
The Role of Annexin A1 in DNA Damage Response in Placental Cells: Impact on Gestational Diabetes Mellitus

 , and
, and
Abstract
:1. Introduction
2. Results
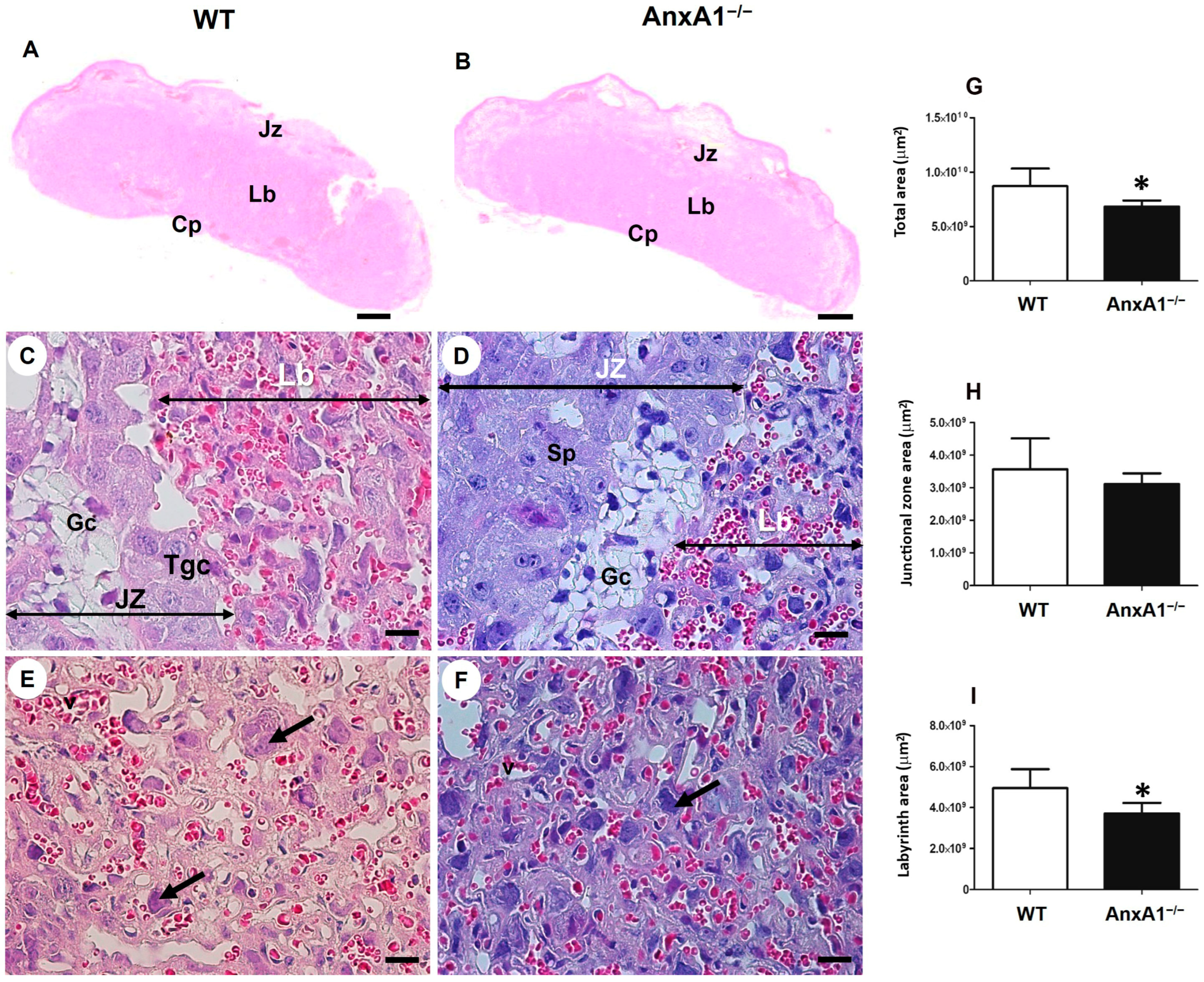
2.1. Deficiency of AnxA1 Gene Modifies the Placental Morphology
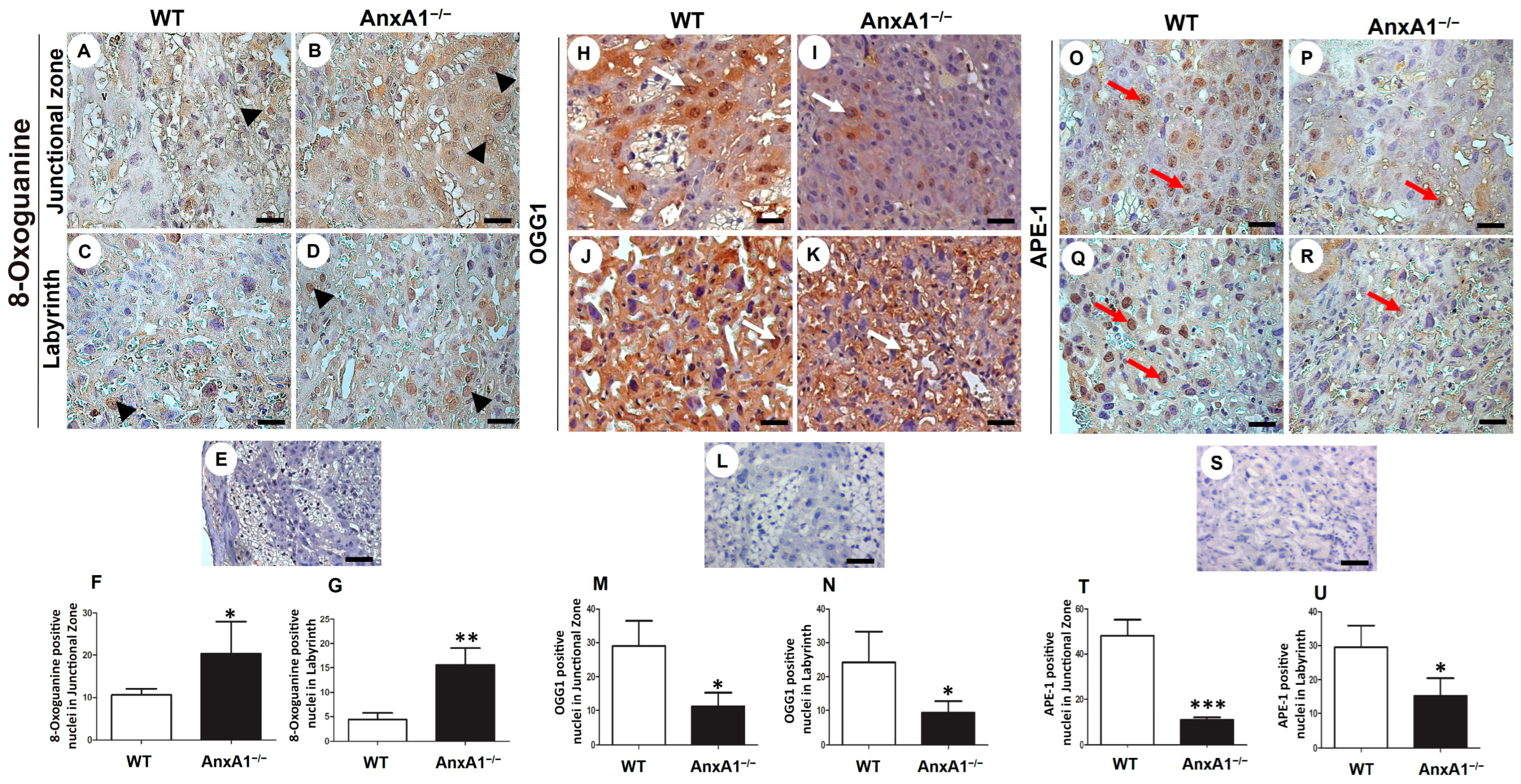
2.2. Oxidative DNA Damage Is Augmented, and the Expression of Repair Enzymes Is Impaired in Placenta from AnxA1−/− Mice
2.3. DNA Double-Strand Breaks and Apoptosis Are Enhanced in Placenta from AnxA1−/− Mice
2.4. GDM Alters Maternal Clinical Parameters
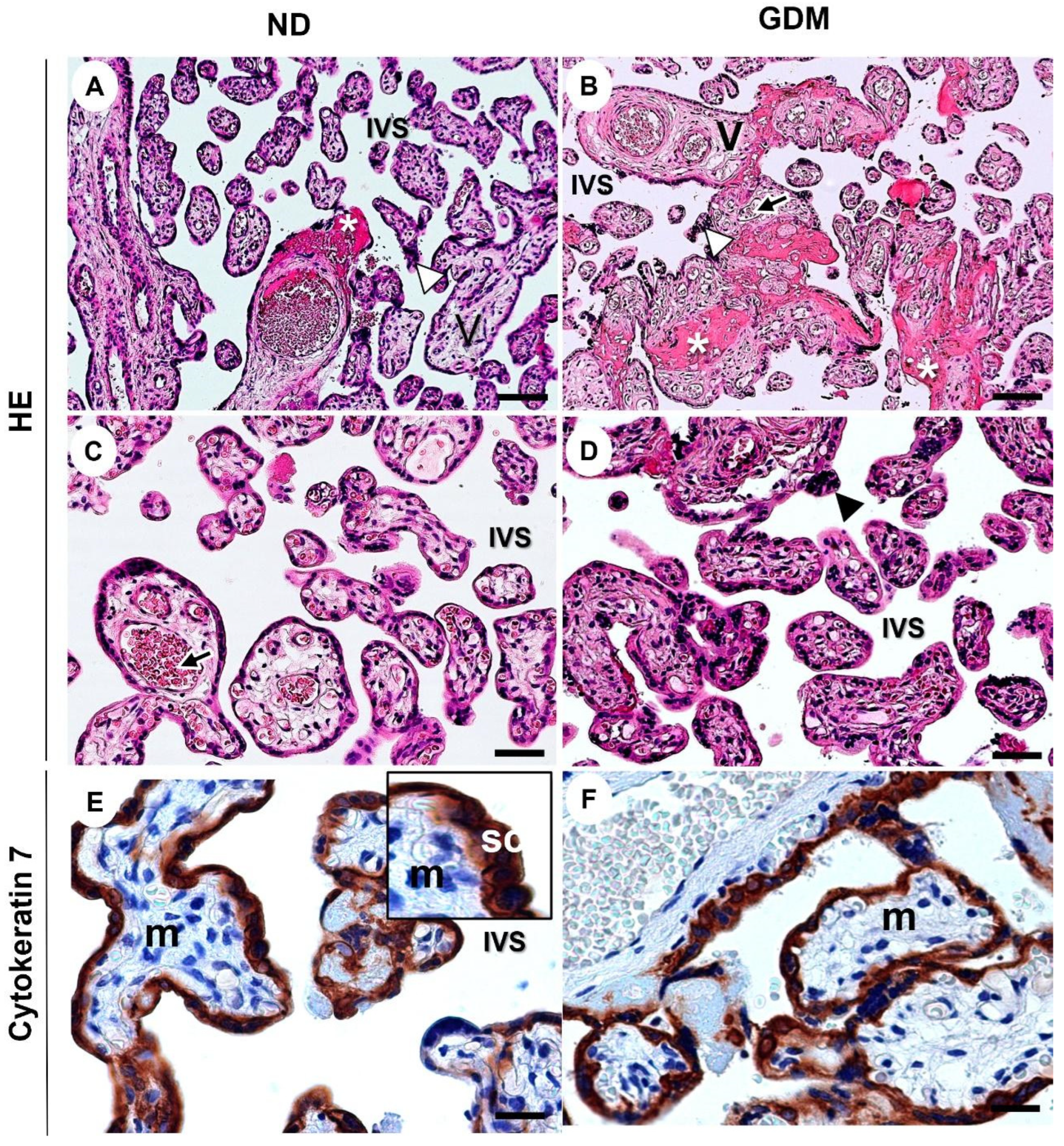
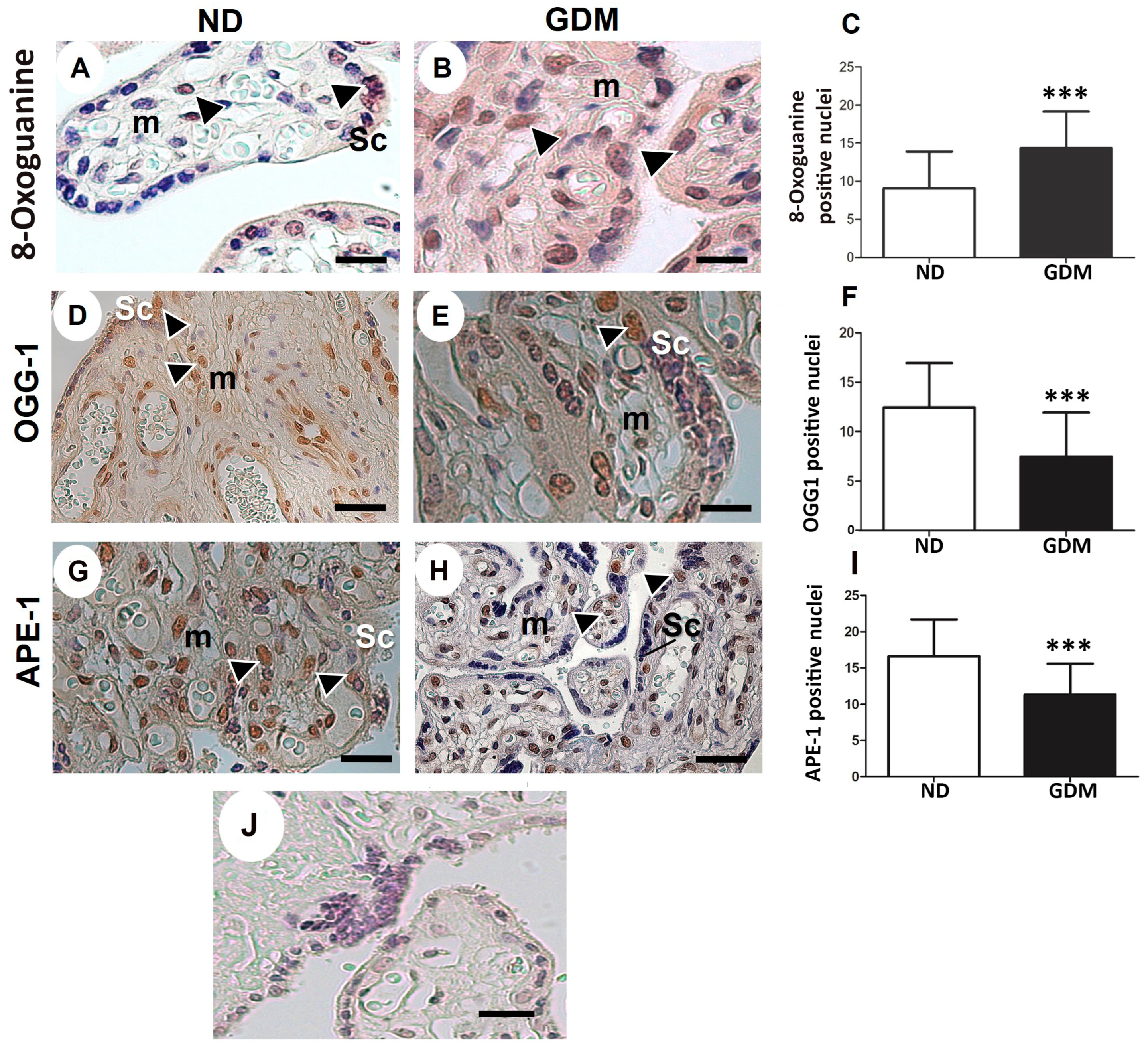
2.5. GDM Changes the Villous Morphology, Leads to Oxidative DNA Damage, and Impairs the Expression of Nuclear DNA Repair Enzymes
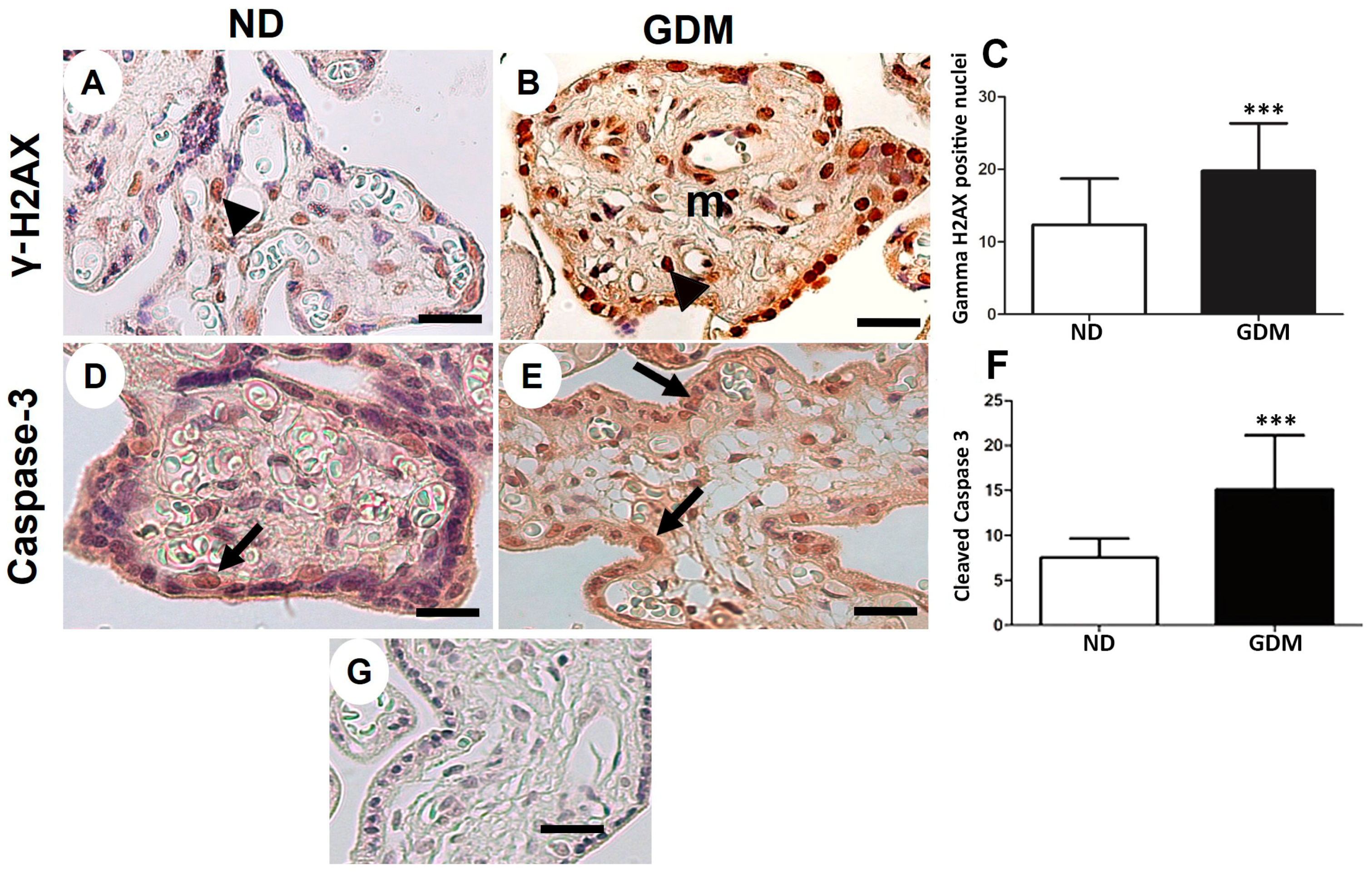
2.6. Placenta from Women with GDM Presents Augmented DNA Double-Strand Breaks and Apoptosis
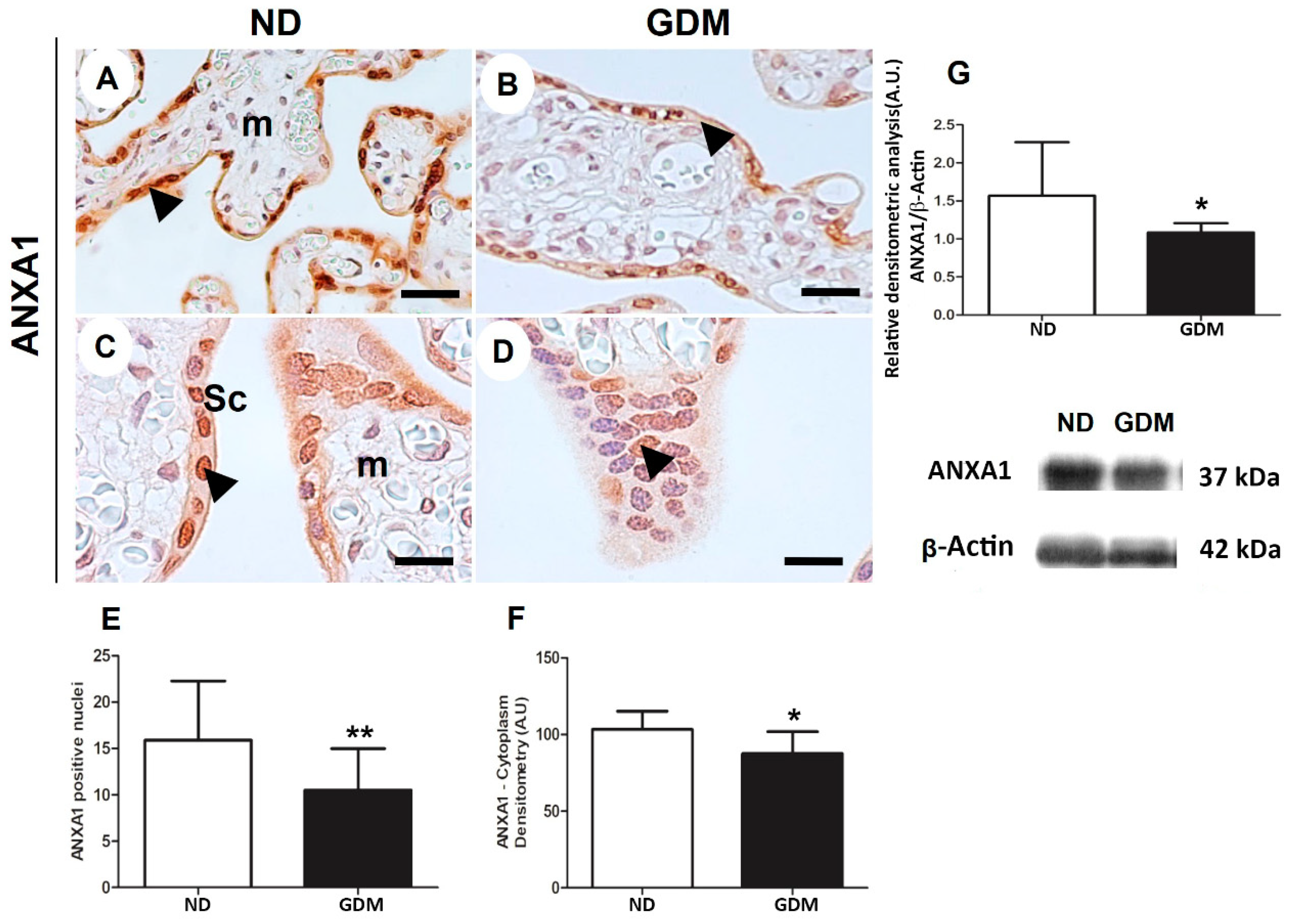
2.7. ANXA1 Expression Is Impaired in the Placenta from Women with GDM
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
4.2. AnxA1 Knockout (AnxA1−/−) Placental Samples
4.3. Population Characterization and Collection of Human Placenta Samples
4.4. Preparation of Placenta Samples for Histological Assessment
4.5. Morphological and Morphometric Analyses of Placenta Samples from AnxA1−/− Mice
4.6. Immunohistochemical Detection of Oxidative DNA Damage, DNA-Double Strand Breaks, DNA Repair Enzymes, and Apoptosis in GDM and AnxA1−/− Placenta Samples
4.7. Nuclear OGG1, APE-1, and Caspase-3 Analysis in Placenta from AnxA1−/− Mice
4.8. Morphological Analysis of Placenta from Pregnant Women with GDM
4.9. Analysis of Nuclear OGG1, APE-1, and Caspase-3 in Placenta from Pregnant Women with GDM
4.10. Detection and Analysis of Cytoplasmic and Nuclear ANXA1 in GDM Placentas
4.11. Western Blot Analysis of ANXA1 Using Placental Extracts from Patients with GDM
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Flower, R.J. Lipocortin and the mechanism of action of the glucocorticoids. Br. J. Pharmacol. 1988, 94, 987–1015. [Google Scholar] [CrossRef] [PubMed]
- Crumpton, M.J.; Dedman, J.R. Protein terminology tangle. Nature 1990, 345, 212. [Google Scholar] [CrossRef] [PubMed]
- Perretti, M.; D’Acquisto, F. Annexin A1 and glucocorticoids as effectors of the resolution of inflammation. Nat. Rev. Immunol. 2009, 9, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.H.; Pervaiz, S. Annexin 1: The new face of an old molecule. FASEB J. 2007, 21, 968–975. [Google Scholar] [CrossRef] [Green Version]
- Chittajallu, R.; Vignes, M.; Dev, K.K.; Barnes, J.M.; Collingridge, G.L.; Henley, J.M. Regulation of glutamate release by presynaptic kainate receptors in the hippocampus. Nature 1996, 379, 78–81. [Google Scholar] [CrossRef]
- Boudhraa, Z.; Bouchon, B.; Viallard, C.; D’Incan, M.; Degoul, F. Annexin A1 localization and its relevance to cancer. Clin. Sci. 2016, 130, 205–220. [Google Scholar] [CrossRef]
- Rhee, H.J.; Kim, G.Y.; Huh, J.W.; Kim, S.W.; Na, D.S. Annexin I is a stress protein induced by heat, oxidative stress and a sulfhydryl-reactive agent. Eur. J. Biochem. 2000, 267, 3220–3225. [Google Scholar] [CrossRef] [Green Version]
- Nair, S.; Hande, M.P.; Lim, L.H. Annexin-1 protects MCF7 breast cancer cells against heat-induced growth arrest and DNA damage. Cancer Lett. 2010, 294, 111–117. [Google Scholar] [CrossRef]
- Hirata, A.; Hirata, F. Lipocortin (Annexin) I heterotetramer binds to purine RNA and pyrimidine DNA. Biochem. Biophys. Res. Commun. 1999, 265, 200–204. [Google Scholar] [CrossRef]
- Hirata, A.; Hirata, F. DNA chain unwinding and annealing reactions of lipocortin (annexin) I heterotetramer: Regulation by Ca2+ and Mg2+. Biochem. Biophys. Res. Commun. 2002, 291, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Hickey, R.; Malkas, L. The biochemical status of the DNA synthesome can distinguish between permanent and temporary cell growth arrest. Cell Growth Differ. 1997, 8, 1359–1369. [Google Scholar] [PubMed]
- Bokhari, B.; Sharma, S. Stress Marks on the Genome: Use or Lose? Int. J. Mol. Sci. 2019, 20, 364. [Google Scholar] [CrossRef] [Green Version]
- Jackson, S.P.; Bartek, J. The DNA-damage response in human biology and disease. Nature 2009, 461, 1071–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreli, J.B.; Santos, J.H.; Rocha, C.R.; Damasceno, D.C.; Morceli, G.; Rudge, M.V.; Bevilacqua, E.; Calderon, I.M. DNA damage and its cellular response in mother and fetus exposed to hyperglycemic environment. BioMed Res. Int. 2014, 2014, 676758. [Google Scholar] [CrossRef] [Green Version]
- Moreli, J.B.; Santos, J.H.; Lorenzon-Ojea, A.R.; Corrêa-Silva, S.; Fortunato, R.S.; Rocha, C.R.; Rudge, M.V.; Damasceno, D.C.; Bevilacqua, E.; Calderon, I.M. Hyperglycemia Differentially Affects Maternal and Fetal DNA Integrity and DNA Damage Response. Int. J. Biol. Sci. 2016, 12, 466–477. [Google Scholar] [CrossRef] [Green Version]
- Whitaker, A.M.; Schaich, M.A.; Smith, M.R.; Flynn, T.S.; Freudenthal, B.D. Base excision repair of oxidative DNA damage: From mechanism to disease. Front. Biosci. 2017, 22, 1493–1522. [Google Scholar]
- American Diabetes Association Professional Practice Committee. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2022. Diabetes Care 2022, 45 (Suppl. 1), S17–S38. [Google Scholar] [CrossRef]
- Desoye, G.; Wells, J.C.K. Pregnancies in Diabetes and Obesity: The Capacity-Load Model of Placental Adaptation. Diabetes 2021, 70, 823–830. [Google Scholar] [CrossRef]
- Damasceno, D.C.; Netto, A.O.; Iessi, I.L.; Gallego, F.Q.; Corvino, S.B.; Dallaqua, B.; Sinzato, Y.K.; Bueno, A.; Calderon, I.M.; Rudge, M.V. Streptozotocin-induced diabetes models: Pathophysiological mechanisms and fetal outcomes. BioMed Res. Int. 2014, 2014, 819065. [Google Scholar] [CrossRef] [Green Version]
- Hoch, D.; Gauster, M.; Hauguel-de Mouzon, S.; Desoye, G. Diabesity-associated oxidative and inflammatory stress signalling in the early human placenta. Mol. Asp. Med. 2019, 66, 21–30. [Google Scholar] [CrossRef]
- Bedell, S.; Hutson, J.; de Vrijer, B.; Eastabrook, G. Effects of Maternal Obesity and Gestational Diabetes Mellitus on the Placenta: Current Knowledge and Targets for Therapeutic Interventions. Curr. Vasc. Pharmacol. 2021, 19, 176–192. [Google Scholar] [CrossRef]
- Sgarbosa, F.; Barbisan, L.F.; Brasil, M.A.; Costa, E.; Calderon, I.M.; Gonçalves, C.R.; Bevilacqua, E.; Rudge, M.V. Changes in apoptosis and Bcl-2 expression in human hyperglycemic, term placental trophoblast. Diabetes Res. Clin. Pract. 2006, 73, 143–149. [Google Scholar] [CrossRef]
- Moreli, J.B.; Corrêa-Silva, S.; Damasceno, D.C.; Sinzato, Y.K.; Lorenzon-Ojea, A.R.; Borbely, A.U.; Rudge, M.V.; Bevilacqua, E.; Calderon, I.M. Changes in the TNF-alpha/IL-10 ratio in hyperglycemia-associated pregnancies. Diabetes Res. Clin. Pract. 2015, 107, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Corrêa-Silva, S.; Alencar, A.P.; Moreli, J.B.; Borbely, A.U.; de SLima, L.; Scavone, C.; Damasceno, D.C.; Rudge, M.V.C.; Bevilacqua, E.; Calderon, I.M.P. Hyperglycemia induces inflammatory mediators in the human chorionic villous. Cytokine 2018, 111, 41–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myatt, L.; Cui, X. Oxidative stress in the placenta. Histochem. Cell Biol. 2004, 122, 369–382. [Google Scholar] [CrossRef]
- Prieto-Fernández, L.; Menéndez, S.T.; Otero-Rosales, M.; Montoro-Jiménez, I.; Hermida-Prado, F.; García-Pedrero, J.M.; Álvarez-Teijeiro, S. Pathobiological functions and clinical implications of annexin dysregulation in human cancers. Front. Cell Dev. Biol. 2022, 10, 1009908. [Google Scholar] [CrossRef]
- Ganesan, T.; Sinniah, A.; Ibrahim, Z.A.; Chik, Z.; Alshawsh, M.A. Annexin A1: A Bane or a Boon in Cancer? A Systematic Review. Molecules 2020, 25, 3700. [Google Scholar] [CrossRef] [PubMed]
- Petrella, A.; Festa, M.; Ercolino, S.F.; Zerilli, M.; Stassi, G.; Solito, E.; Parente, L. Induction of annexin-1 during TRAIL-induced apoptosis in thyroid carcinoma cells. Cell Death Differ. 2005, 12, 1358–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alldridge, L.C.; Bryant, C.E. Annexin 1 regulates cell proliferation by disruption of cell morphology and inhibition of cyclin D1 expression through sustained activation of the ERK1/2 MAPK signal. Exp. Cell Res. 2003, 290, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Oliani, S.M.; Perretti, M. Cell localization of the anti-inflammatory protein annexin 1 during experimental inflammatory response. Ital. J. Anat. Embryol. 2001, 106, 69–77. [Google Scholar]
- de Renty, C.; DePamphilis, M.; Ullah, Z. Cytoplasmic localization of p21 protects trophoblast giant cells from DNA damage induced apoptosis. PLoS ONE 2014, 9, e97434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hay, W.W., Jr. Placental-fetal glucose exchange and fetal glucose metabolism. Trans. Am. Clin. Climatol. Assoc. 2006, 117, 321–340. [Google Scholar] [PubMed]
- Desoye, G.; Cervar-Zivkovic, M. Diabetes Mellitus, Obesity, and the Placenta. Obstet. Gynecol. Clin. N. Am. 2020, 47, 65–79. [Google Scholar]
- de Oliveira Cardoso, M.F.; Moreli, J.B.; Gomes, A.O.; de Freitas Zanon, C.; Silva, A.E.; Paulesu, L.R.; Ietta, F.; Mineo, J.R.; Ferro, E.A.; Oliani, S.M. Annexin A1 peptide is able to induce an anti-parasitic effect in human placental explants infected by Toxoplasma gondii. Microb. Pathog. 2018, 123, 153–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molás, R.B.; Ribeiro, M.R.; Ramalho Dos Santos, M.J.C.; Borbely, A.U.; Oliani, D.V.; Oliani, A.H.; Nadkarni, S.; Nogueira, M.L.; Moreli, J.B.; Oliani, S.M. The involvement of annexin A1 in human placental response to maternal Zika virus infection. Antiviral. Res. 2020, 179, 104809. [Google Scholar] [CrossRef]
- Liao, L.; Yan, W.J.; Tian, C.M.; Li, M.Y.; Tian, Y.Q.; Zeng, G.Q. Knockdown of Annexin A1 Enhances Radioresistance and Inhibits Apoptosis in Nasopharyngeal Carcinoma. Technol. Cancer Res. Treat. 2018, 17, 1533034617750309. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Liu, L.; Li, X.; Chen, W.; Lu, Z. Correction to: Tat-NTS Suppresses the Proliferation, Migration and Invasion of Glioblastoma Cells by Inhibiting Annexin-A1 Nuclear Translocation. Cell. Mol. Neurobiol. 2022, 42, 2727–2732. [Google Scholar] [CrossRef]
- Hebeda, C.B.; Machado, I.D.; Reif-Silva, I.; Moreli, J.B.; Oliani, S.M.; Nadkarni, S.; Perretti, M.; Bevilacqua, E.; Farsky, S.H.P. Endogenous annexin A1 (AnxA1) modulates early-phase gestation and offspring sex-ratio skewing. J. Cell. Physiol. 2018, 233, 6591–6603. [Google Scholar] [CrossRef]
- Meyerholz, D.K.; Beck, A.P. Fundamental Concepts for Semiquantitative Tissue Scoring in Translational Research. ILAR J. 1998, 59, 13–17. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ND (n = 10) | GDM (n = 10) | |
|---|---|---|
| Maternal age (years) | 30.85 ± 6.11 | 29.43 ± 5.36 |
| BMI (Kg/m2) | 34.88 ± 8.44 | 38.05 ± 4.37 |
| Pregnancy weight gain (Kg) | 12.83 ± 7.50 | 12.34 ± 6.94 |
| HbA1c (%) | 5.36 ± 0.63 | 6.32 ± 0.82 * |
| Placental weight (g) | 623.10 ± 78.38 | 632.50 ± 179.80 |
| Newborn weight (g) | 3062.00 ± 276.60 | 3540.00 ± 574.50 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreli, J.B.; Santos, M.R.d.; Calderon, I.d.M.P.; Hebeda, C.B.; Farsky, S.H.P.; Bevilacqua, E.; Oliani, S.M. The Role of Annexin A1 in DNA Damage Response in Placental Cells: Impact on Gestational Diabetes Mellitus. Int. J. Mol. Sci. 2023, 24, 10155. https://doi.org/10.3390/ijms241210155
Moreli JB, Santos MRd, Calderon IdMP, Hebeda CB, Farsky SHP, Bevilacqua E, Oliani SM. The Role of Annexin A1 in DNA Damage Response in Placental Cells: Impact on Gestational Diabetes Mellitus. International Journal of Molecular Sciences. 2023; 24(12):10155. https://doi.org/10.3390/ijms241210155
Chicago/Turabian StyleMoreli, Jusciele Brogin, Mayk Ricardo dos Santos, Iracema de Mattos Paranhos Calderon, Cristina Bichels Hebeda, Sandra Helena Poliselli Farsky, Estela Bevilacqua, and Sonia Maria Oliani. 2023. "The Role of Annexin A1 in DNA Damage Response in Placental Cells: Impact on Gestational Diabetes Mellitus" International Journal of Molecular Sciences 24, no. 12: 10155. https://doi.org/10.3390/ijms241210155