

A Yeast Mitotic Tale for the Nucleus and the Vacuoles to Embrace

 , , and
, , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. The Yeast Nucleus: A Model to Address Nuclear Morphology and Architecture
1.1. Constituents of the Nuclear Envelope with a Role in Shaping the Nucleus
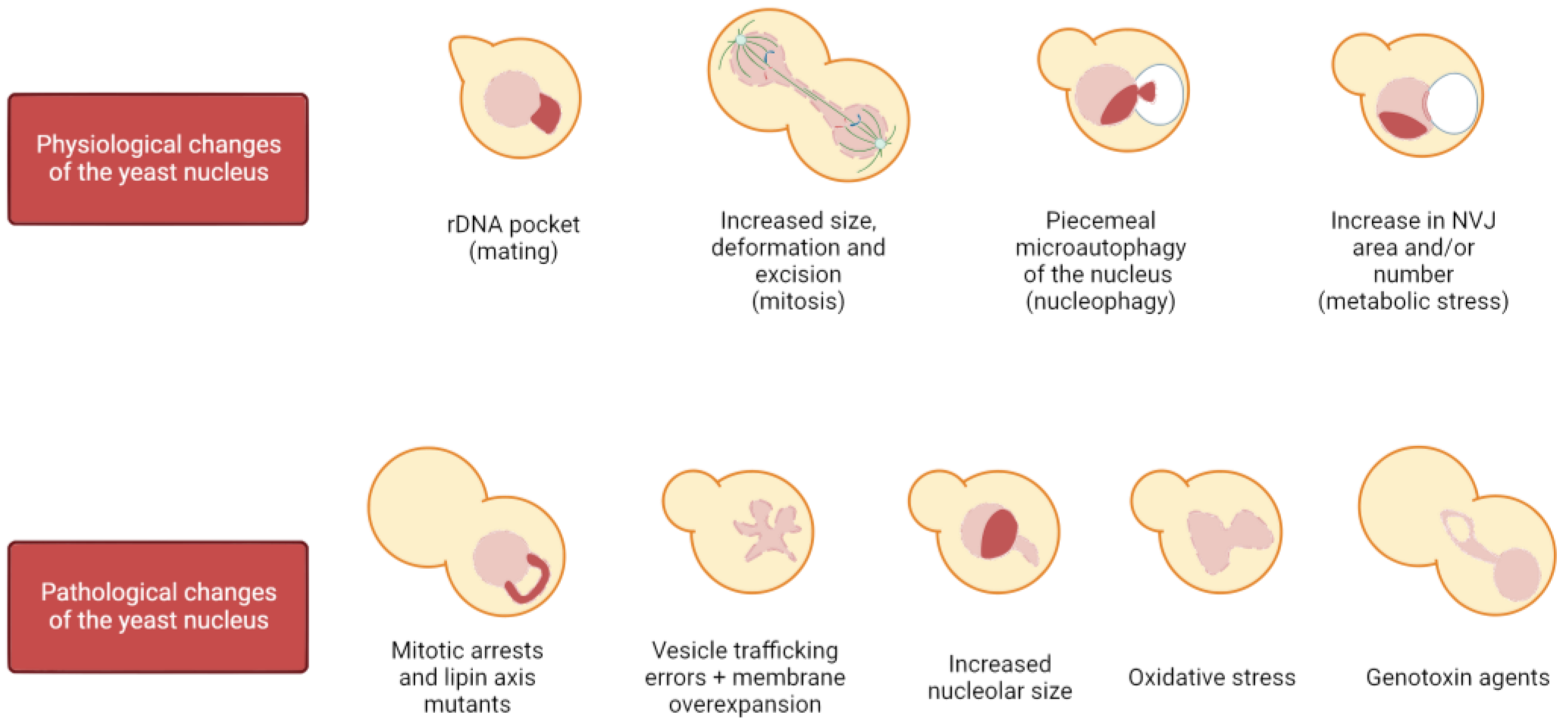
1.2. Physiological Changes in Nuclear Shape and Structure
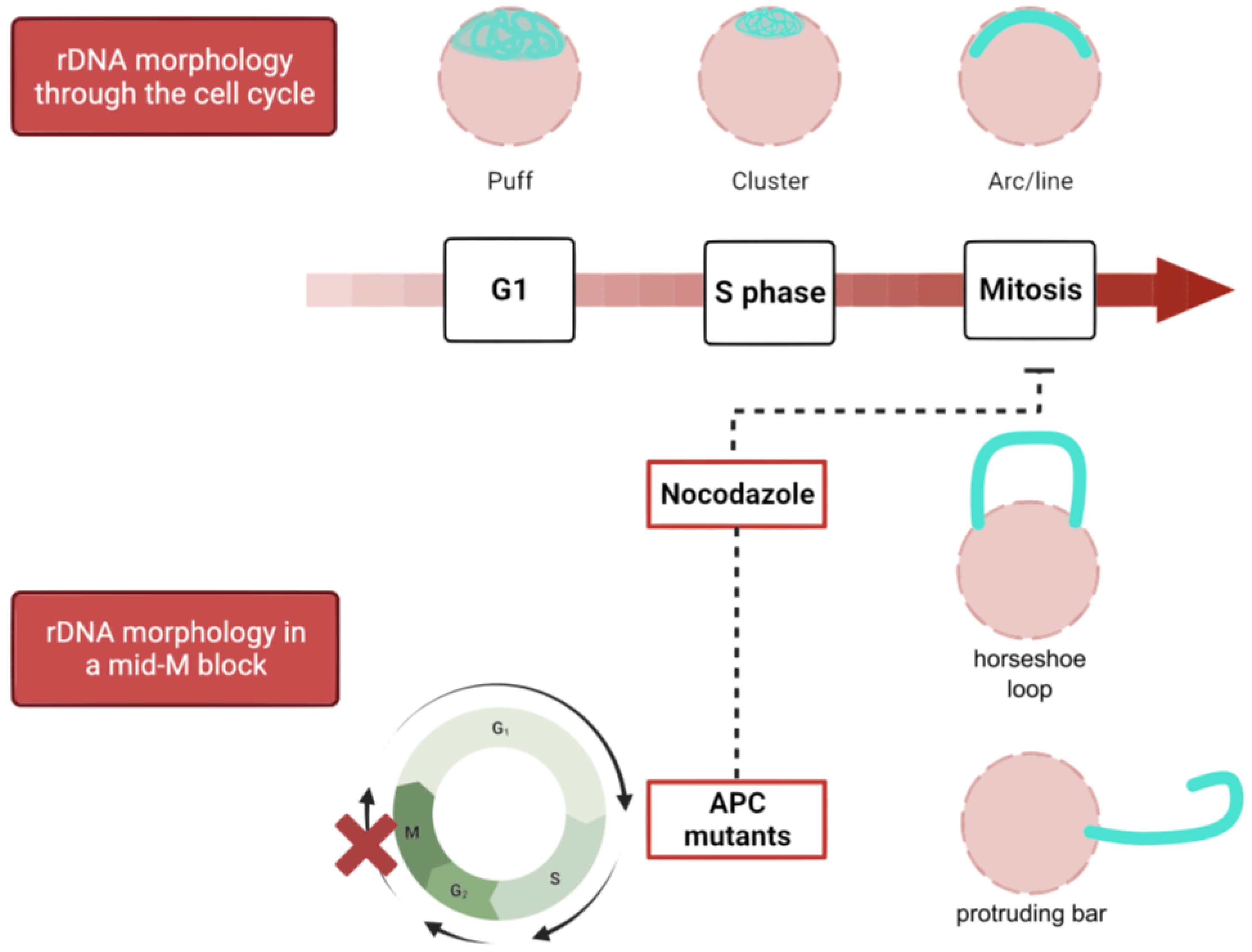
1.2.1. Changes in the Nucleus during Mitosis
1.2.2. Traits of Nuclear Shapes during Mitotic Arrests
2. The Nucleolus and the rDNA
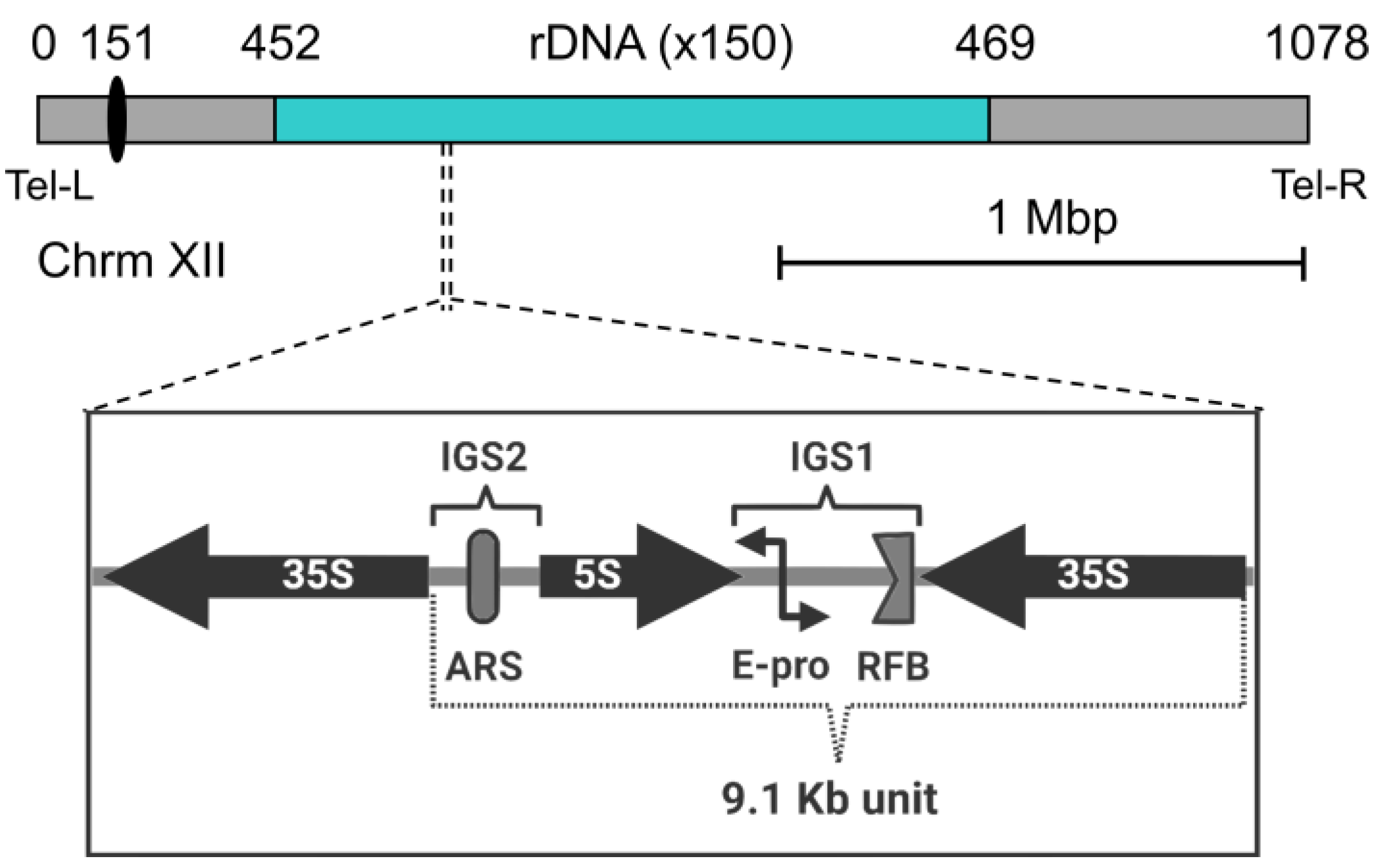
2.1. Features of the Nucleolus and the rDNA in the Budding Yeast
2.2. Shapes of the Nucleolus/rDNA during the Yeast Cell Cycle and NE Expansion
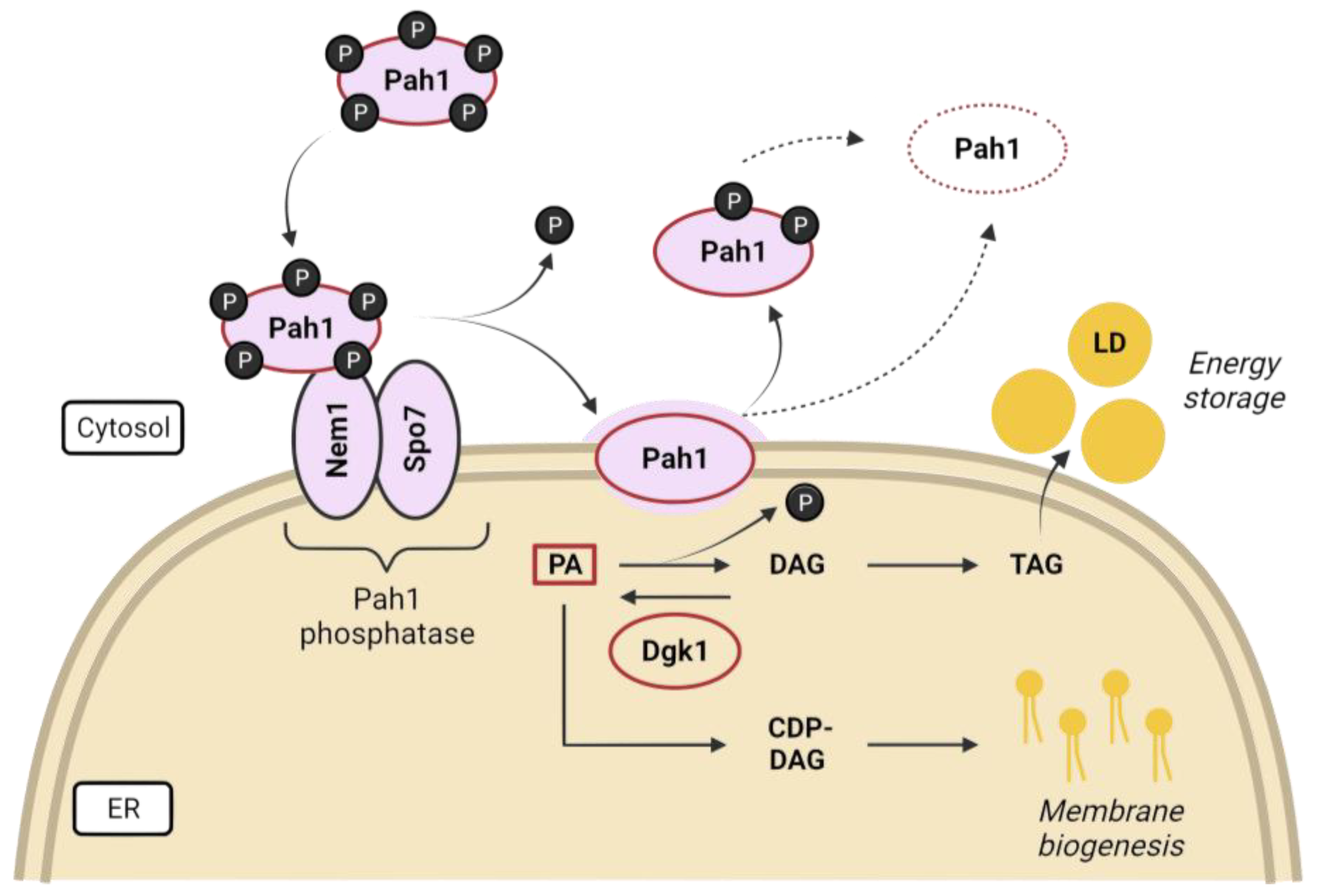
3. Lipid Homeostasis and Its Relationship to Nuclear Shape
4. The Vacuole, Nucleophagy, and TORC1
4.1. The Vacuole
4.2. Nucleophagy
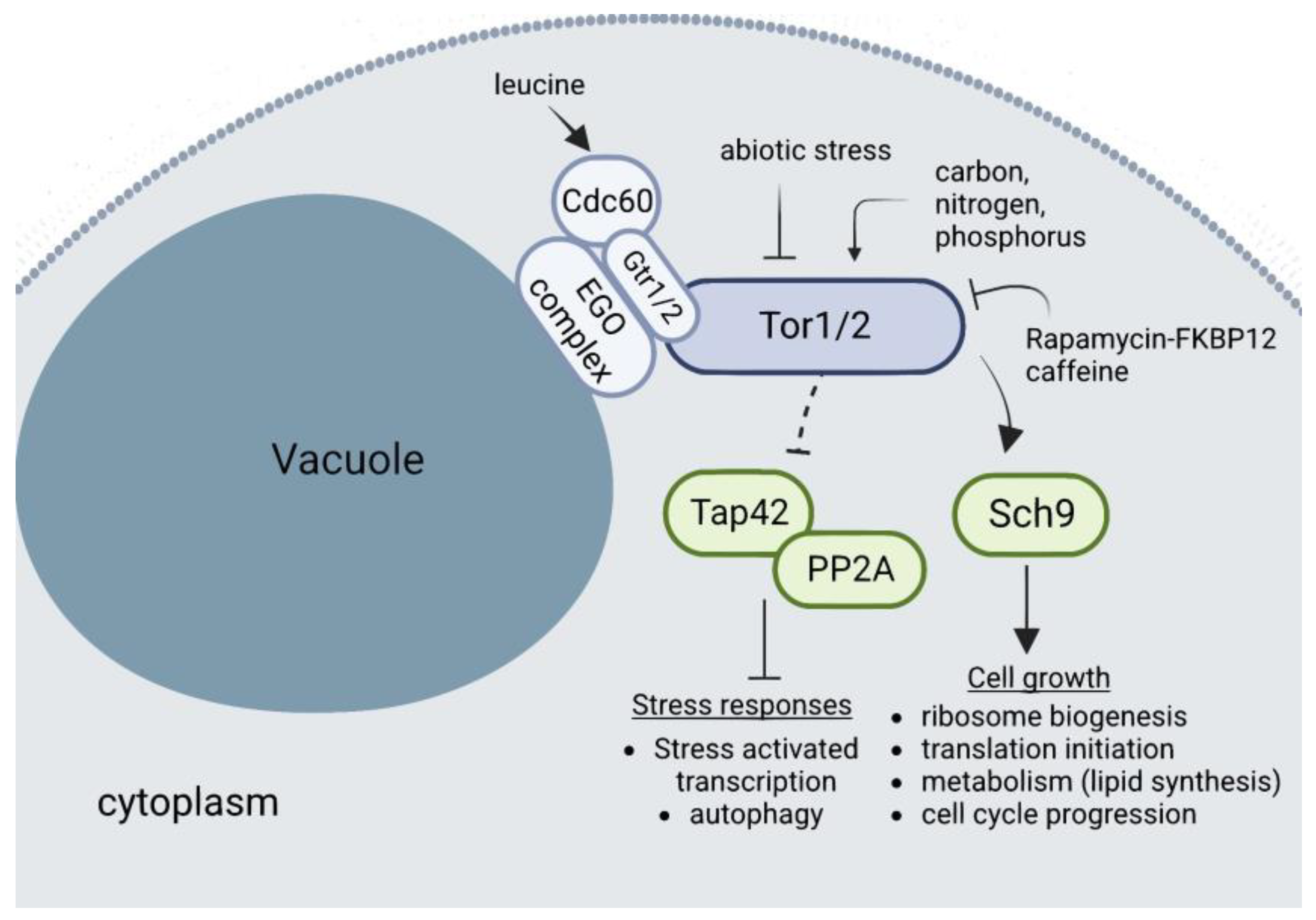
4.3. TORC1
5. Establishing a Morphological Axis Whereby the Vacuole Shapes the Nucleus, Nucleolus, and rDNA during Mitotic Arrests
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Deolal, P.; Mishra, K. Regulation of Diverse Nuclear Shapes: Pathways Working Independently, Together. Commun. Integr. Biol. 2021, 14, 158–175. [Google Scholar] [CrossRef] [PubMed]
- Dechat, T.; Pfleghaar, K.; Sengupta, K.; Shimi, T.; Shumaker, D.K.; Solimando, L.; Goldman, R.D. Nuclear Lamins: Major Factors in the Structural Organization and Function of the Nucleus and Chromatin. Genes Dev. 2008, 22, 832–853. [Google Scholar] [CrossRef] [PubMed]
- Wilhelmsen, K.; Ketema, M.; Truong, H.; Sonnenberg, A. KASH-Domain Proteins in Nuclear Migration, Anchorage and Other Processes. J. Cell Sci. 2006, 119, 5021–5029. [Google Scholar] [CrossRef] [PubMed]
- Hattier, T.; Andrulis, E.D.; Tartakoff, A.M. Immobility, Inheritance and Plasticity of Shape of the Yeast Nucleus. BMC Cell Biol. 2007, 8, 47. [Google Scholar] [CrossRef]
- Andrulis, E.D.; Zappulla, D.C.; Ansari, A.; Perrod, S.; Laiosa, C.V.; Gartenberg, M.R.; Sternglanz, R. Esc1, a Nuclear Periphery Protein Required for Sir4-Based Plasmid Anchoring and Partitioning. Mol. Cell. Biol. 2002, 22, 8292–8301. [Google Scholar] [CrossRef]
- Male, G.; Deolal, P.; Manda, N.K.; Yagnik, S.; Mazumder, A.; Mishra, K. Nucleolar Size Regulates Nuclear Envelope Shape in Saccharomyces cerevisiae. J. Cell Sci. 2020, 133, jcs242172. [Google Scholar] [CrossRef]
- Horigome, C.; Okada, T.; Shimazu, K.; Gasser, S.M.; Mizuta, K. Ribosome Biogenesis Factors Bind a Nuclear Envelope SUN Domain Protein to Cluster Yeast Telomeres. EMBO J. 2011, 30, 3799–3811. [Google Scholar] [CrossRef]
- D’Angelo, M.A.; Hetzer, M.W. Structure, Dynamics and Function of Nuclear Pore Complexes. Trends Cell Biol. 2008, 18, 456–466. [Google Scholar] [CrossRef]
- Alber, F.; Dokudovskaya, S.; Veenhoff, L.M.; Zhang, W.; Kipper, J.; Devos, D.; Suprapto, A.; Karni-Schmidt, O.; Williams, R.; Chait, B.T.; et al. The Molecular Architecture of the Nuclear Pore Complex. Nature 2007, 450, 695–701. [Google Scholar] [CrossRef]
- Rout, M.P.; Aitchison, J.D.; Suprapto, A.; Hjertaas, K.; Zhao, Y.; Chait, B.T. The Yeast Nuclear Pore Complex: Composition, Architecture, and Transport Mechanism. J. Cell Biol. 2000, 148, 635–652. [Google Scholar] [CrossRef]
- Kim, S.J.; Fernandez-Martinez, J.; Nudelman, I.; Shi, Y.; Zhang, W.; Raveh, B.; Herricks, T.; Slaughter, B.D.; Hogan, J.A.; Upla, P.; et al. Integrative Structure and Functional Anatomy of a Nuclear Pore Complex. Nature 2018, 555, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Winey, M.; Yarar, D.; Giddings, T.H.; Mastronarde, D.N. Nuclear Pore Complex Number and Distribution throughout the Saccharomyces cerevisiae Cell Cycle by Three-Dimensional Reconstruction from Electron Micrographs of Nuclear Envelopes. Mol. Biol. Cell 1997, 8, 2119. [Google Scholar] [CrossRef] [PubMed]
- Aitchison, J.D.; Rout, M.P. The Yeast Nuclear Pore Complex and Transport through It. Genetics 2012, 190, 855–883. [Google Scholar] [CrossRef] [PubMed]
- De Magistris, P.; Antonin, W. The Dynamic Nature of the Nuclear Envelope. Curr. Biol. 2018, 28, R487–R497. [Google Scholar] [CrossRef]
- Schmid, M.; Arib, G.; Laemmli, C.; Nishikawa, J.; Durussel, T.; Laemmli, U.K. Nup-PI: The Nucleopore-Promoter Interaction of Genes in Yeast. Mol. Cell 2006, 21, 379–391. [Google Scholar] [CrossRef]
- Nagai, S.; Dubrana, K.; Tsai-Pflugfelder, M.; Davidson, M.B.; Roberts, T.M.; Brown, G.W.; Varela, E.; Hediger, F.; Gasser, S.M.; Krogan, N.J. Functional Targeting of DNA Damage to a Nuclear Pore–Associated SUMO-Dependent Ubiquitin Ligase. Science 2008, 322, 597. [Google Scholar] [CrossRef]
- Gomar-Alba, M.; Pozharskaia, V.; Cichocki, B.; Schaal, C.; Kumar, A.; Jacquel, B.; Charvin, G.; Igual, J.C.; Mendoza, M. Nuclear Pore Complex Acetylation Regulates MRNA Export and Cell Cycle Commitment in Budding Yeast. EMBO J. 2022, 41, e110271. [Google Scholar] [CrossRef]
- Prunuske, A.J.; Ullman, K.S. The Nuclear Envelope: Form and Reformation. Curr. Opin. Cell Biol. 2006, 18, 108–116. [Google Scholar] [CrossRef]
- Ungricht, R.; Kutay, U. Mechanisms and Functions of Nuclear Envelope Remodelling. Nat. Rev. Mol. Cell Biol. 2017, 18, 229–245. [Google Scholar] [CrossRef]
- Guacci, V.; Hogan, E.; Koshland, D. Centromere Position in Budding Yeast: Evidence for Anaphase A. Mol. Biol. Cell 1997, 8, 957. [Google Scholar] [CrossRef]
- Jin, Q.W.; Trelles-Sticken, E.; Scherthan, H.; Loidl, J. Yeast Nuclei Display Prominent Centromere Clustering That Is Reduced in Nondividing Cells and in Meiotic Prophase. J. Cell Biol. 1998, 141, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Heun, P.; Laroche, T.; Shimada, K.; Furrer, P.; Gasser, S.M. Chromosome Dynamics in the Yeast Interphase Nucleus. Science 2001, 294, 2181–2186. [Google Scholar] [CrossRef] [PubMed]
- Bystricky, K.; Heun, P.; Gehlen, L.; Langowski, J.; Gasser, S.M. Long-Range Compaction and Flexibility of Interphase Chromatin in Budding Yeast Analyzed by High-Resolution Imaging Techniques. Proc. Natl. Acad. Sci. USA 2004, 101, 16495–16500. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.; Klee, S.K.; Evangelista, M.; Boone, C.; Pellman, D. Control of Mitotic Spindle Position by the Saccharomyces cerevisiae Formin Bni1p. J. Cell Biol. 1999, 144, 947–961. [Google Scholar] [CrossRef]
- Yang, C.H.; Lambie, E.J.; Hardin, J.; Craft, J.; Snyder, M. Higher Order Structure Is Present in the Yeast Nucleus: Autoantibody Probes Demonstrate That the Nucleolus Lies Opposite the Spindle Pole Body. Chromosoma 1989, 98, 123–128. [Google Scholar] [CrossRef]
- Bystricky, K.; Laroche, T.; van Houwe, G.; Blaszczyk, M.; Gasser, S.M. Chromosome Looping in Yeast: Telomere Pairing and Coordinated Movement Reflect Anchoring Efficiency and Territorial Organization. J. Cell Biol. 2005, 168, 375–387. [Google Scholar] [CrossRef]
- Jaspersen, S.L.; Winey, M. The Budding Yeast Spindle Pole Body: Structure, Duplication, and Function. Annu. Rev. Cell Dev. Biol. 2004, 20, 1–28. [Google Scholar] [CrossRef]
- Jaspersen, S.L.; Giddings, T.H.; Winey, M. Mps3p Is a Novel Component of the Yeast Spindle Pole Body That Interacts with the Yeast Centrin Homologue Cdc31p. J. Cell Biol. 2002, 159, 945–956. [Google Scholar] [CrossRef]
- Bupp, J.M.; Martin, A.E.; Stensrud, E.S.; Jaspersen, S.L. Telomere Anchoring at the Nuclear Periphery Requires the Budding Yeast Sad1-UNC-84 Domain Protein Mps3. J. Cell Biol. 2007, 179, 845. [Google Scholar] [CrossRef]
- Fan, J.; Jin, H.; Koch, B.A.; Yu, H.-G. Mps2 Links Csm4 and Mps3 to Form a Telomere-Associated LINC Complex in Budding Yeast. Life Sci. Alliance 2020, 3. [Google Scholar] [CrossRef]
- Chan, J.N.Y.; Poon, B.P.K.; Salvi, J.; Olsen, J.B.; Emili, A.; Mekhail, K. Perinuclear Cohibin Complexes Maintain Replicative Life Span via Roles at Distinct Silent Chromatin Domains. Dev. Cell 2011, 20, 867–879. [Google Scholar] [CrossRef] [PubMed]
- Taddei, A.; Gasser, S.M. Structure and Function in the Budding Yeast Nucleus. Genetics 2012, 192, 107–129. [Google Scholar] [CrossRef] [PubMed]
- Banday, S.; Farooq, Z.; Rashid, R.; Abdullah, E.; Altaf, M. Role of Inner Nuclear Membrane Protein Complex Lem2-Nur1 in Heterochromatic Gene Silencing. J. Biol. Chem. 2016, 291, 20021–20029. [Google Scholar] [CrossRef] [PubMed]
- Webster, B.M.; Thaller, D.J.; Jäger, J.; Ochmann, S.E.; Borah, S.; Lusk, C.P. Chm7 and Heh1 Collaborate to Link Nuclear Pore Complex Quality Control with Nuclear Envelope Sealing. EMBO J. 2016, 35, 2447–2467. [Google Scholar] [CrossRef]
- Webster, B.M.; Colombi, P.; Jäger, J.; Patrick Lusk, C. Surveillance of Nuclear Pore Complex Assembly by ESCRT-III/Vps4. Cell 2014, 159, 388–401. [Google Scholar] [CrossRef] [PubMed]
- Mekhail, K.; Seebacher, J.; Gygi, S.P.; Moazed, D. Role for Perinuclear Chromosome Tethering in Maintenance of Genome Stability. Nature 2008, 456, 667–670. [Google Scholar] [CrossRef]
- Cavanaugh, A.M.; Jaspersen, S.L. Big Lessons from Little Yeast: Budding and Fission Yeast Centrosome Structure, Duplication, and Function. Annu. Rev. Genet. 2017, 51, 361–383. [Google Scholar] [CrossRef]
- Fan, J.; Sun, Z.; Wang, Y. The Assembly of a Noncanonical LINC Complex in Saccharomyces cerevisiae. Curr. Genet. 2022, 68, 91–96. [Google Scholar] [CrossRef]
- Sosa Ponce, M.L.; Moradi-Fard, S.; Zaremberg, V.; Cobb, J.A. SUNny Ways: The Role of the SUN-Domain Protein Mps3 Bridging Yeast Nuclear Organization and Lipid Homeostasis. Front. Genet. 2020, 11, 136. [Google Scholar] [CrossRef]
- Wente, S.R.; Blobel, G. NUP145 Encodes a Novel Yeast Glycine-Leucine-Phenylalanine-Glycine (GLFG) Nucleoporin Required for Nuclear Envelope Structure. J. Cell Biol. 1994, 125, 955–969. [Google Scholar] [CrossRef]
- Mekhail, K.; Moazed, D. The Nuclear Envelope in Genome Organization, Expression and Stability. Nat. Rev. Mol. Cell Biol. 2010, 11, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Phillip, J.M.; Aifuwa, I.; Walston, J.; Wirtz, D. The Mechanobiology of Aging. Annu. Rev. Biomed. Eng. 2015, 17, 113–141. [Google Scholar] [CrossRef] [PubMed]
- Zink, D.; Fischer, A.H.; Nickerson, J.A. Nuclear Structure in Cancer Cells. Nat. Rev. Cancer 2004, 4, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Scaffidi, P.; Misteli, T. Lamin A-Dependent Nuclear Defects in Human Aging. Science 2006, 312, 1059. [Google Scholar] [CrossRef]
- Denholtz, M.; Zhu, Y.; He, Z.; Lu, H.; Isoda, T.; Döhrmann, S.; Nizet, V.; Murre, C. Upon Microbial Challenge, Human Neutrophils Undergo Rapid Changes in Nuclear Architecture and Chromatin Folding to Orchestrate an Immediate Inflammatory Gene Program. Genes Dev. 2020, 34, 149–165. [Google Scholar] [CrossRef]
- Rowat, A.C.; Jaalouk, D.E.; Zwerger, M.; Ung, W.L.; Eydelnant, I.A.; Olins, D.E.; Olins, A.L.; Herrmann, H.; Weitz, D.A.; Lammerding, J. Nuclear Envelope Composition Determines the Ability of Neutrophil-Type Cells to Passage through Micron-Scale Constrictions. J. Biol. Chem. 2013, 288, 8610–8618. [Google Scholar] [CrossRef]
- Manley, H.R.; Keightley, M.C.; Lieschke, G.J. The Neutrophil Nucleus: An Important Influence on Neutrophil Migration and Function. Front. Immunol. 2018, 9, 2867. [Google Scholar] [CrossRef]
- Hoffmann, K.; Dreger, C.K.; Olins, A.L.; Olins, D.E.; Shultz, L.D.; Lucke, B.; Karl, H.; Kaps, R.; Müller, D.; Vayá, A.; et al. Mutations in the Gene Encoding the Lamin B Receptor Produce an Altered Nuclear Morphology in Granulocytes (Pelger–Huët Anomaly). Nat. Genet. 2002, 31, 410–414. [Google Scholar] [CrossRef]
- Stone, E.M.; Heun, P.; Laroche, T.; Pillus, L.; Gasser, S.M. MAP Kinase Signaling Induces Nuclear Reorganization in Budding Yeast. Curr. Biol. 2000, 10, 373–382. [Google Scholar] [CrossRef]
- Laporte, D.; Courtout, F.; Salin, B.; Ceschin, J.; Sagot, I. An Array of Nuclear Microtubules Reorganizes the Budding Yeast Nucleus during Quiescence. J. Cell Biol. 2013, 203, 585–594. [Google Scholar] [CrossRef]
- Wang, R.; Kamgoue, A.; Normand, C.; Léger-Silvestre, I.; Mangeat, T.; Gadal, O. High Resolution Microscopy Reveals the Nuclear Shape of Budding Yeast during Cell Cycle and in Various Biological States. J. Cell Sci. 2016, 129, 4480–4495. [Google Scholar] [CrossRef] [PubMed]
- Witkin, K.L.; Chong, Y.; Shao, S.; Webster, M.T.; Lahiri, S.; Walters, A.D.; Lee, B.; Koh, J.L.Y.; Prinz, W.A.; Andrews, B.J.; et al. The Budding Yeast Nuclear Envelope Adjacent to the Nucleolus Serves as a Membrane Sink during Mitotic Delay. Curr. Biol. 2012, 22, 1128–1133. [Google Scholar] [CrossRef] [PubMed]
- Walters, A.D.; May, C.K.; Dauster, E.S.; Cinquin, B.P.; Smith, E.A.; Robellet, X.; D’Amours, D.; Larabell, C.A.; Cohen-Fix, O. The Yeast Polo Kinase Cdc5 Regulates the Shape of the Mitotic Nucleus. Curr. Biol. 2014, 24, 2861–2867. [Google Scholar] [CrossRef] [PubMed]
- Meseroll, R.A.; Cohen-Fix, O. The Malleable Nature of the Budding Yeast Nuclear Envelope: Flares, Fusion, and Fenestrations. J. Cell. Physiol. 2016, 231, 2353–2360. [Google Scholar] [CrossRef]
- Güttinger, S.; Laurell, E.; Kutay, U. Orchestrating Nuclear Envelope Disassembly and Reassembly during Mitosis. Nat. Rev. Mol. Cell Biol. 2009, 10, 178–191. [Google Scholar] [CrossRef]
- De Souza, C.P.C.; Osmani, S.A. Mitosis, Not Just Open or Closed. Eukaryot. Cell 2007, 6, 1521–1527. [Google Scholar] [CrossRef]
- Expósito-Serrano, M.; Sánchez-Molina, A.; Gallardo, P.; Salas-Pino, S.; Daga, R.R. Selective Nuclear Pore Complex Removal Drives Nuclear Envelope Division in Fission Yeast. Curr. Biol. 2020, 30, 3212–3222.e2. [Google Scholar] [CrossRef]
- Dey, G.; Culley, S.; Curran, S.; Schmidt, U.; Henriques, R.; Kukulski, W.; Baum, B. Closed Mitosis Requires Local Disassembly of the Nuclear Envelope. Nature 2020, 585, 119–123. [Google Scholar] [CrossRef]
- Liu, S.; Kwon, M.; Mannino, M.; Yang, N.; Renda, F.; Khodjakov, A.; Pellman, D. Nuclear Envelope Assembly Defects Link Mitotic Errors to Chromothripsis. Nature 2018, 561, 551–555. [Google Scholar] [CrossRef]
- Raab, M.; Gentili, M.; de Belly, H.; Thiam, H.-R.; Vargas, P.; Jimenez, A.J.; Lautenschlaeger, F.; Voituriez, R.; Lennon-Duménil, A.-M.; Manel, N.; et al. ESCRT III Repairs Nuclear Envelope Ruptures during Cell Migration to Limit DNA Damage and Cell Death. Science 2016, 352, 359–362. [Google Scholar] [CrossRef]
- Varshney, N.; Sanyal, K. Nuclear Migration in Budding Yeasts: Position before Division. Curr. Genet. 2019, 65, 1341–1346. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.M. The Anaphase Promoting Complex/Cyclosome: A Machine Designed to Destroy. Nat. Rev. Mol. Cell Biol. 2006, 7, 644–656. [Google Scholar] [CrossRef] [PubMed]
- Michaelis, C.; Ciosk, R.; Nasmyth, K. Cohesins: Chromosomal Proteins That Prevent Premature Separation of Sister Chromatids. Cell 1997, 91, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Etemad, B.; Kops, G.J. Attachment Issues: Kinetochore Transformations and Spindle Checkpoint Silencing. Curr. Opin. Cell Biol. 2016, 39, 101–108. [Google Scholar] [CrossRef]
- Machín, F.; Torres-Rosell, J.; Jarmuz, A.; Aragón, L. Spindle-Independent Condensation-Mediated Segregation of Yeast Ribosomal DNA in Late Anaphase. J. Cell Biol. 2005, 168, 209–219. [Google Scholar] [CrossRef]
- Guacci, V.; Hogan, E.; Koshland, D. Chromosome Condensation and Sister Chromatid Pairing in Budding Yeast. J. Cell Biol. 1994, 125, 517. [Google Scholar] [CrossRef]
- Palmer, R.E.; Koval, M.; Koshland, D. The Dynamics of Chromosome Movement in the Budding Yeast Saccharomyces Cerevisiae. J. Cell Biol. 1989, 109, 3355–3366. [Google Scholar] [CrossRef]
- Palmer, R.E.; Sullivan, D.S.; Huffaker, T.; Koshland, D. Role of Astral Microtubules and Actin in Spindle Orientation and Migration in the Budding Yeast, Saccharomyces Cerevisiae. J. Cell Biol. 1992, 119, 583–593. [Google Scholar] [CrossRef]
- McStay, B. Nucleolar Organizer Regions: Genomic “dark Matter” Requiring Illumination. Genes Dev. 2016, 30, 1598–1610. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nagata, S. Ribosomal RNA Gene Repeats, Their Stability and Cellular Senescence. Proc. Jpn. Acad. Ser. B 2014, 90, 119–129. [Google Scholar] [CrossRef]
- Toussaint, M.; Levasseur, G.; Tremblay, M.; Paquette, M.; Conconi, A. Psoralen Photocrosslinking, a Tool to Study the Chromatin Structure of RNA Polymerase I—Transcribed Ribosomal Genes. Biochem. Cell Biol. 2005, 83, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Pasero, P.; Marilley, M. Size Variation of RDNA Clusters in the Yeasts Saccharomyces cerevisiae and Schizosaccharomyces pombe. Mol. Gen. Genet. MGG 1993, 236, 448–452. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, B.D.; Hogan, E.; Koshland, D. In Vivo Requirements for RDNA Chromosome Condensation Reveal Two Cell-Cycle-Regulated Pathways for Mitotic Chromosome Folding. Genes Dev. 2004, 18, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, B.D.; Hogan, E.; Koshland, D. In Vivo Dissection of the Chromosome Condensation Machinery: Reversibility of Condensation Distinguishes Contributions of Condensin and Cohesin. J. Cell Biol. 2002, 156, 805. [Google Scholar] [CrossRef]
- Machín, F.; Quevedo, O.; Ramos-Pérez, C.; García-Luis, J. Cdc14 Phosphatase: Warning, No Delay Allowed for Chromosome Segregation! Curr. Genet. 2016, 62, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Blanco, A.; Mayán-Santos, M.; Schneider, D.A.; Machín, F.; Jarmuz, A.; Tschochner, H.; Aragón, L. Cdc14 Inhibits Transcription by RNA Polymerase I during Anaphase. Nature 2009, 458, 219–222. [Google Scholar] [CrossRef]
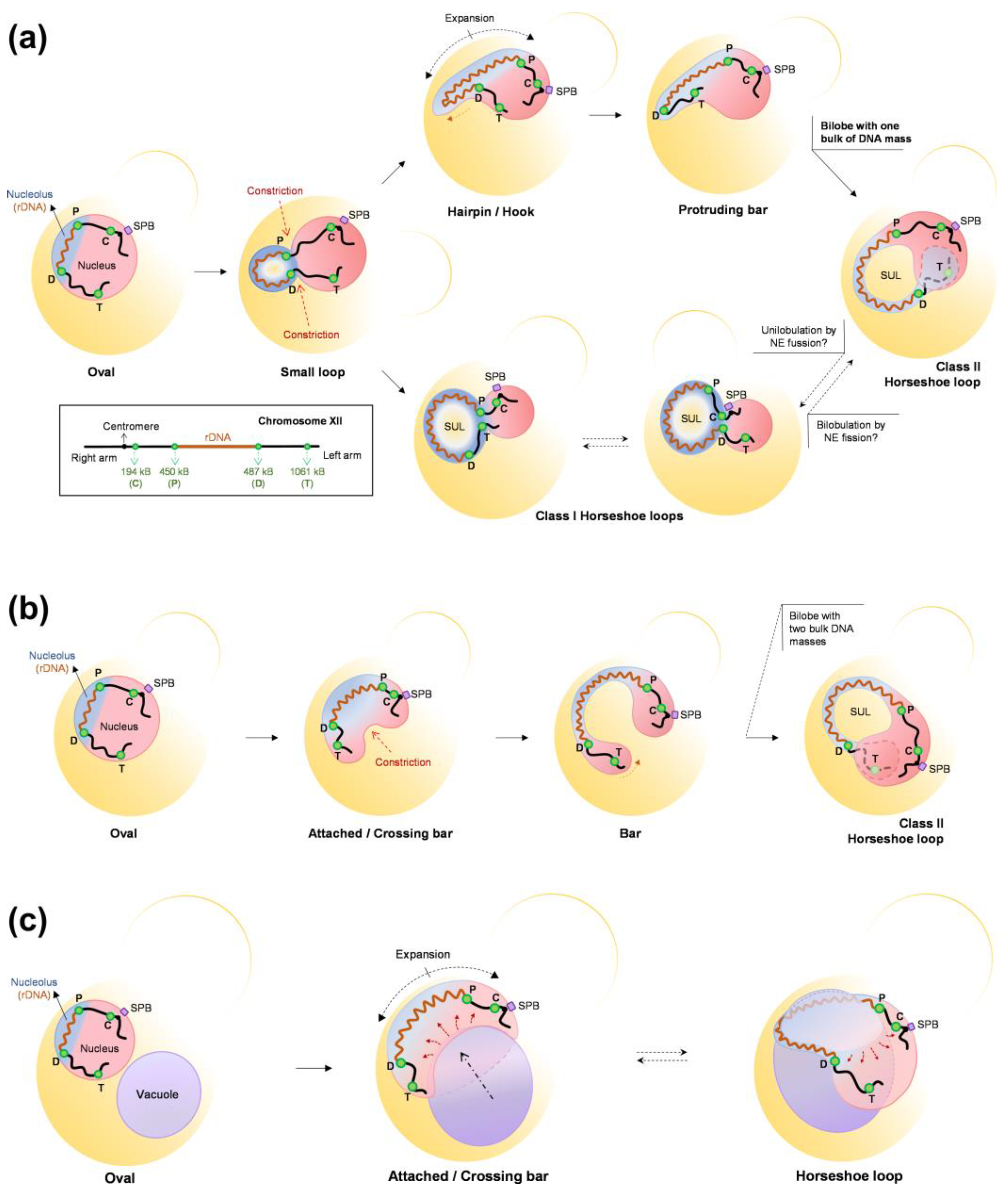
- Machín, F.; Torres-Rosell, J.; De Piccoli, G.; Carballo, J.A.; Cha, R.S.; Jarmuz, A.; Aragón, L. Transcription of Ribosomal Genes Can Cause Nondisjunction. J. Cell Biol. 2006, 173, 893–903. [Google Scholar] [CrossRef]
- Wang, B.D.; Butylin, P.; Strunnikov, A. Condensin Function in Mitotic Nucleolar Segregation Is Regulated by RDNA Transcription. Cell Cycle 2006, 5, 2260–2267. [Google Scholar] [CrossRef]
- Kobayashi, T.; Horiuchi, T. A Yeast Gene Product, Fob1 Protein, Required for Both Replication Fork Blocking and Recombinational Hotspot Activities. Genes Cells 1996, 1, 465–474. [Google Scholar] [CrossRef]
- Sinclair, D.A.; Guarente, L. Extrachromosomal RDNA Circles—A Cause of Aging in Yeast. Cell 1997, 91, 1033–1042. [Google Scholar] [CrossRef]
- Petes, T.D. Unequal Meiotic Recombination within Tandem Arrays of Yeast Ribosomal DNA Genes. Cell 1980, 19, 765–774. [Google Scholar] [CrossRef] [PubMed]
- Torres-Rosell, J.; Sunjevaric, I.; De Piccoli, G.; Sacher, M.; Eckert-Boulet, N.; Reid, R.; Jentsch, S.; Rothstein, R.; Aragón, L.; Lisby, M. The Smc5-Smc6 Complex and SUMO Modification of Rad52 Regulates Recombinational Repair at the Ribosomal Gene Locus. Nat. Cell Biol. 2007, 9, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, T.; Kobayashi, T. Visualization of the Dynamic Behavior of Ribosomal RNA Gene Repeats in Living Yeast Cells. Genes Cells 2011, 16, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Torres-Rosell, J.; Machín, F.; Aragón, L. Cdc14 and the Temporal Coordination between Mitotic Exit and Chromosome Segregation. Cell Cycle 2005, 4, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Ayra-Plasencia, J.; Machín, F. DNA Double-Strand Breaks in Telophase Lead to Coalescence between Segregated Sister Chromatid Loci. Nat. Commun. 2019, 10, 2862. [Google Scholar] [CrossRef]
- Freeman, L.; Aragon-Alcaide, L.; Strunnikov, A. The Condensin Complex Governs Chromosome Condensation and Mitotic Transmission of Rdna. J. Cell Biol. 2000, 149, 811–824. [Google Scholar] [CrossRef]
- Matos-Perdomo, E.; Machín, F. The Ribosomal DNA Metaphase Loop of Saccharomyces cerevisiae Gets Condensed upon Heat Stress in a Cdc14-Independent TORC1-Dependent Manner. Cell Cycle 2018, 17, 200–215. [Google Scholar] [CrossRef]
- Harris, B.; Bose, T.; Lee, K.K.; Wang, F.; Lu, S.; Ross, R.T.; Zhang, Y.; French, S.L.; Beyer, A.L.; Slaughter, B.D.; et al. Cohesion Promotes Nucleolar Structure and Function. Mol. Biol. Cell 2014, 25, 337–346. [Google Scholar] [CrossRef]
- Lamothe, R.; Costantino, L.; Koshland, D.E. The Spatial Regulation of Condensin Activity in Chromosome Condensation. Genes Dev. 2020, 34, 819–831. [Google Scholar] [CrossRef]
- St-Pierre, J.; Douziech, M.; Bazile, F.; Pascariu, M.; Bonneil, É.; Sauvé, V.; Ratsima, H.; D’Amours, D. Polo Kinase Regulates Mitotic Chromosome Condensation by Hyperactivation of Condensin DNA Supercoiling Activity. Mol. Cell 2009, 34, 416–426. [Google Scholar] [CrossRef]
- Li, H.; Tsang, C.K.; Watkins, M.; Bertram, P.G.; Zheng, X.F.S. Nutrient Regulates Tor1 Nuclear Localization and Association with RDNA Promoter. Nature 2006, 442, 1058–1061. [Google Scholar] [CrossRef]
- Martin, D.E.; Powers, T.; Hall, M.N. Regulation of Ribosome Biogenesis: Where Is TOR? Cell Metab. 2006, 4, 259–260. [Google Scholar] [CrossRef] [PubMed]
- Tsang, C.K.; Bertram, P.G.; Ai, W.; Drenan, R.; Zheng, X.F.S. Chromatin-Mediated Regulation of Nucleolar Structure and RNA Pol I Localization by TOR. EMBO J. 2003, 22, 6045–6056. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, A.D.; Lim, K.; Mari, M.; Edgar, J.R.; Gal, L.; Sterk, P.; Jenkins, B.J.; Koulman, A.; Savage, D.B.; Schuldiner, M.; et al. Compartmentalized Synthesis of Triacylglycerol at the Inner Nuclear Membrane Regulates Nuclear Organization. Dev. Cell 2019, 50, 755–766.e6. [Google Scholar] [CrossRef]
- Campbell, J.L.; Lorenz, A.; Witkin, K.L.; Hays, T.; Loidl, J.; Cohen-Fix, O. Yeast Nuclear Envelope Subdomains with Distinct Abilities to Resist Membrane Expansion. Mol. Biol. Cell 2006, 17, 1768–1778. [Google Scholar] [CrossRef] [PubMed]
- Makarova, M.; Gu, Y.; Chen, J.-S.; René, E.; Beckley, J.; Gould, K.L. Temporal Regulation of Lipin Activity Diverged to Account for Differences in Mitotic Programs. Curr. Biol. 2016, 26, 237–243. [Google Scholar] [CrossRef]
- Garcia, M.; Kumanski, S.; Elías-Villalobos, A.; Cazevieille, C.; Soulet, C.; Moriel-Carretero, M. Nuclear Ingression of Cytoplasmic Bodies Accompanies a Boost in Autophagy. Life Sci. Alliance 2022, 5, e202101160. [Google Scholar] [CrossRef]
- Jorgensen, P.; Edgington, N.P.; Schneider, B.L.; Rupeš, I.; Tyers, M.; Futcher, B. The Size of the Nucleus Increases as Yeast Cells Grow. Mol. Biol. Cell 2007, 18, 3523–3532. [Google Scholar] [CrossRef]
- Deolal, P.; Male, G.; Mishra, K. The Challenge of Staying in Shape: Nuclear Size Matters. Curr. Genet. 2021, 67, 605–612. [Google Scholar] [CrossRef]
- Neumann, F.R.; Nurse, P. Nuclear Size Control in Fission Yeast. J. Cell Biol. 2007, 179, 593–600. [Google Scholar] [CrossRef]
- Gregory, T. Genome Size Evolution in Animals. Evol. Genome 2005, 1, 4–87. [Google Scholar]
- Walters, A.D.; Amoateng, K.; Wang, R.; Chen, J.-H.; McDermott, G.; Larabell, C.A.; Gadal, O.; Cohen-Fix, O. Nuclear Envelope Expansion in Budding Yeast Is Independent of Cell Growth and Does Not Determine Nuclear Volume. Mol. Biol. Cell 2019, 30, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Walters, A.D.; Bommakanti, A.; Cohen-Fix, O. Shaping the Nucleus: Factors and Forces. J. Cell. Biochem. 2012, 113, 2813–2821. [Google Scholar] [CrossRef] [PubMed]
- Webster, M.T.; McCaffery, J.M.; Cohen-Fix, O. Vesicle Trafficking Maintains Nuclear Shape in Saccharomyces cerevisiae during Membrane Proliferation. J. Cell Biol. 2010, 191, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Kume, K.; Cantwell, H.; Neumann, F.R.; Jones, A.W.; Snijders, A.P.; Nurse, P. A Systematic Genomic Screen Implicates Nucleocytoplasmic Transport and Membrane Growth in Nuclear Size Control. PLOS Genet. 2017, 13, e1006767. [Google Scholar] [CrossRef] [PubMed]
- Siniossoglou, S.; Santos-Rosa, H.; Rappsilber, J.; Mann, M.; Hurt, E. A Novel Complex of Membrane Proteins Required for Formation of a Spherical Nucleus. EMBO J. 1998, 17, 6449–6464. [Google Scholar] [CrossRef]
- Santos-Rosa, H.; Leung, J.; Grimsey, N.; Peak-Chew, S.; Siniossoglou, S. The Yeast Lipin Smp2 Couples Phospholipid Biosynthesis to Nuclear Membrane Growth. EMBO J. 2005, 24, 1931–1941. [Google Scholar] [CrossRef]
- Han, G.S.; O’Hara, L.; Carman, G.M.; Siniossoglou, S. An Unconventional Diacylglycerol Kinase That Regulates Phospholipid Synthesis and Nuclear Membrane Growth. J. Biol. Chem. 2008, 283, 20433–20442. [Google Scholar] [CrossRef]
- O’Hara, L.; Han, G.S.; Sew, P.C.; Grimsey, N.; Carman, G.M.; Siniossoglou, S. Control of Phospholipid Synthesis by Phosphorylation of the Yeast Lipin Pah1p/Smp2p Mg2+-Dependent Phosphatidate Phosphatase. J. Biol. Chem. 2006, 281, 34537–34548. [Google Scholar] [CrossRef]
- Friederichs, J.M.; Ghosh, S.; Smoyer, C.J.; McCroskey, S.; Miller, B.D.; Weaver, K.J.; Delventhal, K.M.; Unruh, J.; Slaughter, B.D.; Jaspersen, S.L. The SUN Protein Mps3 Is Required for Spindle Pole Body Insertion into the Nuclear Membrane and Nuclear Envelope Homeostasis. PLoS Genet. 2011, 7, e1002365. [Google Scholar] [CrossRef]
- Wright, R.; Basson, M.; D’Ari, L.; Rine, J. Increased Amounts of HMG-CoA Reductase Induce “Karmellae”: A Proliferation of Stacked Membrane Pairs Surrounding the Yeast Nucleus. J. Cell Biol. 1988, 107, 101. [Google Scholar] [CrossRef]
- Siniossoglou, S. Lipins, Lipids and Nuclear Envelope Structure. Traffic 2009, 10, 1181–1187. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, A.D.; Sembongi, H.; Su, W.-M.; Abreu, S.; Reggiori, F.; Carman, G.M.; Siniossoglou, S. Lipid Partitioning at the Nuclear Envelope Controls Membrane Biogenesis. Mol. Biol. Cell 2015, 26, 3641–3657. [Google Scholar] [CrossRef] [PubMed]
- Carman, G.M.; Han, G.S. Phosphatidate Phosphatase Regulates Membrane Phospholipid Synthesis via Phosphatidylserine Synthase. Adv. Biol. Regul. 2018, 67, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Han, G.S.; Wu, W.I.; Carman, G.M. The Saccharomyces cerevisiae Lipin Homolog Is a Mg2+-Dependent Phosphatidate Phosphatase Enzyme. J. Biol. Chem. 2006, 281, 9210–9218. [Google Scholar] [CrossRef]
- Choi, H.-S.; Su, W.-M.; Morgan, J.M.; Han, G.-S.; Xu, Z.; Karanasios, E.; Siniossoglou, S.; Carman, G.M. Phosphorylation of Phosphatidate Phosphatase Regulates Its Membrane Association and Physiological Functions in Saccharomyces cerevisiae. J. Biol. Chem. 2011, 286, 1486–1498. [Google Scholar] [CrossRef]
- Choi, H.S.; Su, W.M.; Han, G.S.; Plote, D.; Xu, Z.; Carman, G.M. Pho85p-Pho80p Phosphorylation of Yeast Pah1p Phosphatidate Phosphatase Regulates Its Activity, Location, Abundance, and Function in Lipid Metabolism. J. Biol. Chem. 2012, 287, 11290–11301. [Google Scholar] [CrossRef]
- Su, W.M.; Han, G.S.; Casciano, J.; Carman, G.M. Protein Kinase A-Mediated Phosphorylation of Pah1p Phosphatidate Phosphatase Functions in Conjunction with the Pho85p-Pho80p and Cdc28p-Cyclin B Kinases to Regulate Lipid Synthesis in Yeast. J. Biol. Chem. 2012, 287, 33364–33376. [Google Scholar] [CrossRef]
- Huffman, T.A.; Mothe-Satney, I.; Lawrence, J.C. Insulin-Stimulated Phosphorylation of Lipin Mediated by the Mammalian Target of Rapamycin. Proc. Natl. Acad. Sci. USA 2002, 99, 1047–1052. [Google Scholar] [CrossRef]
- Grimsey, N.; Han, G.S.; O’Hara, L.; Rochford, J.J.; Carman, G.M.; Siniossoglou, S. Temporal and Spatial Regulation of the Phosphatidate Phosphatases Lipin 1 and 2. J. Biol. Chem. 2008, 283, 29166–29174. [Google Scholar] [CrossRef]
- Harris, T.E.; Huffman, T.A.; Chi, A.; Shabanowitz, J.; Hunt, D.F.; Kumar, A.; Lawrence, J.C. Insulin Controls Subcellular Localization and Multisite Phosphorylation of the Phosphatidic Acid Phosphatase, Lipin 1. J. Biol. Chem. 2007, 282, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Karanasios, E.; Han, G.-S.; Xu, Z.; Carman, G.M.; Siniossoglou, S. A Phosphorylation-Regulated Amphipathic Helix Controls the Membrane Translocation and Function of the Yeast Phosphatidate Phosphatase. Proc. Natl. Acad. Sci. USA 2010, 107, 17539–17544. [Google Scholar] [CrossRef] [PubMed]
- Dubots, E.; Cottier, S.; Péli-Gulli, M.-P.; Jaquenoud, M.; Bontron, S.; Schneiter, R.; De Virgilio, C. TORC1 Regulates Pah1 Phosphatidate Phosphatase Activity via the Nem1/Spo7 Protein Phosphatase Complex. PLoS ONE 2014, 9, e104194. [Google Scholar] [CrossRef]
- Rahman, M.A.; Mostofa, M.G.; Ushimaru, T. The Nem1/Spo7–Pah1/Lipin Axis Is Required for Autophagy Induction after TORC 1 Inactivation. FEBS J. 2018, 285, 1840–1860. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Okamoto, K. The Nem1-Spo7 Protein Phosphatase Complex Is Required for Efficient Mitophagy in Yeast. Biochem. Biophys. Res. Commun. 2018, 496, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Pascual, F.; Carman, G.M. Phosphatidate Phosphatase, a Key Regulator of Lipid Homeostasis. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2013, 1831, 514–522. [Google Scholar] [CrossRef]
- Carman, G.M.; Han, G.S. Fat-Regulating Phosphatidic Acid Phosphatase: A Review of Its Roles and Regulation in Lipid Homeostasis. J. Lipid Res. 2019, 60, 2–6. [Google Scholar] [CrossRef]
- Arnone, J.T.; Walters, A.D.; Cohen-Fix, O. The Dynamic Nature of the Nuclear Envelope. Nucleus 2013, 4, 261–266. [Google Scholar] [CrossRef]
- Romanauska, A.; Köhler, A. The Inner Nuclear Membrane Is a Metabolically Active Territory That Generates Nuclear Lipid Droplets. Cell 2018, 174, 700–715.e18. [Google Scholar] [CrossRef]
- Barbosa, A.D.; Siniossoglou, S. New Kid on the Block: Lipid Droplets in the Nucleus. FEBS J. 2020, 287, 4838–4843. [Google Scholar] [CrossRef]
- Hariri, H.; Rogers, S.; Ugrankar, R.; Liu, Y.L.; Feathers, J.R.; Henne, W.M. Lipid Droplet Biogenesis Is Spatially Coordinated at ER–Vacuole Contacts under Nutritional Stress. EMBO Rep. 2018, 19, 57–72. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Roberts, P.; Chen, Y.; Kvam, E.; Shulga, N.; Huang, K.; Lemmon, S.; Goldfarb, D.S. Nucleus–Vacuole Junctions in Saccharomyces cerevisiae Are Formed Through the Direct Interaction of Vac8p with Nvj1p. Mol. Biol. Cell 2000, 11, 2445. [Google Scholar] [CrossRef] [PubMed]
- Tomioka, Y.; Kotani, T.; Kirisako, H.; Oikawa, Y.; Kimura, Y.; Hirano, H.; Ohsumi, Y.; Nakatogawa, H. TORC1 Inactivation Stimulates Autophagy of Nucleoporin and Nuclear Pore Complexes. J. Cell Biol. 2020, 219, e201910063. [Google Scholar] [CrossRef] [PubMed]
- Scorrano, L.; De Matteis, M.A.; Emr, S.; Giordano, F.; Hajnóczky, G.; Kornmann, B.; Lackner, L.L.; Levine, T.P.; Pellegrini, L.; Reinisch, K.; et al. Coming Together to Define Membrane Contact Sites. Nat. Commun. 2019, 10, 1287. [Google Scholar] [CrossRef]
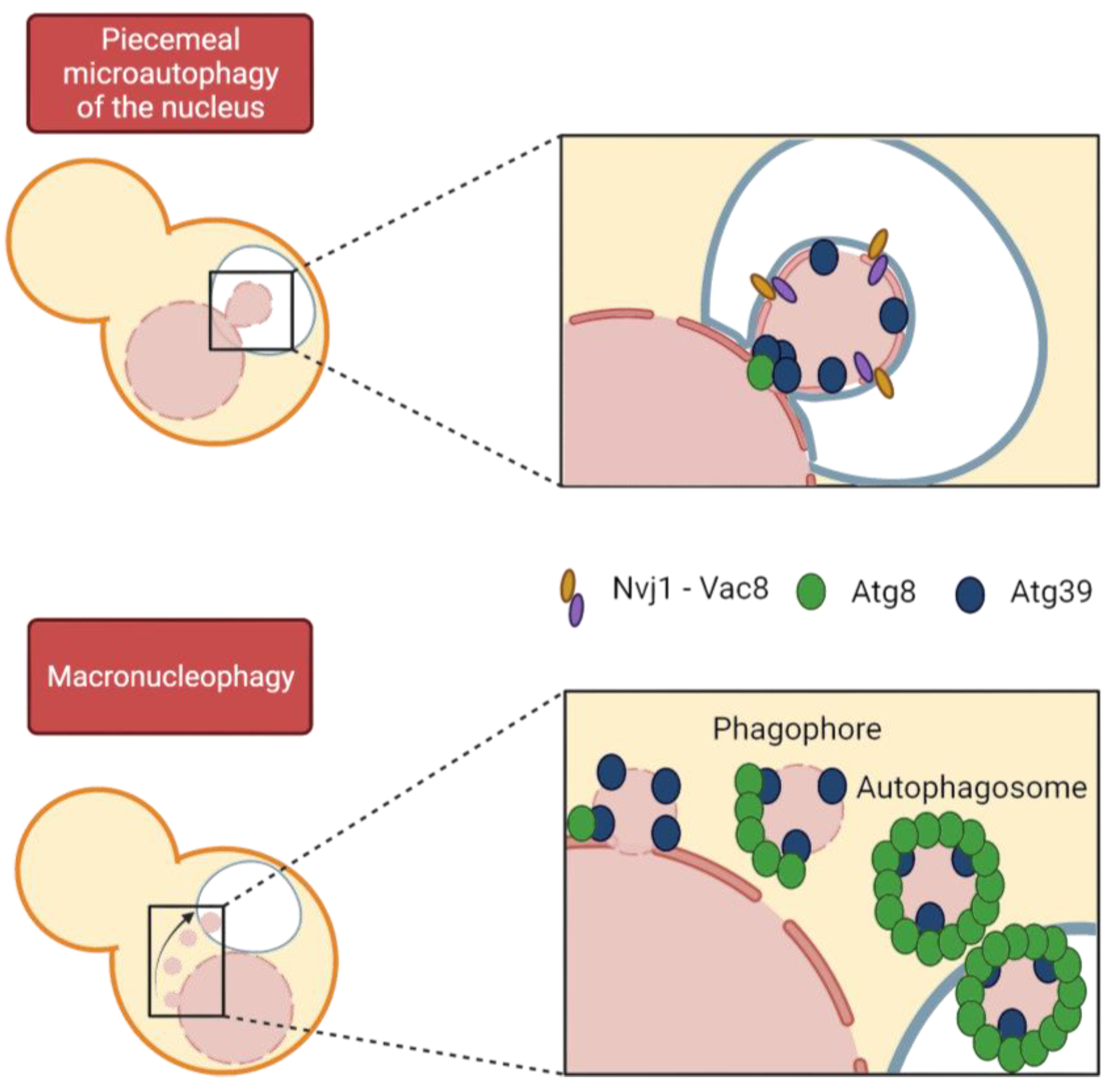
- Kvam, E.; Goldfarb, D.S. Nucleus-Vacuole Junctions and Piecemeal Microautophagy of the Nucleus in S. cerevisiae. Autophagy 2007, 3, 85–92. [Google Scholar] [CrossRef]
- Banta, L.M.; Robinson, J.S.; Klionsky, D.J.; Emr, S.D. Organelle Assembly in Yeast: Characterization of Yeast Mutants Defective in Vacuolar Biogenesis and Protein Sorting. J. Cell Biol. 1988, 107, 1369–1383. [Google Scholar] [CrossRef]
- Jin, Y.; Weisman, L.S. The Vacuole/Lysosome Is Required for Cell-Cycle Progression. Elife 2015, 4, e08160. [Google Scholar] [CrossRef]
- Weisman, L.S. Yeast Vacuole Inheritance and Dynamics. Annu. Rev. Genet. 2003, 37, 435–460. [Google Scholar] [CrossRef]
- Baba, M.; Takeshige, K.; Baba, N.; Ohsumi, Y. Ultrastructural Analysis of the Autophagic Process in Yeast: Detection of Autophagosomes and Their Characterization. J. Cell Biol. 1994, 124, 903–913. [Google Scholar] [CrossRef]
- Sasser, T.; Qiu, Q.S.; Karunakaran, S.; Padolina, M.; Reyes, A.; Flood, B.; Smith, S.; Gonzales, C.; Fratti, R.A. Yeast Lipin 1 Orthologue Pah1p Regulates Vacuole Homeostasis and Membrane Fusion. J. Biol. Chem. 2012, 287, 2221–2236. [Google Scholar] [CrossRef]
- Michaillat, L.; Mayer, A. Identification of Genes Affecting Vacuole Membrane Fragmentation in Saccharomyces cerevisiae. PLoS ONE 2013, 8, e54160. [Google Scholar] [CrossRef]
- Papandreou, M.E.; Tavernarakis, N. Nucleophagy Mediators and Mechanisms. Prog. Mol. Biol. Transl. Sci. 2020, 172, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Madura, K. Degradation of Specific Nuclear Proteins Occurs in the Cytoplasm in Saccharomyces cerevisiae. Genetics 2014, 197, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Roberts, P.; Moshitch-Moshkovitz, S.; Kvam, E.; O’Toole, E.; Winey, M.; Goldfarb, D.S. Piecemeal Microautophagy of Nucleus in Saccharomyces Cerevisiae. Mol. Biol. Cell 2003, 14, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Krick, R.; Mühe, Y.; Prick, T.; Bredschneider, M.; Bremer, S.; Wenzel, D.; Eskelinen, E.L.; Thumm, M. Piecemeal Microautophagy of the Nucleus: Genetic and Morphological Traits. Autophagy 2009, 5, 270–272. [Google Scholar] [CrossRef]
- Mijaljica, D.; Prescott, M.; Devenish, R.J. A Late Form of Nucleophagy in Saccharomyces cerevisiae. PLoS ONE 2012, 7, e40013. [Google Scholar] [CrossRef]
- Mochida, K.; Oikawa, Y.; Kimura, Y.; Kirisako, H.; Hirano, H.; Ohsumi, Y.; Nakatogawa, H. Receptor-Mediated Selective Autophagy Degrades the Endoplasmic Reticulum and the Nucleus. Nature 2015, 522, 359–362. [Google Scholar] [CrossRef]
- Otto, F.B.; Thumm, M. Mechanistic Dissection of Macro- and Micronucleophagy. Autophagy 2021, 17, 626–639. [Google Scholar] [CrossRef]
- Tosal-Castano, S.; Peselj, C.; Kohler, V.; Habernig, L.; Berglund, L.L.; Ebrahimi, M.; Vögtle, F.N.; Höög, J.; Andréasson, C.; Büttner, S. Snd3 Controls Nucleus-Vacuole Junctions in Response to Glucose Signaling. Cell Rep. 2021, 34, 108637. [Google Scholar] [CrossRef]
- Dawaliby, R.; Mayer, A. Microautophagy of the Nucleus Coincides with a Vacuolar Diffusion Barrier at Nuclear-Vacuolar Junctions. Mol. Biol. Cell 2010, 21, 4173–4183. [Google Scholar] [CrossRef] [PubMed]
- Kvam, E.; Gable, K.; Dunn, T.M.; Goldfarb, D.S. Targeting of Tsc13p to Nucleus-Vacuole Junctions: A Role for Very-Long-Chain Fatty Acids in the Biogenesis of Microautophagic Vesicles. Mol. Biol. Cell 2005, 16, 3987–3998. [Google Scholar] [CrossRef]
- Henne, W.M.; Zhu, L.; Balogi, Z.; Stefan, C.; Pleiss, J.A.; Emr, S.D. Mdm1/Snx13 Is a Novel ER–Endolysosomal Interorganelle Tethering Protein. J. Cell Biol. 2015, 210, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Otto, F.B.; Thumm, M. Nucleophagy—Implications for Microautophagy and Health. Int. J. Mol. Sci. 2020, 21, 4506. [Google Scholar] [CrossRef] [PubMed]
- Millen, J.I.; Krick, R.; Prick, T.; Thumm, M.; Goldfarb, D.S. Measuring Piecemeal Microautophagy of the Nucleus in Saccharomyces cerevisiae. Autophagy 2009, 5, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Wilfling, F.; Ronchi, P.; Allegretti, M.; Mosalaganti, S.; Jentsch, S.; Beck, M.; Pfander, B. Selective Autophagy Degrades Nuclear Pore Complexes. Nat. Cell Biol. 2020, 22, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Mostofa, M.G.; Rahman, M.A.; Koike, N.; Yeasmin, A.M.S.T.; Islam, N.; Waliullah, T.M.; Hosoyamada, S.; Shimobayashi, M.; Kobayashi, T.; Hall, M.N.; et al. CLIP and Cohibin Separate RDNA from Nucleolar Proteins Destined for Degradation by Nucleophagy. J. Cell Biol. 2018, 217, 2675–2690. [Google Scholar] [CrossRef] [PubMed]
- Mostofa, M.G.; Morshed, S.; Shibata, R.; Takeichi, Y.; Rahman, M.A.; Hosoyamada, S.; Kobayashi, T.; Ushimaru, T. RDNA Condensation Promotes RDNA Separation from Nucleolar Proteins Degraded for Nucleophagy after TORC1 Inactivation. Cell Rep. 2019, 28, 3423–3434.e2. [Google Scholar] [CrossRef]
- Reggiori, F.; Klionsky, D.J. Autophagic Processes in Yeast: Mechanism, Machinery and Regulation. Genetics 2013, 194, 341–361. [Google Scholar] [CrossRef]
- Mijaljica, D.; Klionsky, D.J. The Necessity of Nucleophagic Modality. Autophagy 2022, 18, 443. [Google Scholar] [CrossRef]
- Papandreou, M.E.; Tavernarakis, N. Nucleophagy: From Homeostasis to Disease. Cell Death Differ. 2019, 26, 630–639. [Google Scholar] [CrossRef]
- Loewith, R.; Hall, M.N. Target of Rapamycin (TOR) in Nutrient Signaling and Growth Control. Genetics 2011, 189, 1177–1201. [Google Scholar] [CrossRef] [PubMed]
- Noda, T.; Ohsumi, Y. Tor, a Phosphatidylinositol Kinase Homologue, Controls Autophagy in Yeast. J. Biol. Chem. 1998, 273, 3963–3966. [Google Scholar] [CrossRef] [PubMed]
- Noda, T. Regulation of Autophagy through TORC1 and MTORC1. Biomolecules 2017, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Terasawa, M.; Mostofa, M.G.; Ushimaru, T. The TORC1–Nem1/Spo7–Pah1/Lipin Axis Regulates Microautophagy Induction in Budding Yeast. Biochem. Biophys. Res. Commun. 2018, 504, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Ueda, S.; Ozaki, R.; Kaneko, A.; Akizuki, R.; Katsuta, H.; Miura, A.; Matsuura, A.; Ushimaru, T. TORC1, Tel1/Mec1, and Mpk1 Regulate Autophagy Induction after DNA Damage in Budding Yeast. Cell. Signal. 2019, 62, 109344. [Google Scholar] [CrossRef] [PubMed]
- Morshed, S.; Sharmin, T.; Ushimaru, T. TORC1 Regulates ESCRT-0 Complex Formation on the Vacuolar Membrane and Microautophagy Induction in Yeast. Biochem. Biophys. Res. Commun. 2020, 522, 88–94. [Google Scholar] [CrossRef]
- Kvam, E.; Goldfarb, D.S. Structure and Function of Nucleus-Vacuole Junctions: Outer-Nuclear-Membrane Targeting of Nvj1p and a Role in Tryptophan Uptake. J. Cell Sci. 2006, 119, 3622–3633. [Google Scholar] [CrossRef]
- Ha, C.W.; Huh, W.K. Rapamycin Increases RDNA Stability by Enhancing Association of Sir2 with RDNA in Saccharomyces cerevisiae. Nucleic Acids Res. 2011, 39, 1336–1350. [Google Scholar] [CrossRef]
- Inokoshi, J.; Tomoda, H.; Hashimoto, H.; Watanabe, A.; Takeshima, H.; Ōmura, S. Cerulenin-Resistant Mutants of Saccharomyces cerevisiae with an Altered Fatty Acid Synthase Gene. Mol. Gen. Genet. MGG 1994, 244, 90–96. [Google Scholar] [CrossRef]
- Nakatogawa, H.; Mochida, K. Reticulophagy and Nucleophagy: New Findings and Unsolved Issues. Autophagy 2015, 11, 2377–2378. [Google Scholar] [CrossRef]
- Krick, R.; Muehe, Y.; Prick, T.; Bremer, S.; Schlotterhose, P.; Eskelinen, E.L.; Millen, J.; Goldfarb, D.S.; Thumm, M. Piecemeal Microautophagy of the Nucleus Requires the Core Macroautophagy Genes. Mol. Biol. Cell 2008, 19, 4492–4505. [Google Scholar] [CrossRef] [PubMed]
- Shpilka, T.; Welter, E.; Borovsky, N.; Amar, N.; Mari, M.; Reggiori, F.; Elazar, Z. Lipid Droplets and Their Component Triglycerides and Steryl Esters Regulate Autophagosome Biogenesis. EMBO J. 2015, 34, 2117–2131. [Google Scholar] [CrossRef]
- Matos-Perdomo, E.; Santana-Sosa, S.; Ayra-Plasencia, J.; Medina-Suárez, S.; Machín, F. The Vacuole Shapes the Nucleus and the Ribosomal DNA Loop during Mitotic Delays. Life Sci. Alliance 2022, 5, e202101161. [Google Scholar] [CrossRef] [PubMed]
- Nakatogawa, H.; Suzuki, K.; Kamada, Y.; Ohsumi, Y. Dynamics and Diversity in Autophagy Mechanisms: Lessons from Yeast. Nat. Rev. Mol. Cell Biol. 2009, 10, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.L.; Cuervo, A.M. Autophagy and Human Disease: Emerging Themes. Curr. Opin. Genet. Dev. 2014, 26, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Papandreou, M.-E.; Konstantinidis, G.; Tavernarakis, N. Nucleophagy Delays Aging and Preserves Germline Immortality. Nat. Aging 2023, 3, 34–46. [Google Scholar] [CrossRef]
- Lazar-Stefanita, L.; Scolari, V.F.; Mercy, G.; Muller, H.; Guérin, T.M.; Thierry, A.; Mozziconacci, J.; Koszul, R. Cohesins and Condensins Orchestrate the 4D Dynamics of Yeast Chromosomes during the Cell Cycle. EMBO J. 2017, 36, 2684–2697. [Google Scholar] [CrossRef]
- Almacellas, E.; Pelletier, J.; Day, C.; Ambrosio, S.; Tauler, A.; Mauvezin, C. Lysosomal Degradation Ensures Accurate Chromosomal Segregation to Prevent Chromosomal Instability. Autophagy 2021, 17, 796–813. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santana-Sosa, S.; Matos-Perdomo, E.; Ayra-Plasencia, J.; Machín, F. A Yeast Mitotic Tale for the Nucleus and the Vacuoles to Embrace. Int. J. Mol. Sci. 2023, 24, 9829. https://doi.org/10.3390/ijms24129829
Santana-Sosa S, Matos-Perdomo E, Ayra-Plasencia J, Machín F. A Yeast Mitotic Tale for the Nucleus and the Vacuoles to Embrace. International Journal of Molecular Sciences. 2023; 24(12):9829. https://doi.org/10.3390/ijms24129829
Chicago/Turabian StyleSantana-Sosa, Silvia, Emiliano Matos-Perdomo, Jessel Ayra-Plasencia, and Félix Machín. 2023. "A Yeast Mitotic Tale for the Nucleus and the Vacuoles to Embrace" International Journal of Molecular Sciences 24, no. 12: 9829. https://doi.org/10.3390/ijms24129829
APA StyleSantana-Sosa, S., Matos-Perdomo, E., Ayra-Plasencia, J., & Machín, F. (2023). A Yeast Mitotic Tale for the Nucleus and the Vacuoles to Embrace. International Journal of Molecular Sciences, 24(12), 9829. https://doi.org/10.3390/ijms24129829