

Role of CIV NS1 Protein in Innate Immunity and Viral Replication

 ,
,
Abstract
:1. Introduction
2. Results
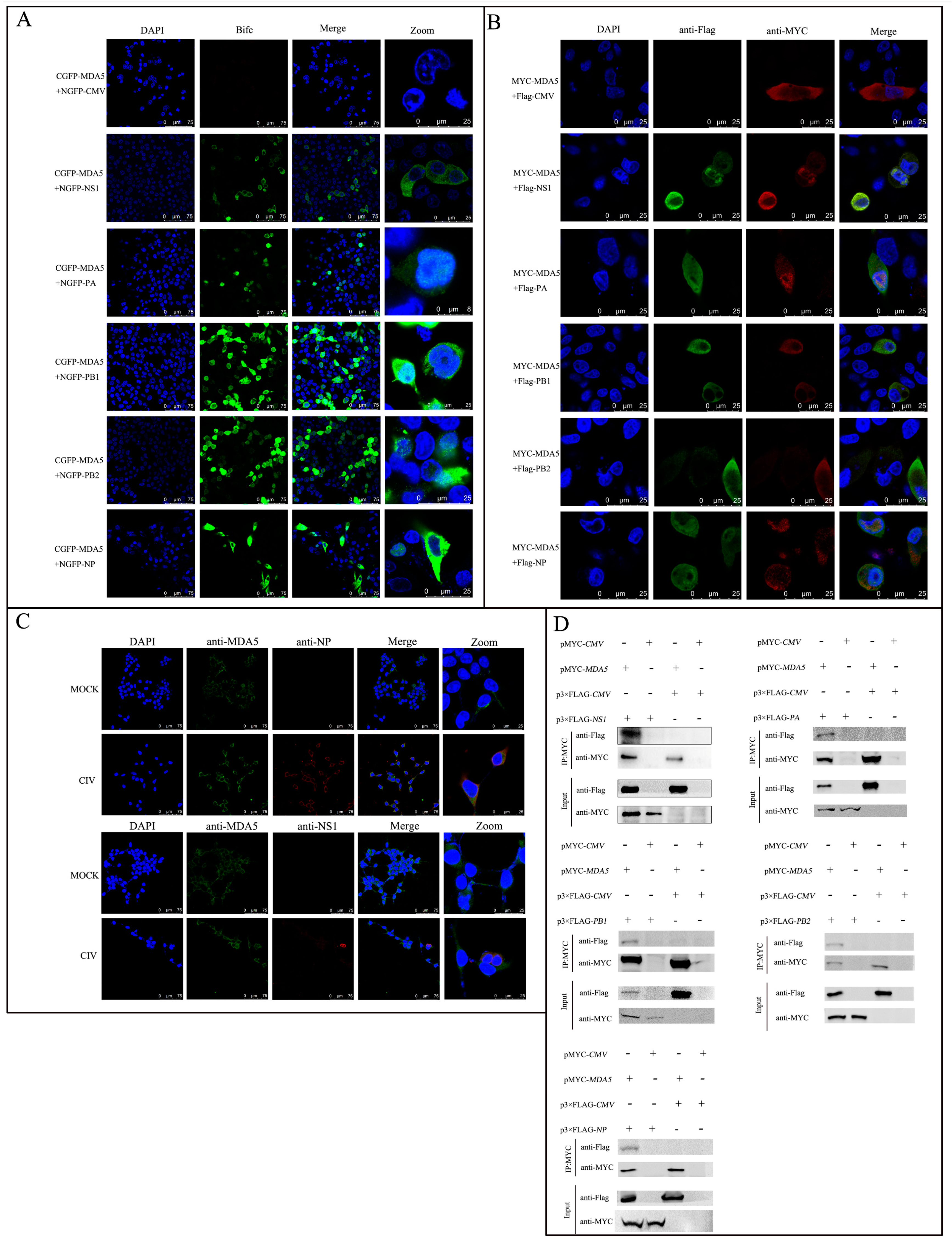
2.1. CIV Proteins Interact with MDA5
2.2. NS1 Does Not Affect the Interaction between the RNP Subunits and MDA5
2.3. Effect of CIV Protein on IFN-β Promoter Activated by MDA5
2.4. Overexpression of NS1 Protein Antagonizes the Activation of IRF3 and IFN-β Promoters by the CARD Region of MDA5
2.5. NS1 Protein Inhibited the Expression of the RIG-I Pathway and Its Downstream ISGs and Cytokines
2.6. Recombinant NS1-Null Virus Strains Promoted Activation of the IFN-β and IRF3 Promoters
2.7. Deletion of NS1 Protein Inhibited H3N2 Replication
2.8. Deletion of NS1 Protein Inhibited Innate Immunity
3. Discussion
4. Materials and Methods
4.1. Viruses and Cells
4.2. Reagents and Antibodies
4.3. Quantitative Real-Time RT-PCR (qPCR)
4.4. Construction of Plasmids
4.5. Transfection
4.6. Luciferase Assay
4.7. Rescue of rH3N2 and rH3N2ΔNS1 Viruses
4.8. Bimolecular Fluorescence Complementation (BiFC)
4.9. Coimmunoprecipitation (co-IP) and Western Blotting
4.10. Confocal Immunofluorescence Microscopy
4.11. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Shao, W.; Li, X.; Goraya, M.U.; Wang, S.; Chen, J.L. Evolution of Influenza A Virus by Mutation and Re-Assortment. Int. J. Mol. Sci. 2017, 18, 1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, X.; Ma, K.; Liu, Y. Influenza A virus infection in dogs: Epizootiology, evolution and prevention—A review. Acta Vet. Hung. 2016, 64, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Trovao, N.S.; Wang, G.; Zhao, W.; He, P.; Zhou, H.; Mo, Y.; Wei, Z.; Ouyang, K.; Huang, W.; et al. Emergence and Evolution of Novel Reassortant Influenza A Viruses in Canines in Southern China. mBio 2018, 9, e00909-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, P.C.; Dubovi, E.J.; Castleman, W.L.; Stephenson, I.; Gibbs, E.P.; Chen, L.; Smith, C.; Hill, R.C.; Ferro, P.; Pompey, J.; et al. Transmission of equine influenza virus to dogs. Science 2005, 310, 482–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrish, C.R.; Voorhees, I. H3N8 and H3N2 Canine Influenza Viruses: Understanding These New Viruses in Dogs. Vet. Clin. N. Am. Small Anim. Pract. 2019, 49, 643–649. [Google Scholar] [CrossRef]
- Song, D.; Moon, H.J.; An, D.J.; Jeoung, H.Y.; Kim, H.; Yeom, M.J.; Hong, M.; Nam, J.H.; Park, S.J.; Park, B.K.; et al. A novel reassortant canine H3N1 influenza virus between pandemic H1N1 and canine H3N2 influenza viruses in Korea. J. Gen. Virol. 2012, 93, 551–554. [Google Scholar] [CrossRef]
- Xie, X.; Na, W.; Kang, A.; Yeom, M.; Yuk, H.; Moon, H.; Kim, S.J.; Kim, H.W.; Kim, J.K.; Pang, M.; et al. Comparison of the virulence of three H3N2 canine influenza virus isolates from Korea and China in mouse and Guinea pig models. BMC Vet. Res. 2018, 14, 149. [Google Scholar] [CrossRef]
- Cooper, D.A.; Banerjee, S.; Chakrabarti, A.; Garcia-Sastre, A.; Hesselberth, J.R.; Silverman, R.H.; Barton, D.J. RNase L targets distinct sites in influenza A virus RNAs. J. Virol. 2015, 89, 2764–2776. [Google Scholar] [CrossRef] [Green Version]
- Zhirnov, O.P. Unique Bipolar Gene Architecture in the RNA Genome of Influenza A Virus. Biochemistry 2020, 85, 387–392. [Google Scholar] [CrossRef]
- Ayllon, J.; Garcia-Sastre, A. The NS1 protein: A multitasking virulence factor. Curr. Top Microbiol. Immunol. 2015, 386, 73–107. [Google Scholar] [CrossRef]
- Cheng, J.; Tao, J.; Li, B.; Shi, Y.; Liu, H. The tyrosine 73 and serine 83 dephosphorylation of H1N1 swine influenza virus NS1 protein attenuates virus replication and induces high levels of beta interferon. Virol. J. 2019, 16, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mor, A.; White, A.; Zhang, K.; Thompson, M.; Esparza, M.; Muñoz-Moreno, R.; Koide, K.; Lynch, K.W.; García-Sastre, A.; Fontoura, B.M.A. Influenza virus mRNA trafficking through host nuclear speckles. Nat. Microbiol. 2016, 1, 16069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, C.F.; Read, E.; Wise, H.M.; Amorim, M.J.; Digard, P. Influenza A Virus NS1 Protein Promotes Efficient Nuclear Export of Unspliced Viral M1 mRNA. J. Virol. 2017, 91, e00528-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arias-Mireles, B.H.; de Rozieres, C.M.; Ly, K.; Joseph, S. RNA Modulates the Interaction between Influenza A Virus NS1 and Human PABP1. Biochemistry 2018, 57, 3590–3598. [Google Scholar] [CrossRef]
- De Lucas, S.; Peredo, J.; Marion, R.M.; Sanchez, C.; Ortin, J. Human Staufen1 protein interacts with influenza virus ribonucleoproteins and is required for efficient virus multiplication. J. Virol. 2010, 84, 7603–7612. [Google Scholar] [CrossRef] [Green Version]
- Evseev, D.; Magor, K.E. Molecular Evolution of the Influenza A Virus Non-structural Protein 1 in Interspecies Transmission and Adaptation. Front. Microbiol. 2021, 12, 693204. [Google Scholar] [CrossRef]
- Gaba, A.; Xu, F.; Lu, Y.; Park, H.S.; Liu, G.; Zhou, Y. The NS1 Protein of Influenza A Virus Participates in Necroptosis by Interacting with MLKL and Increasing Its Oligomerization and Membrane Translocation. J. Virol. 2019, 93, e01835-18. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.H.; Zhao, B.; Shi, J.; Savage, N.; Shen, Q.; Byrnes, J.; Yang, L.; Hwang, W.; Li, P. Molecular recognition of a host protein by NS1 of pandemic and seasonal influenza A viruses. Proc. Natl. Acad. Sci. USA 2020, 117, 6550–6558. [Google Scholar] [CrossRef]
- Liu, Y.; Olagnier, D.; Lin, R. Host and Viral Modulation of RIG-I-Mediated Antiviral Immunity. Front. Immunol. 2016, 7, 662. [Google Scholar] [CrossRef] [Green Version]
- Hao, Q.; Jiao, S.; Shi, Z.; Li, C.; Meng, X.; Zhang, Z.; Wang, Y.; Song, X.; Wang, W.; Zhang, R.; et al. A non-canonical role of the p97 complex in RIG-I antiviral signaling. EMBO J. 2015, 34, 2903–2920. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Shao, Q.; Zang, Y.; Guo, Q.; Zhang, Y.; Li, Z. Pigeon RIG-I Function in Innate Immunity against H9N2 IAV and IBDV. Viruses 2015, 7, 4131–4151. [Google Scholar] [CrossRef] [PubMed]
- Jureka, A.S.; Kleinpeter, A.B.; Tipper, J.L.; Harrod, K.S.; Petit, C.M. The influenza NS1 protein modulates RIG-I activation via a strain-specific direct interaction with the second CARD of RIG-I. J. Biol. Chem. 2020, 295, 1153–1164. [Google Scholar] [CrossRef] [PubMed]
- Fu, C.; Ye, S.; Liu, Y.; Li, S. Role of CARD Region of MDA5 Gene in Canine Influenza Virus Infection. Viruses 2020, 12, 307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, L.; Cui, J.; Song, Y.; Zhang, S.; Han, F.; Yuan, R.; Gong, L.; Jiao, P.; Liao, M. Duck MDA5 functions in innate immunity against H5N1 highly pathogenic avian influenza virus infections. Vet. Res. 2014, 45, 66. [Google Scholar] [CrossRef] [Green Version]
- Benitez, A.A.; Panis, M.; Xue, J.; Varble, A.; Shim, J.V.; Frick, A.L.; Lopez, C.B.; Sachs, D.; TenOever, B.R. In Vivo RNAi Screening Identifies MDA5 as a Significant Contributor to the Cellular Defense against Influenza A Virus. Cell Rep. 2015, 11, 1714–1726. [Google Scholar] [CrossRef] [Green Version]
- Tangwangvivat, R.; Chaiyawong, S.; Nonthabenjawan, N.; Charoenkul, K.; Janethanakit, T.; Udom, K.; Kesdangsakonwut, S.; Tantilertcharoen, R.; Thontiravong, A.; Amonsin, A. Transmission and pathogenicity of canine H3N2 influenza virus in dog and guinea pig models. Virol. J. 2022, 19, 162. [Google Scholar] [CrossRef]
- Haye, K.; Burmakina, S.; Moran, T.; García-Sastre, A.; Fernandez-Sesma, A. The NS1 protein of a human influenza virus inhibits type I interferon production and the induction of antiviral responses in primary human dendritic and respiratory epithelial cells. J. Virol. 2009, 83, 6849–6862. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, L.; Nogales, A.; Reilly, E.C.; Topham, D.J.; Murcia, P.R.; Parrish, C.R.; Martinez, S.L. A live-attenuated influenza vaccine for H3N2 canine influenza virus. Virology 2017, 504, 96–106. [Google Scholar] [CrossRef]
- Nogales, A.; Huang, K.; Chauche, C.; DeDiego, M.L.; Murcia, P.R.; Parrish, C.R.; Martinez-Sobrido, L. Canine influenza viruses with modified NS1 proteins for the development of live-attenuated vaccines. Virology 2017, 500, 1–10. [Google Scholar] [CrossRef]
- Nogales, A.; Rodriguez, L.; Chauche, C.; Huang, K.; Reilly, E.C.; Topham, D.J.; Murcia, P.R.; Parrish, C.R.; Martinez-Sobrido, L. Temperature-Sensitive Live-Attenuated Canine Influenza Virus H3N8 Vaccine. J. Virol. 2017, 91, e02211-16. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, L.; Nogales, A.; Murcia, P.R.; Parrish, C.R.; Martinez-Sobrido, L. A bivalent live-attenuated influenza vaccine for the control and prevention of H3N8 and H3N2 canine influenza viruses. Vaccine 2017, 35, 4374–4381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diamond, M.S.; Kanneganti, T.D. Innate immunity: The first line of defense against SARS-CoV-2. Nat. Immunol. 2022, 23, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Koyama, S.; Ishii, K.J.; Coban, C.; Akira, S. Innate immune response to viral infection. Cytokine 2008, 43, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Bhowmik, D.; Du, M.; Tian, Y.; Ma, S.; Wu, J.; Chen, Z.; Yin, Q.; Zhu, F. Cooperative DNA binding mediated by KicGAS/ORF52 oligomerization allows inhibition of DNA-induced phase separation and activation of cGAS. Nucleic Acids Res. 2021, 49, 9389–9403. [Google Scholar] [CrossRef]
- Li, X.D.; Wu, J.; Gao, D.; Wang, H.; Sun, L.; Chen, Z.J. Pivotal roles of cGAS-cGAMP signaling in antiviral defense and immune adjuvant effects. Science 2013, 341, 1390–1394. [Google Scholar] [CrossRef] [Green Version]
- Ni, G.; Ma, Z.; Damania, B. cGAS and STING: At the intersection of DNA and RNA virus-sensing networks. PLoS Pathog. 2018, 14, e1007148. [Google Scholar] [CrossRef] [Green Version]
- Schoggins, J.W.; MacDuff, D.A.; Imanaka, N.; Gainey, M.D.; Shrestha, B.; Eitson, J.L.; Mar, K.B.; Richardson, R.B.; Ratushny, A.V.; Litvak, V.; et al. Pan-viral specificity of IFN-induced genes reveals new roles for cGAS in innate immunity. Nature 2014, 505, 691–695. [Google Scholar] [CrossRef] [Green Version]
- Bhowmik, D.; Zhu, F. Evasion of Intracellular DNA Sensing by Human Herpesviruses. Front. Cell. Infect. Microbiol. 2021, 11, 647992. [Google Scholar] [CrossRef]
- Maringer, K.; Fernandez-Sesma, A. Message in a bottle: Lessons learned from antagonism of STING signalling during RNA virus infection. Cytokine Growth Factor Rev. 2014, 25, 669–679. [Google Scholar] [CrossRef] [Green Version]
- Webb, L.G.; Fernandez-Sesma, A. RNA viruses and the cGAS-STING pathway: Reframing our understanding of innate immune sensing. Curr. Opin. Virol. 2022, 53, 101206. [Google Scholar] [CrossRef]
- Bhattacharya, M.; Bhowmik, D.; Tian, Y.; He, H.; Zhu, F.; Yin, Q. The Dengue virus protease NS2B3 cleaves cyclic GMP-AMP synthase (cGAS) to suppress cGAS activation. J. Biol. Chem. 2023, 299, 102986. [Google Scholar] [CrossRef] [PubMed]
- Xiaoyong, C.; Shasha, L.; Shasha, L.; Mohsan, U.G.; Mohamed, M.; Mohamed, M.; Shile, H.; Ji-Long, C.; Ji-Long, C. Host Immune Response to Influenza A Virus Infection. Front. Immunol. 2018, 9, 320. [Google Scholar]
- Bamming, D.; Horvath, C.M. Regulation of signal transduction by enzymatically inactive antiviral RNA helicase proteins MDA5, RIG-I, and LGP2. J. Biol. Chem. 2009, 284, 9700–9712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.; Wei, F.; Zhang, Y.; Wang, T.; Gao, W.; Yu, S.; Sun, H.; Pu, J.; Sun, Y.; Wang, M.; et al. IFI16 directly senses viral RNA and enhances RIG-I transcription and activation to restrict influenza virus infection. Nat. Microbiol. 2021, 6, 932–945. [Google Scholar] [CrossRef] [PubMed]
- Fang, R.; Jiang, Q.; Zhou, X.; Wang, C.; Guan, Y.; Tao, J.; Xi, J.; Feng, J.; Jiang, Z. MAVS activates TBK1 and IKKε through TRAFs in NEMO dependent and independent manner. PLoS Pathog. 2017, 13, e1006720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, B.; Chen, Y.; Liu, X.; Yuan, F.; Tan, E.; Lei, Y.; Song, N.; Han, Y.; Pascal, B.D.; Griffin, P.R.; et al. Ordered assembly of the cytosolic RNA-sensing MDA5-MAVS signaling complex via binding to unanchored K63-linked poly-ubiquitin chains. Immunity 2021, 54, 2218–2230. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, L.; Wang, G.; Shi, W.; Hu, Y.; Wang, B.; Zeng, X.; Tian, G.; Deng, G.; Shi, J.; et al. Influenza A virus use of BinCARD1 to facilitate the binding of viral NP to importin alpha7 is counteracted by TBK1-p62 axis-mediated autophagy. Cell. Mol. Immunol. 2022, 19, 1168–1184. [Google Scholar] [CrossRef]
- Wang, Z.; Ye, S.; Yao, C.; Wang, J.; Mao, J.; Xu, L.; Liu, Y.; Fu, C.; Lu, G.; Li, S. Antiviral Activity of Canine RIG-I against Canine Influenza Virus and Interactions between Canine RIG-I and CIV. Viruses 2021, 13, 2048. [Google Scholar] [CrossRef] [PubMed]
- Liniger, M.; Summerfield, A.; Zimmer, G.; McCullough, K.C.; Ruggli, N. Chicken cells sense influenza A virus infection through MDA5 and CARDIF signaling involving LGP2. J. Virol. 2012, 86, 705–717. [Google Scholar] [CrossRef] [Green Version]
- Weber, M.; Gawanbacht, A.; Habjan, M.; Rang, A.; Borner, C.; Schmidt, A.M.; Veitinger, S.; Jacob, R.; Devignot, S.; Kochs, G.; et al. Incoming RNA virus nucleocapsids containing a 5’-triphosphorylated genome activate RIG-I and antiviral signaling. Cell Host Microbe 2013, 13, 336–346. [Google Scholar] [CrossRef] [Green Version]
- Baskin, C.R.; Bielefeldt-Ohmann, H.; Garcia-Sastre, A.; Tumpey, T.M.; Van Hoeven, N.; Carter, V.S.; Thomas, M.J.; Proll, S.; Solorzano, A.; Billharz, R.; et al. Functional genomic and serological analysis of the protective immune response resulting from vaccination of macaques with an NS1-truncated influenza virus. J. Virol. 2007, 81, 11817–11827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kochs, G.; Koerner, I.; Thiel, L.; Kothlow, S.; Kaspers, B.; Ruggli, N.; Summerfield, A.; Pavlovic, J.; Stech, J.; Staeheli, P. Properties of H7N7 influenza A virus strain SC35M lacking interferon antagonist NS1 in mice and chickens. J. Gen. Virol. 2007, 88, 1403–1409. [Google Scholar] [CrossRef] [PubMed]
- Egorov, A.; Brandt, S.; Sereinig, S.; Romanova, J.; Ferko, B.; Katinger, D.; Grassauer, A.; Alexandrova, G.; Katinger, H.; Muster, T. Transfectant influenza A viruses with long deletions in the NS1 protein grow efficiently in Vero cells. J. Virol. 1998, 72, 6437–6441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, E.; Neumann, G.; Kawaoka, Y.; Hobom, G.; Webster, R.G. A DNA transfection system for generation of influenza A virus from eight plasmids. Proc. Natl. Acad. Sci. USA 2000, 97, 6108–6113. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′~3′) |
|---|---|
| IL-1β | F: TCAAGAACACAGTGGAATTTGAGTCTT R: TCAGTTATATCCTGGCCACCTCTG |
| IL-6 | F: TTCATTCCTTAGGATAGTGCTGAG R: TCCTGAGGAGTGAAGATAACAATTT |
| IL-8 | F: AAACACACTCCACACCTTTCCAT R: GGCACACCTCATTTCCATTGAA |
| IL-2 | F: AGTAACCTCAACTCCTGCCACAAT R: TTGCTCCATCTGTTGCTCTGTTTC |
| RIG-I | F: CTCCAAGAAGAAGGCTGGTTC R: AAGCAATCTATACTCCTCTAGACTTTC |
| LGP2 | F: TCACTCCCTCCTACTCTGGCTC R: TTTCGGATCACTTCTTGCTGGTCT |
| IRF3 | F: GGACCTGCACATTTCCAACAGC R: CAGTGACCCAGAAATCCATGTCCT |
| OAS | F: CCAGGGTAACTCAGGAAGGAAAGT R: CATCTCCATCAAACACGGGCTG |
| STA1 | F: TTGACAGCAAAGTGAGAAACGTGA R: ATTGGCTTCATGTTCTCGGTTCTG |
| IFN-β | F: GAAATCACGCCAGTTCCAGAAG R: TCTCATTCCATCCTGTTCTAGAGATATT |
| IPS1 | F: GACCACAAGATGTCCGCAAGC R: GGCAAGCTGTCTCTGGTGGA |
| TRIM25 | F: TGAAACACTATATCAGGCAGTCCC R: AAATGTATGGGTTTGTGCGTGGAT |
| MX1 | F: ATCACTGACTCGAATCCTGTACCC R: GCCTACCTTCTCCTCATATTGGCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, C.; Zhu, W.; Cao, N.; Liu, W.; Lu, Z.; Wong, Z.; Guan, K.; Hu, C.; Han, B.; Zeng, S.; et al. Role of CIV NS1 Protein in Innate Immunity and Viral Replication. Int. J. Mol. Sci. 2023, 24, 10056. https://doi.org/10.3390/ijms241210056
Fu C, Zhu W, Cao N, Liu W, Lu Z, Wong Z, Guan K, Hu C, Han B, Zeng S, et al. Role of CIV NS1 Protein in Innate Immunity and Viral Replication. International Journal of Molecular Sciences. 2023; 24(12):10056. https://doi.org/10.3390/ijms241210056
Chicago/Turabian StyleFu, Cheng, Wenhui Zhu, Nan Cao, Wenjun Liu, Zhier Lu, Ziyuan Wong, Kaiting Guan, Chunyan Hu, Baoting Han, Sen Zeng, and et al. 2023. "Role of CIV NS1 Protein in Innate Immunity and Viral Replication" International Journal of Molecular Sciences 24, no. 12: 10056. https://doi.org/10.3390/ijms241210056