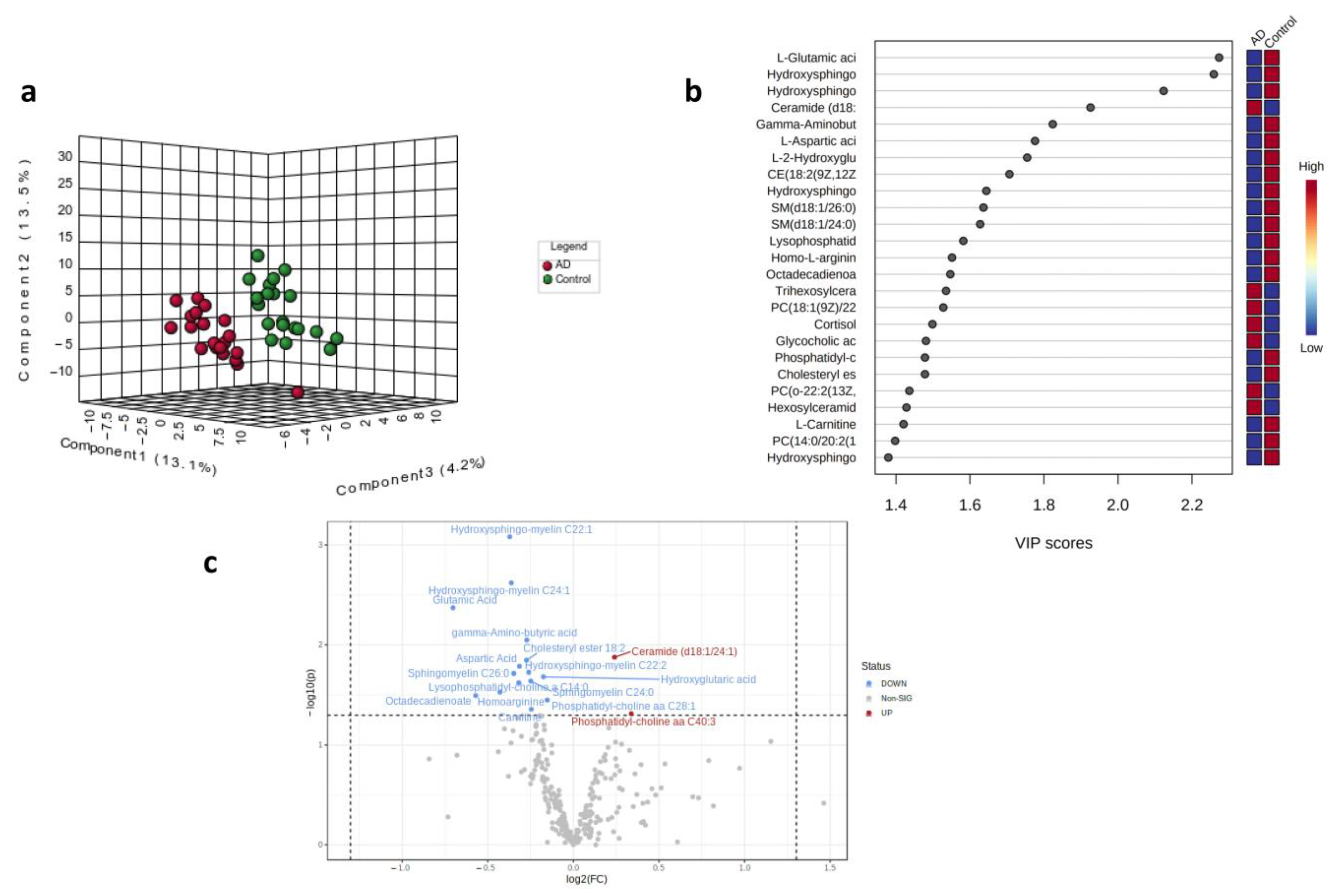
2.2. Partial Least Squares Discriminant Analysis (PLS-DA) and Volcano Plot for Control and AD Subjects
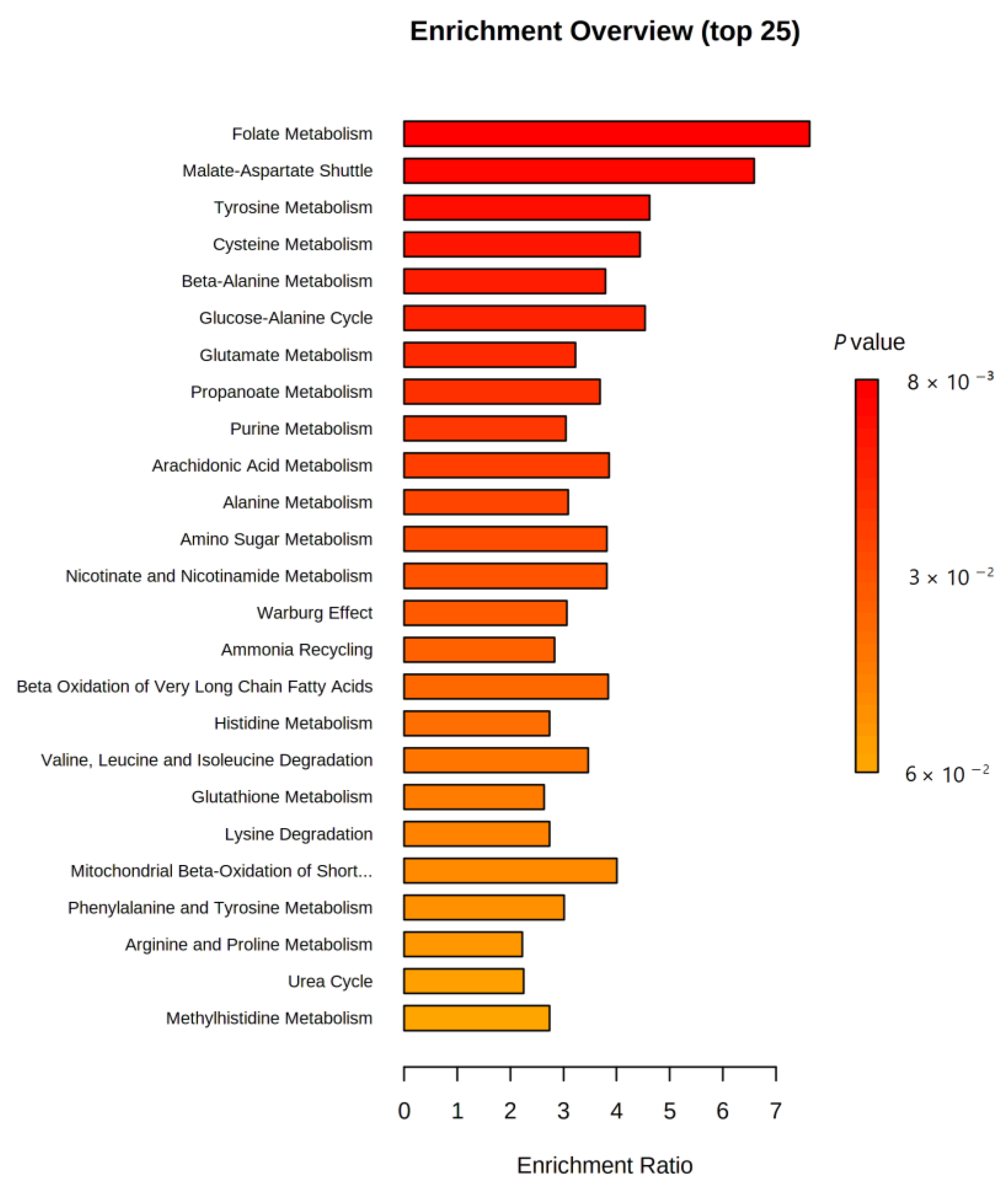
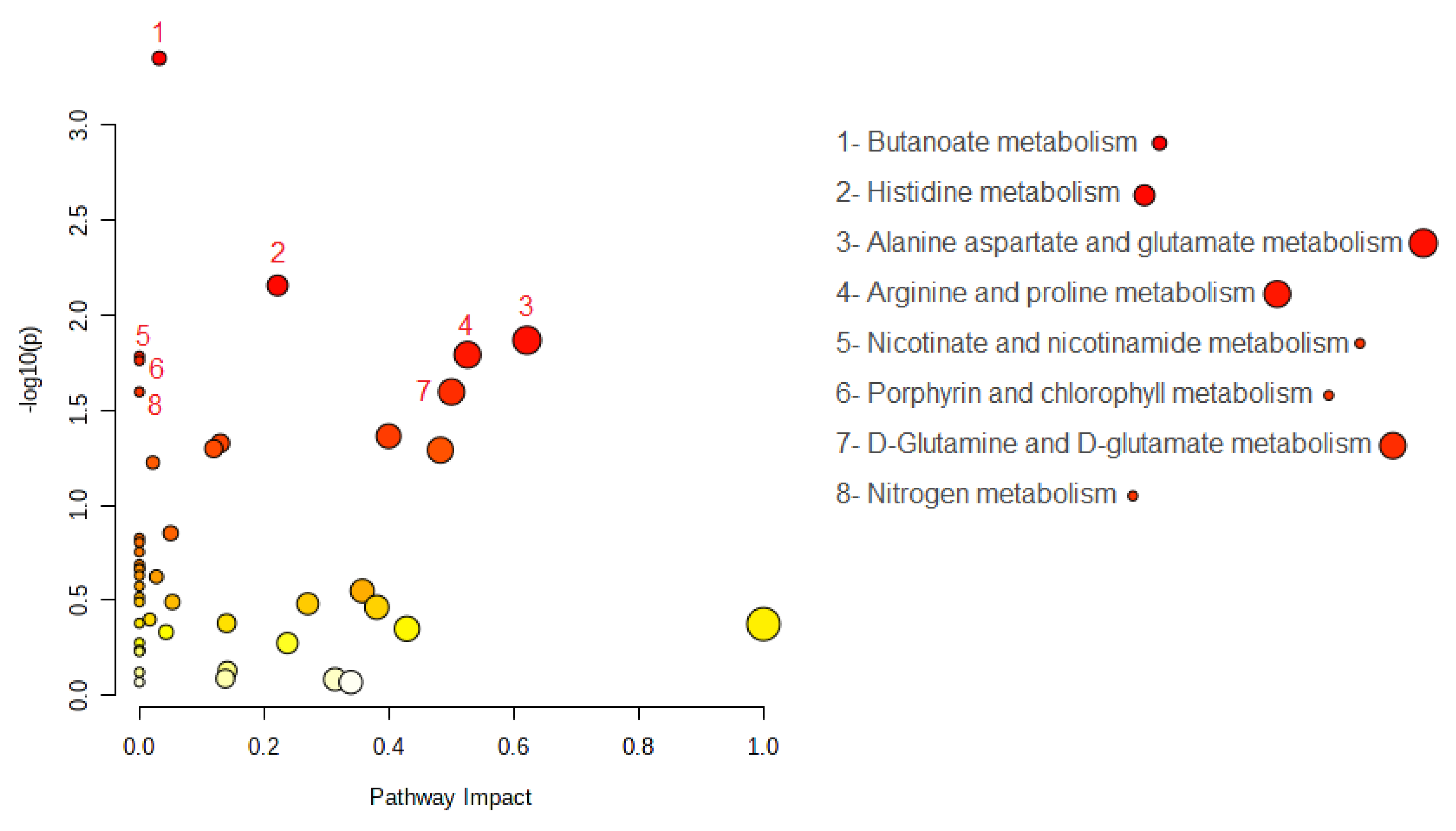
2.3. MSEA and Pathway Analysis
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Blood Sample Collection
4.3. Sample Analysis by MxP Quant 500 Assay by Biocrates Life Sciences
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alzheimer’s Association. 2022 Alzheimer’s disease facts and figures. Alzheimer’s Dement 2022, 18, 700–789. [Google Scholar] [CrossRef]
- Razay, G.; Vreugdenhil, A.; Wilcock, G. The metabolic syndrome and Alzheimer disease. Arch. Neurol. 2007, 64, 93–96. [Google Scholar] [CrossRef]
- Litke, R.; Garcharna, L.C.; Jiwani, S.; Neugroschl, J. Modifiable Risk Factors in Alzheimer Disease and Related Dementias: A Review. Clin. Ther. 2021, 43, 953–965. [Google Scholar] [CrossRef]
- Butler, L.M.; Houghton, R.; Abraham, A.; Vassilaki, M.; Durán-Pacheco, G. Comorbidity Trajectories Associated with Alzheimer’s Disease: A Matched Case-Control Study in a United States Claims Database. Front. Neurosci 2021, 15, 749305. [Google Scholar] [CrossRef]
- Galvin, J.E.; Chrisphonte, S.; Chang, L.-C. Medical and Social Determinants of Brain Health and Dementia in a Multicultural Community Cohort of Older Adults. J. Alzheimers Dis. 2021, 84, 1563–1576. [Google Scholar] [CrossRef]
- Rujeedawa, T.; Carrillo Félez, E.; Clare, I.C.H.; Fortea, J.; Strydom, A.; Rebillat, A.-S.; Coppus, A.; Levin, J.; Zaman, S.H. The Clinical and Neuropathological Features of Sporadic (Late-Onset) and Genetic Forms of Alzheimer’s Disease. J. Clin. Med. 2021, 10, 4582. [Google Scholar] [CrossRef]
- Frisardi, V.; Matrone, C.; Street, M.E. Metabolic Syndrome and Autophagy: Focus on HMGB1 Protein. Front. Cell Dev. Biol. 2021, 9, 654913. [Google Scholar] [CrossRef]
- Matrone, C.; Iannuzzi, F.; Annunziato, L. The Y682ENPTY687 motif of APP: Progress and insights toward a targeted therapy for Alzheimer’s disease patients. Ageing Res. Rev. 2019, 52, 120–128. [Google Scholar] [CrossRef]
- Matrone, C. A new molecular explanation for age-related neurodegeneration: The Tyr682 residue of amyloid precursor protein. Bioessays 2013, 35, 847–852. [Google Scholar] [CrossRef]
- Reveglia, P.; Paolillo, C.; Ferretti, G.; De Carlo, A.; Angiolillo, A.; Nasso, R.; Caputo, M.; Matrone, C.; di Costanzo, A.; Corso, G. Challenges in LC-MS-based metabolomics for Alzheimer’s disease early detection: Targeted approaches versus untargeted approaches. Metabolomics 2021, 17, 78. [Google Scholar] [CrossRef]
- Fabelo, N.; Martín, V.; Marín, R.; Moreno, D.; Ferrer, I.; Díaz, M. Altered lipid composition in cortical lipid rafts occurs at early stages of sporadic Alzheimer’s disease and facilitates APP/BACE1 interactions. Neurobiol. Aging 2014, 35, 1801–1812. [Google Scholar] [CrossRef] [PubMed]
- Frisardi, V.; Panza, F.; Seripa, D.; Farooqui, T.; Farooqui, A.A. Glycerophospholipids and glycerophospholipid-derived lipid mediators: A complex meshwork in Alzheimer’s disease pathology. Prog. Lipid. Res. 2011, 50, 313–330. [Google Scholar] [CrossRef] [PubMed]
- Ball, M.J. Frequency of stages of Alzheimer-related lesions in different age categories: Concurrences and cautions. Neurobiol. Aging 1997, 18, 375–376; discussion 389–392. [Google Scholar] [CrossRef]
- Braak, H.; Braak, E. Frequency of stages of Alzheimer-related lesions in different age categories. Neurobiol. Aging 1997, 18, 351–357. [Google Scholar] [CrossRef] [PubMed]
- El Gaamouch, F.; Jing, P.; Xia, J.; Cai, D. Alzheimer’s Disease Risk Genes and Lipid Regulators. J. Alzheimer’s Dis. 2016, 53, 15–29. [Google Scholar] [CrossRef]
- Cristofano, A.; Sapere, N.; La Marca, G.; Angiolillo, A.; Vitale, M.; Corbi, G.; Scapagnini, G.; Intrieri, M.; Russo, C.; Corso, G.; et al. Serum Levels of Acyl-Carnitines along the Continuum from Normal to Alzheimer’s Dementia. PLoS ONE 2016, 11, e0155694. [Google Scholar] [CrossRef]
- di Costanzo, A.; Paris, D.; Melck, D.; Angiolillo, A.; Corso, G.; Maniscalco, M.; Motta, A. Blood biomarkers indicate that the preclinical stages of Alzheimer’s disease present overlapping molecular features. Sci. Rep. 2020, 10, 15612. [Google Scholar] [CrossRef]
- Corso, G.; Cristofano, A.; Sapere, N.; la Marca, G.; Angiolillo, A.; Vitale, M.; Fratangelo, R.; Lombardi, T.; Porcile, C.; Intrieri, M.; et al. Serum Amino Acid Profiles in Normal Subjects and in Patients with or at Risk of Alzheimer Dementia. Dement. Geriatr. Cogn. Dis. Extra 2017, 7, 143–159. [Google Scholar] [CrossRef]
- Brereton, R.G.; Lloyd, G.R. Partial least squares discriminant analysis: Taking the magic away. J. Chemom. 2014, 28, 213–225. [Google Scholar] [CrossRef]
- Le Stunff, H.; Véret, J.; Kassis, N.; Denom, J.; Meneyrol, K.; Paul, J.L.; Cruciani-Guglielmacci, C.; Magnan, C.; Janel, N. Deciphering the Link Between Hyperhomocysteinemia and Ceramide Metabolism in Alzheimer-Type Neurodegeneration. Front. Neurol. 2019, 10, 807. [Google Scholar] [CrossRef]
- Filippov, V.; Song, M.A.; Zhang, K.; Vinters, H.V.; Tung, S.; Kirsch, W.M.; Yang, J.; Duerksen-Hughes, P.J. Increased ceramide in brains with Alzheimer’s and other neurodegenerative diseases. J. Alzheimer’s Dis. 2012, 29, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Nevado-Holgado, A.; Whiley, L.; Snowden, S.G.; Soininen, H.; Kloszewska, I.; Mecocci, P.; Tsolaki, M.; Vellas, B.; Thambisetty, M.; et al. Association between Plasma Ceramides and Phosphatidylcholines and Hippocampal Brain Volume in Late Onset Alzheimer’s Disease. J. Alzheimer’s Dis. 2017, 60, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Pujol-Lereis, L.M. Alteration of Sphingolipids in Biofluids: Implications for Neurodegenerative Diseases. Int. J. Mol. Sci. 2019, 20, 3564. [Google Scholar] [CrossRef] [PubMed]
- Casañas-Sánchez, V.; Pérez, J.A.; Fabelo, N.; Herrera-Herrera, A.V.; Fernández, C.; Marín, R.; Gonzalez-Montelongo, M.C.; Diaz, M. Addition of docosahexaenoic acid, but not arachidonic acid, activates glutathione and thioredoxin antioxidant systems in murine hippocampal HT22 cells: Potential implications in neuroprotection. J. Neurochem. 2014, 131, 470–483. [Google Scholar] [CrossRef] [PubMed]
- Grimm, M.O.; Grimm, H.S.; Hartmann, T. Amyloid beta as a regulator of lipid homeostasis. Trends Mol. Med. 2007, 13, 337–344. [Google Scholar] [CrossRef]
- Grimm, M.O.; Grösgen, S.; Riemenschneider, M.; Tanila, H.; Grimm, H.S.; Hartmann, T. From brain to food: Analysis of phosphatidylcholins, lyso-phosphatidylcholins and phosphatidylcholin-plasmalogens derivates in Alzheimer’s disease human post mortem brains and mice model via mass spectrometry. J. Chromatogr. A 2011, 1218, 7713–7722. [Google Scholar] [CrossRef] [PubMed]
- Jazvinšćak Jembrek, M.; Hof, P.R.; Šimić, G. Ceramides in Alzheimer’s Disease: Key Mediators of Neuronal Apoptosis Induced by Oxidative Stress and Aβ Accumulation. Oxid. Med. Cell. Longev. 2015, 2015, 346783. [Google Scholar] [CrossRef]
- Law, B.A.; Liao, X.; Moore, K.S.; Southard, A.; Roddy, P.; Ji, R.; Szulc, Z.; Ala Bielawska, A.; Schulze, P.C.; Cowart, L.A. Lipotoxic very-long-chain ceramides cause mitochondrial dysfunction, oxidative stress, and cell death in cardiomyocytes. FASEB J. 2018, 32, 1403–1416. [Google Scholar] [CrossRef]
- Hicks, D.A.; Nalivaeva, N.N.; Turner, A.J. Lipid rafts and Alzheimer’s disease: Protein-lipid interactions and perturbation of signaling. Front. Physiol. 2012, 3, 189. [Google Scholar] [CrossRef]
- Cheng, H.; Vetrivel, K.S.; Gong, P.; Meckler, X.; Parent, A.; Thinakaran, G. Mechanisms of disease: New therapeutic strategies for Alzheimer’s disease—Targeting APP processing in lipid rafts. Nat. Clin. Pract. Neurol. 2007, 3, 374–382. [Google Scholar] [CrossRef]
- Cordy, J.M.; Hooper, N.M.; Turner, A.J. The involvement of lipid rafts in Alzheimer’s disease. Mol. Membr. Biol. 2006, 23, 111–122. [Google Scholar] [CrossRef]
- Allinson, T.M.; Parkin, E.T.; Condon, T.P.; Schwager, S.L.; Sturrock, E.D.; Turner, A.J.; Hooper, N.M. The role of ADAM10 and ADAM17 in the ectodomain shedding of angiotensin converting enzyme and the amyloid precursor protein. Eur. J. Biochem. 2004, 271, 2539–2547. [Google Scholar] [CrossRef] [PubMed]
- Ehehalt, R.; Keller, P.; Haass, C.; Thiele, C.; Simons, K. Amyloidogenic processing of the Alzheimer beta-amyloid precursor protein depends on lipid rafts. J. Cell Biol. 2003, 160, 113–123. [Google Scholar] [CrossRef]
- Grimm, M.O.; Haupenthal, V.J.; Rothhaar, T.L.; Zimmer, V.C.; Grösgen, S.; Hundsdörfer, B.; Lehmann, J.; Grimm, H.S.; Hartmann, T. Effect of Different Phospholipids on α-Secretase Activity in the Non-Amyloidogenic Pathway of Alzheimer’s Disease. Int. J. Mol. Sci. 2013, 14, 5879–5898. [Google Scholar] [CrossRef] [PubMed]
- Grimm, M.O.; Mett, J.; Grimm, H.S.; Hartmann, T. APP Function and Lipids: A Bidirectional Link. Front. Mol. Neurosci. 2017, 10, 63. [Google Scholar] [CrossRef] [PubMed]
- Rimm, M.O.; Rothhaar, T.L.; Hartmann, T. The role of APP proteolytic processing in lipid metabolism. Exp. Brain Res. 2012, 217, 365–375. [Google Scholar]
- Shimohama, S.; Tanino, H.; Sumida, Y.; Tsuda, J.; Fujimoto, S. Alteration of myo-inositol monophosphatase in Alzheimer’s disease brains. Neurosci. Lett 1998, 245, 159–162. [Google Scholar] [CrossRef]
- Lee, H.N.; Sim, K.M.; Kim, H.; Ju, J.; Pae, A.N.; Park, J.B.; Ryu, H.; Seong, J. Aβ modulates actin cytoskeleton via SHIP2-mediated phosphoinositide metabolism. Sci. Rep. 2019, 9, 15557. [Google Scholar] [CrossRef]
- Matrone, C.; Petrillo, F.; Nasso, R.; Ferretti, G. Fyn Tyrosine Kinase as Harmonizing Factor in Neuronal Functions and Dysfunctions. Int. J. Mol. Sci. 2020, 21, 4444. [Google Scholar] [CrossRef]
- Arcaro, A.; Aubert, M.; del Hierro, M.E.E.; Khanzada, U.K.; Angelidou, S.; Tetley, T.D.; Bittermann, A.G.; Frame, M.C.; Seckl, M.J. Critical role for lipid raft-associated Src kinases in activation of PI3K-Akt signalling. Cell Signal 2007, 19, 1081–1092. [Google Scholar] [CrossRef]
- Iannuzzi, F.; Sirabella, R.; Canu, N.; Maier, T.J.; Annunziato, L.; Matrone, C. Fyn Tyrosine Kinase Elicits Amyloid Precursor Protein Tyr682 Phosphorylation in Neurons from Alzheimer’s Disease Patients. Cells 2020, 9, 1807. [Google Scholar] [CrossRef]
- Bhaskar, K.; Yen, S.H.; Lee, G. Disease-related modifications in tau affect the interaction between Fyn and Tau. J. Biol. Chem. 2005, 280, 35119–35125. [Google Scholar] [CrossRef]
- Bordji, K.; Becerril-Ortega, J.; Nicole, O.; Buisson, A. Activation of extrasynaptic, but not synaptic, NMDA receptors modifies amyloid precursor protein expression pattern and increases amyloid-ß production. J. Neurosci. 2010, 30, 15927–15942. [Google Scholar] [CrossRef] [PubMed]
- Bukke, V.N.; Archana, M.; Villani, R.; Romano, A.D.; Wawrzyniak, A.; Balawender, K.; Orkisz, S.; Beggiato, S.; Serviddio, G.; Cassano, T. The Dual Role of Glutamatergic Neurotransmission in Alzheimer’s Disease: From Pathophysiology to Pharmacotherapy. Int. J. Mol. Sci. 2020, 21, 7452. [Google Scholar] [CrossRef]
- Seckler, J.M.; Lewis, S.J. Advances in D-Amino Acids in Neurological Research. Int. J. Mol. Sci. 2020, 21, 7325. [Google Scholar] [CrossRef]
- Toledo, J.B.; Arnold, M.; Kastenmüller, G.; Chang, R.; Baillie, R.A.; Han, X.; Thambisetty, M.; Tenenbaum, J.D.; Suhre, K.; Thompson, J.W.; et al. Metabolic network failures in Alzheimer’s disease: A biochemical road map. Alzheimer’s Dement. 2017, 13, 965–984. [Google Scholar] [CrossRef]
- Yashiro, K.; Philpot, B.D. Regulation of NMDA receptor subunit expression and its implications for LTD, LTP, and metaplasticity. Neuropharmacology 2008, 55, 1081–1094. [Google Scholar] [CrossRef]
- Mattson, M.P. Glutamate and neurotrophic factors in neuronal plasticity and disease. Ann. N. Y. Acad. Sci. 2008, 1144, 97–112. [Google Scholar] [CrossRef]
- Zeydan, B.; Deelchand, D.K.; Tosakulwong, N.; Lesnick, T.G.; Kantarci, O.H.; Machulda, M.M.; Knopman, D.S.; Lowe, V.J.; Clifford, R.J.J.; Petersen, R.C.; et al. Decreased Glutamate Levels in Patients with Amnestic Mild Cognitive Impairment: An sLASER Proton MR Spectroscopy and PiB-PET Study. J. Neuroimaging 2017, 27, 630–636. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.H.; Yang, H.T.; Chiu, C.C.; Lane, H.Y. Blood levels of D-amino acid oxidase vs. D-amino acids in reflecting cognitive aging. Sci. Rep. 2017, 7, 14849. [Google Scholar] [CrossRef] [PubMed]
- Sang, C.; Philbert, S.A.; Hartland, D.; Unwin, R.; Dowsey, A.W.; Xu, J.; Cooper, G.J. Coenzyme A-Dependent Tricarboxylic Acid Cycle Enzymes Are Decreased in Alzheimer’s Disease Consistent with Cerebral Pantothenate Deficiency. Front. Aging Neurosci. 2022, 14, 893159. [Google Scholar] [CrossRef]
- Cleland, N.R.W.; Al-Juboori, S.I.; Dobrinskikh, E.; Bruce, K.D. Altered substrate metabolism in neurodegenerative disease: New insights from metabolic imaging. J. Neuroinflammation 2021, 18, 248. [Google Scholar] [CrossRef] [PubMed]
- van Dael, P. Role of n-3 long-chain polyunsaturated fatty acids in human nutrition and health: Review of recent studies and recommendations. Nutr. Res. Pract. 2021, 15, 137–159. [Google Scholar] [CrossRef] [PubMed]
- Gerster, H. Can adults adequately convert alpha-linolenic acid (18:3n-3) to eicosapentaenoic acid (20:5n-3) and docosahexaenoic acid (22:6n-3)? Int. J. Vitam. Nutr. Res. 1998, 68, 159–173. [Google Scholar]
- Kalecký, K.; German, D.C.; Montillo, A.A.; Bottiglieri, T. Targeted Metabolomic Analysis in Alzheimer’s Disease Plasma and Brain Tissue in Non-Hispanic Whites. J. Alzheimer’s Dis. 2022, 86, 1875–1895. [Google Scholar] [CrossRef] [PubMed]
- Casanova, R.; Varma, S.; Simpson, B.; Kim, M.; An, Y.; Saldana, S.; Riveros, C.; Moscato, P.; Griswold, M.; Sonntag, D.; et al. Blood metabolite markers of preclinical Alzheimer’s disease in two longitudinally followed cohorts of older individuals. Alzheimer’s Dement. 2016, 12, 815–822. [Google Scholar] [CrossRef]
- Varma, V.R.; Oommen, A.M.; Varma, S.; Casanova, R.; An, Y.; Andrews, R.M.; O’Brien, R.; Pletnikova, O.; Troncoso, J.C.; Toledo, J.; et al. Brain and blood metabolite signatures of pathology and progression in Alzheimer disease: A targeted metabolomics study. PLoS Med. 2018, 15, e1002482. [Google Scholar] [CrossRef]
- McKhann, G.M.; Knopman, D.S.; Chertkow, H.; Hyman, B.T.; Jack Jr, C.R.; Kawas, C.H.; Klunk, W.E.; Koroshetz, W.J.; Manly, J.J.; Mayeux, R.; et al. The diagnosis of dementia due to Alzheimer’s disease: Recommendations from the National Institute on Aging-Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimers Dement 2011, 7, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.E.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Controls (n = 20) | AD (n = 20) | p-Value | |
|---|---|---|---|
| Age (mean ± SD) | 76.85 ± 7.9 | 80.85 ± 5.3 | 0.075 |
| Gender (n %): | 1 | ||
| Male | 10 (50%) | 8 (40%) | |
| Female | 10 (50%) | 12 (60%) | |
| Education (mean of years ± SD) | 11.9 ± 3.7 | 8.35 ± 3.4 | 0.003406 * |
| MMSE score (mean ± SD) | 29.83 ± 1.2 | 20.95 ± 2.9 | 2.86 × 10−12 * |
| GDS (mean ± SD) | 3.05 ± 2.9 | 5.38 ± 3.5 | 0.060041 |
| BMI (mean ± SD) | 27.08 ± 1.6 | 25.3 ± 3.9 | 0.108194 |
| Comorbidities (n, %): | |||
| Alcohol | 12 (60%) | 11 (55%) | 0.749 |
| Smoker | 12 (60%) | 6 (30%) | 0–056 |
| Dyslipidaemia | 6 (30%) | 3 (15%) | 0.255 |
| Diabetes | 3 (15%) | 3 (15%) | 1 |
| Hypertension | 9 (45%) | 6 (30%) | 0.327 |
| TIA/ischemia | 1 (5%) | 1 (5%) | 1 |
| Cardiac ischemia | 0 (0%) | 2 (10%) | 0.167 |
| Prior tumors | 4 (20%) | 2 (10%) | 0.375 |
| Drugs: | |||
| Antihypertension | 9 (45%) | 6 (30%) | 0.327 |
| Hypoglycaemic | 3 (15%) | 3 (15%) | 1 |
| Hypolipidemic | 6 (30%) | 3 (15%) | 0.255 |
| Antiplatelet | 1 (5%) | 3 (15%) | 0.197 |
| Antiacid | 2 (10%) | 4 (20%) | 0.375 |
| Compound | Class | Type | p-Value | FC | log2(FC) | VIP |
|---|---|---|---|---|---|---|
| Hydroxysphingo-myelin C22:1 | sphingolipid | Down | 0.0008 | 0.7728 | −0.3718 | 2.775 |
| Hydroxysphingo-myelin C24:2 | sphingolipid | Down | 0.0024 | 0.7779 | −0.3624 | 2.552 |
| L-Glutamic acid | amino acid | Down | 0.0043 | 0.6142 | −0.7032 | 2.419 |
| Gamma-Aminobutyric acid | amino acid | Down | 0.0089 | 0.8282 | −0.272 | 2.231 |
| Ceramide (d18:1/24:1) | lipid | Up | 0.0133 | 1.1814 | 0.2405 | 2.123 |
| CE (18:2(9Z,12Z)) | lipid | Down | 0.0142 | 0.8273 | −0.2735 | 2.103 |
| L-Aspartic acid | amino acid | Down | 0.0164 | 0.8039 | −0.315 | 2.063 |
| Hydroxysphingo-myelin C22:2 | sphingolipid | Down | 0.0188 | 0.8346 | −0.2608 | 2.023 |
| SM(d18:1/26:0) | sphingolipid | Down | 0.0193 | 0.7856 | −0.3482 | 2.015 |
| L-2-Hydroxyglutaric acid | organic acid | Down | 0.0207 | 0.8858 | −0.175 | 1.993 |
| SM (d18:1/24:0) | sphingolipid | Down | 0.0229 | 0.8416 | −0.2488 | 1.963 |
| Lysophosphatidyl-choline C14:1 | phospholipid | Down | 0.0238 | 0.8017 | −0.319 | 1.951 |
| Homo-L-arginine | amino acid | Down | 0.0296 | 0.7429 | −0.4287 | 1.883 |
| Octadecadienoate | fatty acid derivative | Down | 0.0321 | 0.6734 | −0.5705 | 1.856 |
| Phosphatidyl-choline aa C28:1 | phospholipid | Down | 0.0355 | 0.8997 | −0.1524 | 1.823 |
| L-Carnitine | amino acid | Down | 0.0438 | 0.8435 | −0.2456 | 1.752 |
| PC aa C40:3 | phospholipid | Up | 0.0486 | 1.2641 | 0.3381 | 1.716 |
| Pathway | Total Metabolites | Hits Metabolites | Raw p | FDR |
|---|---|---|---|---|
| Biosynthesis of unsaturated fatty acids | 36 | 4 | 0.176 | 0.482 |
| Steroid hormone biosynthesis | 85 | 2 | 0.238 | 0.482 |
| Fatty acid degradation | 39 | 1 | 0.305 | 0.482 |
| Primary bile acid biosynthesis | 46 | 4 | 0.321 | 0.482 |
| Lysine degradation | 25 | 1 | 0.684 | 0.777 |
| Arachidonic acid metabolism | 36 | 1 | 0.777 | 0.777 |
| Pathway | Raw p | FDR | Total Metabolites in the Pathway | Hits Metabolites | Altered Metabolites | Type |
|---|---|---|---|---|---|---|
| Butanoate metabolism | 0.0004 | 0.02 | 15 | 4-Aminobutanoate L-Glutamate | L-Glutamate | Down |
| Histidine metabolism | 0.0070 | 0.13 | 16 | L-Glutamate L-Histidine N(pi)-Methyl-L-histidine L-Aspartate | L-Glutamate L-Aspartate | Down Down |
| Alanine aspartate and glutamate metabolism | 0.0135 | 0.13 | 28 | L-Aspartate L-Asparagine L-Alanine L-Glutamate 4-Aminobutanoate L-Glutamine | 4-Aminobutanoate L-Glutamate | Down Down |
| Arginine and proline metabolism | 0.0161 | 0.13 | 38 | L-Arginine 4-Aminobutanoate Putrescine Hydroxyproline L-Proline L-Glutamate L-Ornithine | 4-Aminobutanoate L-Glutamate | Down Down |
| Nicotinate and nicotinamide metabolism | 0.0164 | 0.13 | 15 | L-Aspartate | L-Aspartate | Down |
| Porphyrin and chlorophyll metabolism | 0.0173 | 0.13 | 30 | Glycine L-Glutamate | L-Glutamate | Down |
| D-Glutamine and D-glutamate metabolism | 0.0253 | 0.14 | 6 | L-Glutamate L-Glutamine | L-Glutamate | Down |
| Nitrogen metabolism | 0.0253 | 0.14 | 6 | L-Glutamate L-Glutamine | L-Glutamate | Down |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reveglia, P.; Paolillo, C.; Angiolillo, A.; Ferretti, G.; Angelico, R.; Sirabella, R.; Corso, G.; Matrone, C.; Di Costanzo, A. A Targeted Mass Spectrometry Approach to Identify Peripheral Changes in Metabolic Pathways of Patients with Alzheimer’s Disease. Int. J. Mol. Sci. 2023, 24, 9736. https://doi.org/10.3390/ijms24119736
Reveglia P, Paolillo C, Angiolillo A, Ferretti G, Angelico R, Sirabella R, Corso G, Matrone C, Di Costanzo A. A Targeted Mass Spectrometry Approach to Identify Peripheral Changes in Metabolic Pathways of Patients with Alzheimer’s Disease. International Journal of Molecular Sciences. 2023; 24(11):9736. https://doi.org/10.3390/ijms24119736
Chicago/Turabian StyleReveglia, Pierluigi, Carmela Paolillo, Antonella Angiolillo, Gabriella Ferretti, Ruggero Angelico, Rossana Sirabella, Gaetano Corso, Carmela Matrone, and Alfonso Di Costanzo. 2023. "A Targeted Mass Spectrometry Approach to Identify Peripheral Changes in Metabolic Pathways of Patients with Alzheimer’s Disease" International Journal of Molecular Sciences 24, no. 11: 9736. https://doi.org/10.3390/ijms24119736
APA StyleReveglia, P., Paolillo, C., Angiolillo, A., Ferretti, G., Angelico, R., Sirabella, R., Corso, G., Matrone, C., & Di Costanzo, A. (2023). A Targeted Mass Spectrometry Approach to Identify Peripheral Changes in Metabolic Pathways of Patients with Alzheimer’s Disease. International Journal of Molecular Sciences, 24(11), 9736. https://doi.org/10.3390/ijms24119736