
Imaging Approaches to Investigate Pathophysiological Mechanisms of Brain Disease in Zebrafish

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
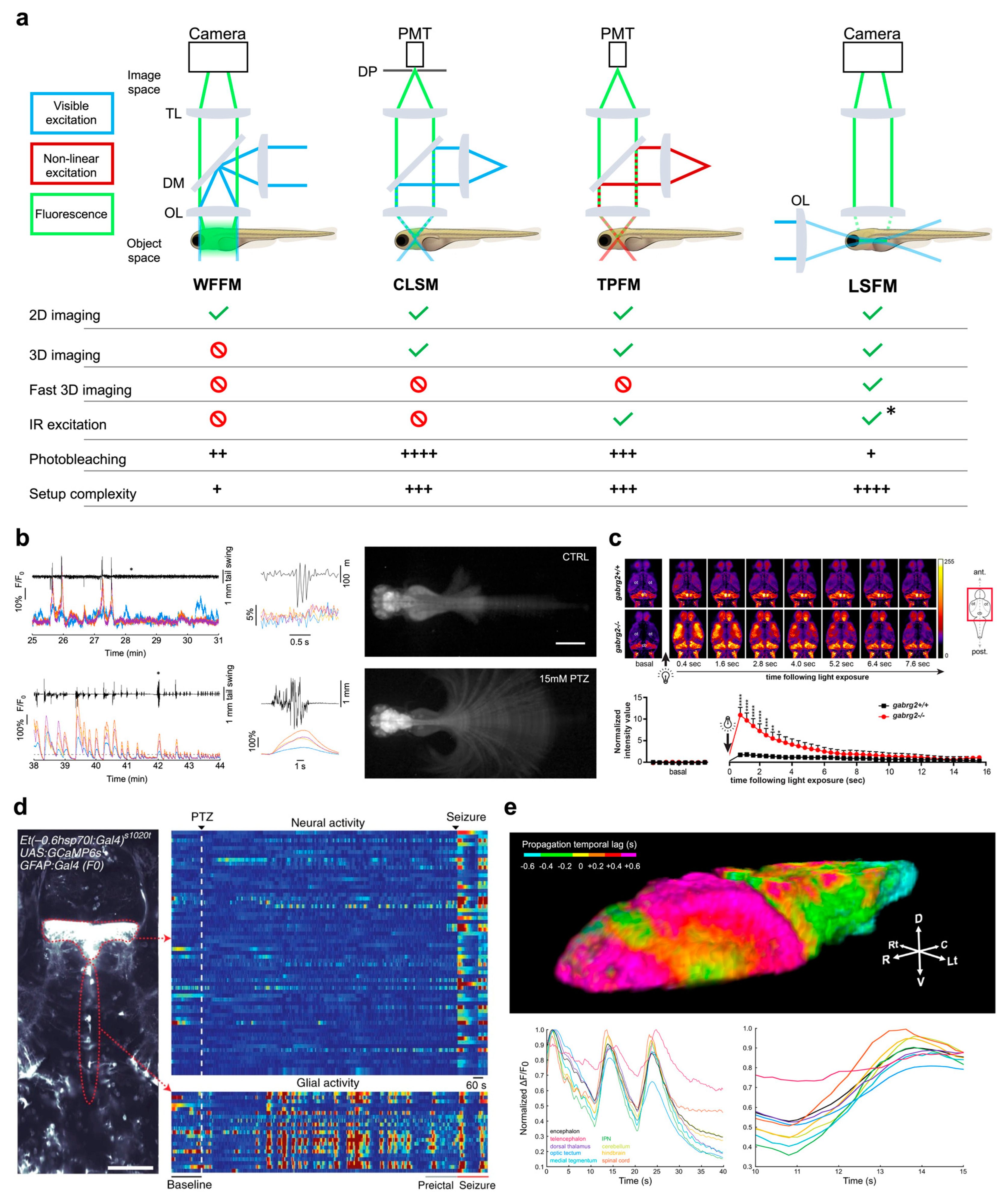
2. Functional Imaging

3. Structural Imaging

4. Behavioral Imaging

5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feigin, V.L.; Vos, T.; Nichols, E.; Owolabi, M.O.; Carroll, W.M.; Dichgans, M.; Deuschl, G.; Parmar, P.; Brainin, M.; Murray, C. The Global Burden of Neurological Disorders: Translating Evidence into Policy. Lancet Neurol. 2020, 19, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Nobrega, M.A.; Pennacchio, L.A. Comparative Genomic Analysis as a Tool for Biological Discovery. J. Physiol. 2004, 554 Pt 1, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Folgueira, M.; Bayley, P.; Navratilova, P.; Becker, T.S.; Wilson, S.W.; Clarke, J.D. Morphogenesis Underlying the Development of the Everted Teleost Telencephalon. Neural Dev. 2012, 7, 32. [Google Scholar] [CrossRef]
- Kawai, H.; Arata, N.; Nakayasu, H. Three-Dimensional Distribution of Astrocytes in Zebrafish Spinal Cord. Glia 2001, 36, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M.; McMurray, J.V.; Eisen, J.S. Identified Motoneurons and Their Innervation of Axial Muscles in the Zebrafish. J. Neurosci. Off. J. Soc. Neurosci. 1986, 6, 2267–2277. [Google Scholar] [CrossRef] [PubMed]
- Koulen, P.; Janowitz, T.; Johnston, L.D.; Ehrlich, B.E. Conservation of Localization Patterns of Ip(3) Receptor Type 1 in Cerebellar Purkinje Cells across Vertebrate Species. J. Neurosci. Res. 2000, 61, 493–499. [Google Scholar] [CrossRef]
- Avila, R.L.; Tevlin, B.R.; Lees, J.P.; Inouye, H.; Kirschner, D.A. Myelin Structure and Composition in Zebrafish. Neurochem. Res. 2007, 32, 197–209. [Google Scholar] [CrossRef]
- Rico, E.P.; Rosemberg, D.B.; Seibt, K.J.; Capiotti, K.M.; Da Silva, R.S.; Bonan, C.D. Zebrafish Neurotransmitter Systems as Potential Pharmacological and Toxicological Targets. Neurotoxicol. Teratol. 2011, 33, 608–617. [Google Scholar] [CrossRef]
- Renier, C.; Faraco, J.H.; Bourgin, P.; Motley, T.; Bonaventure, P.; Rosa, F.; Mignot, E. Genomic and Functional Conservation of Sedative-Hypnotic Targets in the Zebrafish. Pharm. Genom. 2007, 17, 237–253. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, J.B.; He, K.J.; Wang, F.; Liu, C.F. Advances of Zebrafish in Neurodegenerative Disease: From Models to Drug Discovery. Front. Pharmacol. 2021, 12, 713963. [Google Scholar] [CrossRef]
- Baraban, S.C.; Taylor, M.R.; Castro, P.A.; Baier, H. Pentylenetetrazole Induced Changes in Zebrafish Behavior, Neural Activity and C-Fos Expression. Neuroscience 2005, 131, 759–768. [Google Scholar] [CrossRef]
- Lam, C.S.; Korzh, V.; Strahle, U. Zebrafish Embryos Are Susceptible to the Dopaminergic Neurotoxin Mptp. Eur. J. Neurosci. 2005, 21, 1758–1762. [Google Scholar] [CrossRef] [PubMed]
- Thawkar, B.S.; Kaur, G. Zebrafish as a Promising Tool for Modeling Neurotoxin-Induced Alzheimer’s Disease. Neurotox. Res. 2021, 39, 949–965. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The Zebrafish Reference Genome Sequence and Its Relationship to the Human Genome. Nature 2013, 496, 498–503. [Google Scholar]
- Baraban, S.C.; Dinday, M.T.; Hortopan, G.A. Drug Screening in Scn1a Zebrafish Mutant Identifies Clemizole as a Potential Dravet Syndrome Treatment. Nat. Commun. 2013, 4, 2410. [Google Scholar] [CrossRef] [PubMed]
- Samarut, E.; Swaminathan, A.; Riche, R.; Liao, M.; Hassan-Abdi, R.; Renault, S.; Allard, M.; Dufour, L.; Cossette, P.; Soussi-Yanicostas, N.; et al. Gamma-Aminobutyric Acid Receptor Alpha 1 Subunit Loss of Function Causes Genetic Generalized Epilepsy by Impairing Inhibitory Network Neurodevelopment. Epilepsia 2018, 59, 2061–2074. [Google Scholar] [CrossRef] [PubMed]
- Yaksi, E.; Jamali, A.; Verdugo, C.D.; Jurisch-Yaksi, N. Past, Present and Future of Zebrafish in Epilepsy Research. FEBS J. 2021, 288, 7243–7255. [Google Scholar] [CrossRef] [PubMed]
- D’Amora, M.; Galgani, A.; Marchese, M.; Tantussi, F.; Faraguna, U.; De Angelis, F.; Giorgi, F.S. Zebrafish as an Innovative Tool for Epilepsy Modeling: State of the Art and Potential Future Directions. Int. J. Mol. Sci. 2023, 24, 7702. [Google Scholar] [CrossRef]
- Newman, M.; Ebrahimie, E.; Lardelli, M. Using the Zebrafish Model for Alzheimer’s Disease Research. Front. Genet. 2014, 5, 189. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, E.J.; Turner, K.J.; Fernandez, J.M.; Cifuentes, D.; Ghosh, M.; Ijaz, S.; Jain, R.A.; Kubo, F.; Bill, B.R.; Baier, H.; et al. Estrogens Suppress a Behavioral Phenotype in Zebrafish Mutants of the Autism Risk Gene, Cntnap2. Neuron 2016, 89, 725–733. [Google Scholar] [CrossRef]
- Liu, C.X.; Li, C.Y.; Hu, C.C.; Wang, Y.; Lin, J.; Jiang, Y.H.; Li, Q.; Xu, X. Crispr/Cas9-Induced Shank3b Mutant Zebrafish Display Autism-Like Behaviors. Mol. Autism 2018, 9, 23. [Google Scholar] [CrossRef] [PubMed]
- Burgess, H.A.; Granato, M. Modulation of Locomotor Activity in Larval Zebrafish during Light Adaptation. J. Exp. Biol. 2007, 210 Pt 14, 2526–2539. [Google Scholar] [CrossRef] [PubMed]
- Hein, A.M.; Gil, M.A.; Twomey, C.R.; Couzin, I.D.; Levin, S.A. Conserved Behavioral Circuits Govern High-Speed Decision-Making in Wild Fish Shoals. Proc. Natl. Acad. Sci. USA 2018, 115, 12224–12228. [Google Scholar] [CrossRef]
- Gerlai, R. Evolutionary Conservation, Translational Relevance and Cognitive Function: The Future of Zebrafish in Behavioral Neuroscience. Neurosci. Biobehav. Rev. 2020, 116, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.Z.; Schnitzer, M.J. Genetically Encoded Indicators of Neuronal Activity. Nat. Neurosci. 2016, 19, 1142–1153. [Google Scholar] [CrossRef]
- Conti, E.; Allegra Mascaro, A.L.; Pavone, F.S. Large Scale Double-Path Illumination System with Split Field of View for the All-Optical Study of Inter-and Intra-Hemispheric Functional Connectivity on Mice. Methods Protoc. 2019, 2, 11. [Google Scholar] [CrossRef]
- Conti, E.; Scaglione, A.; de Vito, G.; Calugi, F.; Pasquini, M.; Pizzorusso, T.; Micera, S.; Mascaro, A.L.A.; Pavone, F.S. Combining Optogenetic Stimulation and Motor Training Improves Functional Recovery and Perilesional Cortical Activity. Neurorehabilit. Neural Repair. 2022, 36, 107–118. [Google Scholar] [CrossRef]
- Turrini, L.; Fornetto, C.; Marchetto, G.; Mullenbroich, M.C.; Tiso, N.; Vettori, A.; Resta, F.; Masi, A.; Mannaioni, G.; Pavone, F.S.; et al. Optical Mapping of Neuronal Activity during Seizures in Zebrafish. Sci. Rep. 2017, 7, 3025. [Google Scholar] [CrossRef]
- Zipfel, W.R.; Williams, R.M.; Webb, W.W. Nonlinear Magic: Multiphoton Microscopy in the Biosciences. Nat. Biotechnol. 2003, 21, 1369–1377. [Google Scholar] [CrossRef]
- Power, R.M.; Huisken, J. A Guide to Light-Sheet Fluorescence Microscopy for Multiscale Imaging. Nat. Methods 2017, 14, 360–373. [Google Scholar] [CrossRef]
- Mullenbroich, M.C.; Turrini, L.; Silvestri, L.; Alterini, T.; Gheisari, A.; Tiso, N.; Vanzi, F.; Sacconi, L.; Pavone, F.S. Bessel Beam Illumination Reduces Random and Systematic Errors in Quantitative Functional Studies Using Light-Sheet Microscopy. Front. Cell. Neurosci. 2018, 12, 315. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; Lauderdale, J.D.; Sornborger, A.T. Mapping Functional Connectivity between Neuronal Ensembles with Larval Zebrafish Transgenic for a Ratiometric Calcium Indicator. Front. Neural Circuits 2011, 5, 2. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Baraban, S.C. Network Properties Revealed During Multi-Scale Calcium Imaging of Seizure Activity in Zebrafish. eNeuro 2019, 6, ENEURO.0041-19.2019. [Google Scholar] [CrossRef] [PubMed]
- Kanyo, R.; Wang, C.K.; Locskai, L.F.; Li, J.; Allison, W.T.; Kurata, H.T. Functional and Behavioral Signatures of Kv7 Activator Drug Subtypes. Epilepsia 2020, 61, 1678–1690. [Google Scholar] [CrossRef]
- Brenet, A.; Hassan-Abdi, R.; Somkhit, J.; Yanicostas, C.; Soussi-Yanicostas, N. Defective Excitatory/Inhibitory Synaptic Balance and Increased Neuron Apoptosis in a Zebrafish Model of Dravet Syndrome. Cells 2019, 8, 1199. [Google Scholar] [CrossRef]
- Liao, M.; Kundap, U.; Rosch, R.E.; Burrows, D.R.W.; Meyer, M.P.; Bencheikh, B.O.A.; Cossette, P.; Samarut, E. Targeted Knockout of Gaba-a Receptor Gamma 2 Subunit Provokes Transient Light-Induced Reflex Seizures in Zebrafish Larvae. Dis. Model Mech. 2019, 12, dmm040782. [Google Scholar] [CrossRef]
- Liu, J.; Salvati, K.A.; Baraban, S.C. In Vivo Calcium Imaging Reveals Disordered Interictal Network Dynamics in Epileptic Stxbp1b Zebrafish. iScience 2021, 24, 102558. [Google Scholar] [CrossRef]
- Jacobs, G.H. The Evolution of Vertebrate Color Vision. Adv. Exp. Med. Biol. 2012, 739, 156–172. [Google Scholar]
- Diaz Verdugo, C.; Myren-Svelstad, S.; Aydin, E.; Van Hoeymissen, E.; Deneubourg, C.; Vanderhaeghe, S.; Vancraeynest, J.; Pelgrims, R.; Cosacak, M.I.; Muto, A.; et al. Glia-Neuron Interactions Underlie State Transitions to Generalized Seizures. Nat. Commun. 2019, 10, 3830. [Google Scholar] [CrossRef]
- Cozzolino, O.; Sicca, F.; Paoli, E.; Trovato, F.; Santorelli, F.M.; Ratto, G.M.; Marchese, M. Evolution of Epileptiform Activity in Zebrafish by Statistical-Based Integration of Electrophysiology and 2-Photon Ca(2+) Imaging. Cells 2020, 9, 769. [Google Scholar] [CrossRef]
- Niemeyer, J.E.; Gadamsetty, P.; Chun, C.; Sylvester, S.; Lucas, J.P.; Ma, H.; Schwartz, T.H.; Aksay, E.R.F. Seizures Initiate in Zones of Relative Hyperexcitation in a Zebrafish Epilepsy Model. Brain 2022, 145, 2347–2360. [Google Scholar] [CrossRef] [PubMed]
- Hadjiabadi, D.; Lovett-Barron, M.; Raikov, I.G.; Sparks, F.T.; Liao, Z.; Baraban, S.C.; Leskovec, J.; Losonczy, A.; Deisseroth, K.; Soltesz, I. Maximally Selective Single-Cell Target for Circuit Control in Epilepsy Models. Neuron 2021, 109, 2556–2572.e6. [Google Scholar] [CrossRef] [PubMed]
- Andalman, A.S.; Burns, V.M.; Lovett-Barron, M.; Broxton, M.; Poole, B.; Yang, S.J.; Grosenick, L.; Lerner, T.N.; Chen, R.; Benster, T.; et al. Neuronal Dynamics Regulating Brain and Behavioral State Transitions. Cell 2019, 1774, 970–985.e20. [Google Scholar] [CrossRef]
- Haney, W.A.; Moussaoui, B.; Strother, J.A. Prolonged Exposure to Stressors Suppresses Exploratory Behavior in Zebrafish Larvae. J. Exp. Biol. 2020, 223 Pt 22, jeb224964. [Google Scholar] [CrossRef]
- Ahrens, M.B.; Orger, M.B.; Robson, D.N.; Li, J.M.; Keller, P.J. Whole-Brain Functional Imaging at Cellular Resolution Using Light-Sheet Microscopy. Nat. Methods 2013, 10, 413–420. [Google Scholar] [CrossRef]
- Rosch, R.E.; Hunter, P.R.; Baldeweg, T.; Friston, K.J.; Meyer, M.P. Calcium Imaging and Dynamic Causal Modelling Reveal Brain-Wide Changes in Effective Connectivity and Synaptic Dynamics During Epileptic Seizures. PLoS Comput. Biol. 2018, 14, e1006375. [Google Scholar] [CrossRef] [PubMed]
- Winter, M.J.; Windell, D.; Metz, J.; Matthews, P.; Pinion, J.; Brown, J.T.; Hetheridge, M.J.; Ball, J.S.; Owen, S.F.; Redfern, W.S.; et al. 4-Dimensional Functional Profiling in the Convulsant-Treated Larval Zebrafish Brain. Sci. Rep. 2017, 7, 6581. [Google Scholar] [CrossRef] [PubMed]
- Winter, M.J.; Pinion, J.; Tochwin, A.; Takesono, A.; Ball, J.S.; Grabowski, P.; Metz, J.; Trznadel, M.; Tse, K.; Redfern, W.S.; et al. Functional Brain Imaging in Larval Zebrafish for Characterising the Effects of Seizurogenic Compounds Acting via a Range of Pharmacological Mechanisms. Br. J. Pharm. 2021, 178, 2671–2689. [Google Scholar] [CrossRef]
- de Vito, G.; Turrini, L.; Mullenbroich, C.; Ricci, P.; Sancataldo, G.; Mazzamuto, G.; Tiso, N.; Sacconi, L.; Fanelli, D.; Silvestri, L.; et al. Fast Whole-Brain Imaging of Seizures in Zebrafish Larvae by Two-Photon Light-Sheet Microscopy. Biomed. Opt. Express 2022, 13, 1516–1536. [Google Scholar] [CrossRef]
- Turrini, L.; Sorelli, M.; de Vito, G.; Credi, C.; Tiso, N.; Vanzi, F.; Pavone, F.S. Multimodal Characterization of Seizures in Zebrafish Larvae. Biomedicines 2022, 10, 951. [Google Scholar] [CrossRef]
- Batista-Garcia-Ramo, K.; Fernandez-Verdecia, C.I. What We Know About the Brain Structure-Function Relationship. Behav. Sci. 2018, 8, 39. [Google Scholar] [CrossRef] [PubMed]
- Oksenberg, N.; Stevison, L.; Wall, J.D.; Ahituv, N. Function and Regulation of Auts2, a Gene Implicated in Autism and Human Evolution. PLoS Genet. 2013, 9, e1003221. [Google Scholar] [CrossRef] [PubMed]
- Bernier, R.; Golzio, C.; Xiong, B.; Stessman, H.A.; Coe, B.P.; Penn, O.; Witherspoon, K.; Gerdts, J.; Baker, C.; Silfhout, A.T.V.-V.; et al. Disruptive Chd8 Mutations Define a Subtype of Autism Early in Development. Cell 2014, 158, 263–276. [Google Scholar] [CrossRef]
- Kozol, R.A.; Cukier, H.N.; Zou, B.; Mayo, V.; De Rubeis, S.; Cai, G.; Griswold, A.J.; Whitehead, P.L.; Haines, J.L.; Gilbert, J.R.; et al. Two Knockdown Models of the Autism Genes Syngap1 and Shank3 in Zebrafish Produce Similar Behavioral Phenotypes Associated with Embryonic Disruptions of Brain Morphogenesis. Hum. Mol. Genet. 2015, 24, 4006–4023. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Long, F.; Cao, X.; Xiong, B.; Li, Y. Knockout of Katnal2 Leads to Autism-Like Behaviors and Developmental Delay in Zebrafish. Int. J. Mol. Sci. 2022, 23, 8389. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.X.; Peng, X.L.; Hu, C.C.; Li, C.Y.; Li, Q.; Xu, X. Developmental Profiling of Asd-Related Shank3 Transcripts and Their Differential Regulation by Valproic Acid in Zebrafish. Dev. Genes Evol. 2016, 226, 389–400. [Google Scholar] [CrossRef]
- Luffe, T.M.; D’Orazio, A.; Bauer, M.; Gioga, Z.; Schoeffler, V.; Lesch, K.P.; Romanos, M.; Drepper, C.; Lillesaar, C. Increased Locomotor Activity Via Regulation of Gabaergic Signalling in Foxp2 Mutant Zebrafish-Implications for Neurodevelopmental Disorders. Transl. Psychiatry 2021, 11, 529. [Google Scholar] [CrossRef]
- Kozol, R.A.; James, D.M.; Varela, I.; Sumathipala, S.H.; Zuchner, S.; Dallman, J.E. Restoring Shank3 in the Rostral Brainstem of Shank3ab-/- Zebrafish Autism Models Rescues Sensory Deficits. Commun. Biol. 2021, 4, 1411. [Google Scholar] [CrossRef]
- Elsen, G.E.; Choi, L.Y.; Prince, V.E.; Ho, R.K. The Autism Susceptibility Gene Met Regulates Zebrafish Cerebellar Development and Facial Motor Neuron Migration. Dev. Biol. 2009, 335, 78–92. [Google Scholar] [CrossRef]
- Miller, A.C.; Voelker, L.H.; Shah, A.N.; Moens, C.B. Neurobeachin Is Required Postsynaptically for Electrical and Chemical Synapse Formation. Curr. Biol. 2015, 25, 16–28. [Google Scholar] [CrossRef]
- Bhattarai, P.; Cosacak, M.I.; Mashkaryan, V.; Demir, S.; Popova, S.D.; Govindarajan, N.; Brandt, K.; Zhang, Y.; Chang, W.; Ampatzis, K.; et al. Neuron-Glia Interaction through Serotonin-Bdnf-Ngfr Axis Enables Regenerative Neurogenesis in Alzheimer’s Model of Adult Zebrafish Brain. PLoS Biol. 2020, 18, e3000585. [Google Scholar] [CrossRef]
- Vaz, R.L.; Sousa, S.; Chapela, D.; van der Linde, H.C.; Willemsen, R.; Correia, A.D.; Outeiro, T.F.; Afonso, N.D. Identification of Antiparkinsonian Drugs in the 6-Hydroxydopamine Zebrafish Model. Pharmacol. Biochem. Behav. 2020, 189, 172828. [Google Scholar] [CrossRef]
- Kim, G.J.; Mo, H.; Liu, H.; Wu, Z.; Chen, S.; Zheng, J.; Zhao, X.; Nucum, D.; Shortland, J.; Peng, L.; et al. A Zebrafish Screen Reveals Renin-Angiotensin System Inhibitors as Neuroprotective Via Mitochondrial Restoration in Dopamine Neurons. Elife 2021, 10, e69795. [Google Scholar] [CrossRef]
- Podlasz, P.; Jakimiuk, A.; Kasica-Jarosz, N.; Czaja, K.; Wasowicz, K. Neuroanatomical Localization of Galanin in Zebrafish Telencephalon and Anticonvulsant Effect of Galanin Overexpression. ACS Chem. Neurosci. 2018, 9, 3049–3059. [Google Scholar] [CrossRef] [PubMed]
- Corradi, L.; Bruzzone, M.; Maschio, M.D.; Sawamiphak, S.; Filosa, A. Hypothalamic Galanin-Producing Neurons Regulate Stress in Zebrafish through a Peptidergic, Self-Inhibitory Loop. Curr. Biol. 2022, 32, 1497–1510.e5. [Google Scholar] [CrossRef]
- Zada, D.; Tovin, A.; Lerer-Goldshtein, T.; Vatine, G.D.; Appelbaum, L. Altered Behavioral Performance and Live Imaging of Circuit-Specific Neural Deficiencies in a Zebrafish Model for Psychomotor Retardation. PLoS Genet. 2014, 10, e1004615. [Google Scholar] [CrossRef] [PubMed]
- Choe, C.P.; Choi, S.Y.; Kee, Y.; Kim, M.J.; Kim, S.H.; Lee, Y.; Park, H.C.; Ro, H. Transgenic Fluorescent Zebrafish Lines That Have Revolutionized Biomedical Research. Lab. Anim. Res. 2021, 37, 26. [Google Scholar] [CrossRef]
- Paquet, D.; Bhat, R.; Sydow, A.; Mandelkow, E.M.; Berg, S.; Hellberg, S.; Falting, J.; Distel, M.; Koster, R.W.; Schmid, B.; et al. A Zebrafish Model of Tauopathy Allows in Vivo Imaging of Neuronal Cell Death and Drug Evaluation. J. Clin. Investig. 2009, 119, 1382–1395. [Google Scholar] [CrossRef] [PubMed]
- Goedert, M.; Eisenberg, D.S.; Crowther, R.A. Propagation of Tau Aggregates and Neurodegeneration. Annu. Rev. Neurosci. 2017, 40, 189–210. [Google Scholar] [CrossRef] [PubMed]
- Giustiniani, J.; Chambraud, B.; Sardin, E.; Dounane, O.; Guillemeau, K.; Nakatani, H.; Paquet, D.; Kamah, A.; Landrieu, I.; Lippens, G.; et al. Immunophilin Fkbp52 Induces Tau-P301l Filamentous Assembly In Vitro and Modulates Its Activity in a Model of Tauopathy. Proc. Natl. Acad. Sci. USA 2014, 111, 4584–4589. [Google Scholar] [CrossRef] [PubMed]
- Sepulveda-Diaz, J.E.; Naini, S.M.A.; Huynh, M.B.; Ouidja, M.O.; Yanicostas, C.; Chantepie, S.; Villares, J.; Lamari, F.; Jospin, E.; van Kuppevelt, T.H.; et al. Hs3st2 Expression Is Critical for the Abnormal Phosphorylation of Tau in Alzheimer’s Disease-Related Tau Pathology. Brain 2015, 138 Pt 5, 1339–1354. [Google Scholar] [CrossRef] [PubMed]
- Hassan-Abdi, R.; Brenet, A.; Bennis, M.; Yanicostas, C.; Soussi-Yanicostas, N. Neurons Expressing Pathological Tau Protein Trigger Dramatic Changes in Microglial Morphology and Dynamics. Front. Neurosci. 2019, 13, 1199. [Google Scholar] [CrossRef] [PubMed]
- Barbereau, C.; Yehya, A.; Silhol, M.; Cubedo, N.; Verdier, J.M.; Maurice, T.; Rossel, M. Neuroprotective Brain-Derived Neurotrophic Factor Signaling in the Tau-P301l Tauopathy Zebrafish Model. Pharmacol. Res. 2020, 158, 104865. [Google Scholar] [CrossRef]
- Jamadagni, P.; Breuer, M.; Schmeisser, K.; Cardinal, T.; Kassa, B.; Parker, J.A.; Pilon, N.; Samarut, E.; Patten, S.A. Chromatin Remodeller Chd7 Is Required for Gabaergic Neuron Development by Promoting Paqr3 Expression. EMBO Rep. 2021, 22, e50958. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.H.; Kim, S.; Chung, A.Y.; Kim, H.T.; So, J.H.; Ryu, J.; Park, H.C.; Kim, C.H. Visualization of Myelination in Gfp-Transgenic Zebrafish. Dev. Dyn. 2010, 239, 592–597. [Google Scholar] [CrossRef]
- Early, J.J.; Cole, K.L.; Williamson, J.M.; Swire, M.; Kamadurai, H.; Muskavitch, M.; Lyons, D.A. An Automated High-Resolution in Vivo Screen in Zebrafish to Identify Chemical Regulators of Myelination. Elife 2018, 7, e35136. [Google Scholar] [CrossRef]
- Chung, A.Y.; Kim, P.S.; Kim, S.; Kim, E.; Kim, D.; Jeong, I.; Kim, H.K.; Ryu, J.H.; Kim, C.H.; Choi, J.; et al. Generation of Demyelination Models by Targeted Ablation of Oligodendrocytes in the Zebrafish Cns. Mol. Cells 2013, 36, 82–87. [Google Scholar] [CrossRef]
- Pant, D.C.; Dorboz, I.; Schluter, A.; Fourcade, S.; Launay, N.; Joya, J.; Aguilera-Albesa, S.; Yoldi, M.E.; Casasnovas, C.; Willis, M.J.; et al. Loss of the Sphingolipid Desaturase Degs1 Causes Hypomyelinating Leukodystrophy. J. Clin. Investig. 2019, 129, 1240–1256. [Google Scholar] [CrossRef]
- Zada, D.; Tovin, A.; Lerer-Goldshtein, T.; Appelbaum, L. Pharmacological Treatment and Bbb-Targeted Genetic Therapy for Mct8-Dependent Hypomyelination in Zebrafish. Dis. Model. Mech. 2016, 9, 1339–1348. [Google Scholar]
- Xiao, Y.; Faucherre, A.; Pola-Morell, L.; Heddleston, J.M.; Liu, T.L.; Chew, T.L.; Sato, F.; Sehara-Fujisawa, A.; Kawakami, K.; Lopez-Schier, H. High-Resolution Live Imaging Reveals Axon-Glia Interactions During Peripheral Nerve Injury and Repair in Zebrafish. Dis. Model. Mech. 2015, 8, 553–564. [Google Scholar] [CrossRef]
- Xi, Y.; Yu, M.; Godoy, R.; Hatch, G.; Poitras, L.; Ekker, M. Transgenic Zebrafish Expressing Green Fluorescent Protein in Dopaminergic Neurons of the Ventral Diencephalon. Dev. Dyn. 2011, 240, 2539–2547. [Google Scholar] [CrossRef]
- Ren, Q.; Jiang, X.; Zhang, S.; Gao, X.; Paudel, Y.N.; Zhang, P.; Wang, R.; Liu, K.; Jin, M. Neuroprotective Effect of Yiaedaer Peptide against Parkinson’s Disease Like Pathology in Zebrafish. Biomed. Pharmacother. Biomed. Pharmacother. 2022, 147, 112629. [Google Scholar] [CrossRef] [PubMed]
- Stednitz, S.J.; Freshner, B.; Shelton, S.; Shen, T.; Black, D.; Gahtan, E. Selective Toxicity of L-Dopa to Dopamine Transporter-Expressing Neurons and Locomotor Behavior in Zebrafish Larvae. Neurotoxicol. Teratol. 2015, 52 Pt A, 51–56. [Google Scholar] [CrossRef]
- Godoy, R.; Noble, S.; Yoon, K.; Anisman, H.; Ekker, M. Chemogenetic Ablation of Dopaminergic Neurons Leads to Transient Locomotor Impairments in Zebrafish Larvae. J. Neurochem. 2015, 135, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Weston, L.J.; Cook, Z.T.; Stackhouse, T.L.; Sal, M.K.; Schultz, B.I.; Tobias, Z.J.C.; Osterberg, V.R.; Brockway, N.L.; Pizano, S.; Glover, G.; et al. In Vivo Aggregation of Presynaptic Alpha-Synuclein Is Not Influenced by Its Phosphorylation at Serine-129. Neurobiol. Dis. 2021, 152, 105291. [Google Scholar] [CrossRef] [PubMed]
- Lopez, A.; Gorb, A.; Palha, N.; Fleming, A.; Rubinsztein, D.C. A New Zebrafish Model to Measure Neuronal Alpha-Synuclein Clearance In Vivo. Genes 2022, 13, 868. [Google Scholar] [CrossRef] [PubMed]
- Franken, L.E.; Grunewald, K.; Boekema, E.J.; Stuart, M.C.A. A Technical Introduction to Transmission Electron Microscopy for Soft-Matter: Imaging, Possibilities, Choices, and Technical Developments. Small 2020, 16, e1906198. [Google Scholar] [CrossRef]
- Turcotte, R.; Rutledge, D.J.; Belanger, E.; Dill, D.; Macklin, W.B.; Cote, D.C. Intravital Assessment of Myelin Molecular Order with Polarimetric Multiphoton Microscopy. Sci. Rep. 2016, 6, 31685. [Google Scholar] [CrossRef]
- Aspatwar, A.; Tolvanen, M.E.; Jokitalo, E.; Parikka, M.; Ortutay, C.; Harjula, S.K.; Ramet, M.; Vihinen, M.; Parkkila, S. Abnormal Cerebellar Development and Ataxia in Carp Viii Morphant Zebrafish. Hum. Mol. Genet. 2013, 22, 417–432. [Google Scholar] [CrossRef]
- Marschall, S.; Sander, B.; Mogensen, M.; Jorgensen, T.M.; Andersen, P.E. Optical Coherence Tomography-Current Technology and Applications in Clinical and Biomedical Research. Anal. Bioanal. Chem. 2011, 400, 2699–2720. [Google Scholar] [CrossRef]
- Lichtenegger, A.; Baumann, B.; Yasuno, Y. Optical Coherence Tomography Is a Promising Tool for Zebrafish-Based Research—A Review. Bioengineering 2022, 10, 5. [Google Scholar] [CrossRef]
- Lichtenegger, A.; Mukherjee, P.; Tamaoki, J.; Bian, L.; Zhu, L.; El-Sadek, I.A.; Makita, S.; Leskovar, K.; Kobayashi, M.; Baumann, B.; et al. Multicontrast Investigation of In Vivo Wildtype Zebrafish in Three Development Stages Using Polarization-Sensitive Optical Coherence Tomography. J. Biomed. Opt. 2022, 27, 016001. [Google Scholar] [CrossRef] [PubMed]
- Lichtenegger, A.; Mukherjee, P.; Zhu, L.; Morishita, R.; Tomita, K.; Oida, D.; Leskovar, K.; El-Sadek, I.A.; Makita, S.; Kirchberger, S.; et al. Non-Destructive Characterization of Adult Zebrafish Models Using Jones Matrix Optical Coherence Tomography. Biomed. Opt. Express 2022, 13, 2202–2223. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Chen, T.; Mao, G.; Qiu, T.; Lan, Y.; Xiang, X.; Huang, J.; Huang, J.; Lu, T.; Gan, S.; et al. Long-Term and in Vivo Assessment of Abeta Protein-Induced Brain Atrophy in a Zebrafish Model by Optical Coherence Tomography. J. Biophotonics 2020, 13, e202000067. [Google Scholar] [CrossRef]
- Divakar Rao, K.; Upadhyaya, P.; Sharma, M.; Gupta, P.K. Noninvasive Imaging of Ethanol-Induced Developmental Defects in Zebrafish Embryos Using Optical Coherence Tomography. Birth Defects Res. Part B Dev. Reprod. Toxicol. 2012, 95, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.X. Coherent Anti-Stokes Raman Scattering Microscopy. Appl. Spectrosc. 2007, 61, 197–208. [Google Scholar] [CrossRef]
- de Vito, G.; Bifone, A.; Piazza, V. Rotating-Polarization Cars Microscopy: Combining Chemical and Molecular Orientation Sensitivity. Opt. Express 2012, 20, 29369–29377. [Google Scholar] [CrossRef] [PubMed]
- de Vito, G.; Tonazzini, I.; Cecchini, M.; Piazza, V. Rp-Cars: Label-Free Optical Readout of the Myelin Intrinsic Healthiness. Opt. Express 2014, 22, 13733–13743. [Google Scholar] [CrossRef]
- de Vito, G.; Cappello, V.; Tonazzini, I.; Cecchini, M.; Piazza, V. Rp-Cars Reveals Molecular Spatial Order Anomalies in Myelin of an Animal Model of Krabbe Disease. J. Biophotonics 2017, 10, 385–393. [Google Scholar] [CrossRef]
- Squier, J.; Muller, M.; Brakenhoff, G.; Wilson, K.R. Third Harmonic Generation Microscopy. Opt. Express 1998, 3, 315–324. [Google Scholar] [CrossRef]
- Chen, S.Y.; Hsieh, C.S.; Chu, S.W.; Lin, C.Y.; Ko, C.Y.; Chen, Y.C.; Tsai, H.J.; Hu, C.H.; Sun, C.K. Noninvasive Harmonics Optical Microscopy for Long-Term Observation of Embryonic Nervous System Development In Vivo. J. Biomed. Opt. 2006, 11, 054022. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.S.; Ko, C.Y.; Chen, S.Y.; Liu, T.M.; Wu, J.S.; Hu, C.H.; Sun, C.K. In Vivo Long-Term Continuous Observation of Gene Expression in Zebrafish Embryo Nerve Systems by Using Harmonic Generation Microscopy and Morphant Technology. J. Biomed. Opt. 2008, 13, 064041. [Google Scholar] [CrossRef]
- Farrar, M.J.; Wise, F.W.; Fetcho, J.R.; Schaffer, C.B. In Vivo Imaging of Myelin in the Vertebrate Central Nervous System Using Third Harmonic Generation Microscopy. Biophys. J. 2011, 100, 1362–1371. [Google Scholar] [CrossRef]
- Schermelleh, L.; Ferrand, A.; Huser, T.; Eggeling, C.; Sauer, M.; Biehlmaier, O.; Drummen, G.P.C. Super-Resolution Microscopy Demystified. Nat. Cell. Biol. 2019, 21, 72–84. [Google Scholar] [CrossRef]
- Turcotte, R.; Liang, Y.; Tanimoto, M.; Zhang, Q.; Li, Z.; Koyama, M.; Betzig, E.; Ji, N. Dynamic Super-Resolution Structured Illumination Imaging in the Living Brain. Proc. Natl. Acad. Sci. USA 2019, 116, 9586–9591. [Google Scholar] [CrossRef]
- Winter, P.W.; York, A.G.; Nogare, D.D.; Ingaramo, M.; Christensen, R.; Chitnis, A.; Patterson, G.H.; Shroff, H. Two-Photon Instant Structured Illumination Microscopy Improves the Depth Penetration of Super-Resolution Imaging in Thick Scattering Samples. Optica 2014, 1, 181–191. [Google Scholar] [CrossRef]
- York, A.G.; Parekh, S.H.; Nogare, D.D.; Fischer, R.S.; Temprine, K.; Mione, M.; Chitnis, A.B.; Combs, C.A.; Shroff, H. Resolution Doubling in Live, Multicellular Organisms Via Multifocal Structured Illumination Microscopy. Nat. Methods 2012, 9, 749–754. [Google Scholar] [CrossRef]
- Klar, T.A.; Jakobs, S.; Dyba, M.; Egner, A.; Hell, S.W. Fluorescence Microscopy with Diffraction Resolution Barrier Broken by Stimulated Emission. Proc. Natl. Acad. Sci. USA 2000, 97, 8206–8210. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.; Gould, T.J.; Bewersdorf, J.; Zenisek, D. High-Resolution Optical Imaging of Zebrafish Larval Ribbon Synapse Protein Ribeye, Rim2, and Cav 1.4 by Stimulation Emission Depletion Microscopy. Microsc. Microanal. 2012, 18, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Keklikoglou, K.; Arvanitidis, C.; Chatzigeorgiou, G.; Chatzinikolaou, E.; Karagiannidis, E.; Koletsa, T.; Magoulas, A.; Makris, K.; Mavrothalassitis, G.; Papanagnou, E.D.; et al. Micro-Ct for Biological and Biomedical Studies: A Comparison of Imaging Techniques. J. Imaging 2021, 7, 172. [Google Scholar] [CrossRef] [PubMed]
- Rohner, T.C.; Staab, D.; Stoeckli, M. Maldi Mass Spectrometric Imaging of Biological Tissue Sections. Mech. Ageing Dev. 2005, 126, 177–185. [Google Scholar] [CrossRef]
- Ding, Y.; Vanselow, D.J.; Yakovlev, M.A.; Katz, S.R.; Lin, A.Y.; Clark, D.P.; Vargas, P.; Xin, X.; Copper, J.E.; Canfield, V.A.; et al. Computational 3d Histological Phenotyping of Whole Zebrafish by X-Ray Histotomography. Elife 2019, 8, e44898. [Google Scholar] [CrossRef] [PubMed]
- Asslan, M.; Lauzon, N.; Beus, M.; Maysinger, D.; Rousseau, S. Mass Spectrometry Imaging in Zebrafish Larvae for Assessing Drug Safety and Metabolism. Anal. Bioanal. Chem. 2021, 413, 5135–5146. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.M.; Meyer, M.R.; Muller, R.; Herrmann, J. Drug Administration Routes Impact the Metabolism of a Synthetic Cannabinoid in the Zebrafish Larvae Model. Molecules 2020, 25, 4474. [Google Scholar] [CrossRef]
- Villacrez, M.; Hellman, K.; Ono, T.; Sugihara, Y.; Rezeli, M.; Ek, F.; Marko-Varga, G.; Olsson, R. Evaluation of Drug Exposure and Metabolism in Locust and Zebrafish Brains Using Mass Spectrometry Imaging. ACS Chem. Neurosci. 2018, 9, 1994–2000. [Google Scholar] [CrossRef] [PubMed]
- Saint-Amant, L.; Drapeau, P. Time Course of the Development of Motor Behaviors in the Zebrafish Embryo. J. Neurobiol. 1998, 37, 622–632. [Google Scholar] [CrossRef]
- Joo, W.; Vivian, M.D.; Graham, B.J.; Soucy, E.R.; Thyme, S.B. A Customizable Low-Cost System for Massively Parallel Zebrafish Behavioral Phenotyping. Front. Behav. Neurosci. 2020, 14, 606900. [Google Scholar] [CrossRef]
- Patton, E.E.; Zon, L.I.; Langenau, D.M. Zebrafish Disease Models in Drug Discovery: From Preclinical Modelling to Clinical Trials. Nat. Reviews. Drug Discov. 2021, 20, 611–628. [Google Scholar] [CrossRef]
- Wang, L.; Sheng, W.; Tan, Z.; Ren, Q.; Wang, R.; Stoika, R.; Liu, X.; Liu, K.; Shang, X.; Jin, M. Treatment of Parkinson’s Disease in Zebrafish Model with a Berberine Derivative Capable of Crossing Blood Brain Barrier, Targeting Mitochondria, and Convenient for Bioimaging Experiments. Comp. Biochem. Physiol. Toxicol. Pharmacol. CBP 2021, 249, 109151. [Google Scholar] [CrossRef]
- Richendrfer, H.; Pelkowski, S.D.; Colwill, R.M.; Creton, R. On the Edge: Pharmacological Evidence for Anxiety-Related Behavior in Zebrafish Larvae. Behav. Brain Res. 2012, 228, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Wasilewska, I.; Gupta, R.K.; Wojtas, B.; Palchevska, O.; Kuznicki, J. Stim2b Knockout Induces Hyperactivity and Susceptibility to Seizures in Zebrafish Larvae. Cells 2020, 9, 1285. [Google Scholar] [CrossRef]
- Suo, G.; Cao, X.; Zheng, Y.; Li, H.; Zhang, Q.; Tang, J.; Wu, Y. A De Novo Nonsense Mutation of Stxbp1 Causes Early-Onset Epileptic Encephalopathy. Epilepsy Behav. 2021, 123, 108245. [Google Scholar] [CrossRef] [PubMed]
- Dinday, M.T.; Baraban, S.C. Large-Scale Phenotype-Based Antiepileptic Drug Screening in a Zebrafish Model of Dravet Syndrome. eNeuro 2015, 2, ENEURO.0068-15.2015. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, J.; Champagne, N.; Lafreniere, R.G.; Xiong, L.; Spiegelman, D.; Brustein, E.; Lapointe, M.; Peng, H.; Cote, M.; Noreau, A.; et al. De Novo Mutations in the Gene Encoding the Synaptic Scaffolding Protein Shank3 in Patients Ascertained for Schizophrenia. Proc. Natl. Acad. Sci. USA 2010, 107, 7863–7868. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Kim, J.; Marques, J.C.; Grama, A.; Hildebrand, D.G.C.; Gu, W.; Li, J.M.; Robson, D.N. Pan-Neuronal Calcium Imaging with Cellular Resolution in Freely Swimming Zebrafish. Nat. Methods 2017, 14, 1107–1114. [Google Scholar] [CrossRef] [PubMed]
- Colwill, R.M.; Creton, R. Imaging Escape and Avoidance Behavior in Zebrafish Larvae. Rev. Neurosci. 2011, 22, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Cognato Gde, P.; Bortolotto, J.W.; Blazina, A.R.; Christoff, R.R.; Lara, D.R.; Vianna, M.R.; Bonan, C.D. Y-Maze Memory Task in Zebrafish (Danio Rerio): The Role of Glutamatergic and Cholinergic Systems on the Acquisition and Consolidation Periods. Neurobiol. Learn. Mem. 2012, 98, 321–328. [Google Scholar] [CrossRef]
- Boiangiu, R.S.; Mihasan, M.; Gorgan, D.L.; Stache, B.A.; Hritcu, L. Anxiolytic, Promnesic, Anti-Acetylcholinesterase and Antioxidant Effects of Cotinine and 6-Hydroxy-L-Nicotine in Scopolamine-Induced Zebrafish (Danio Rerio) Model of Alzheimer’s Disease. Antioxidants 2021, 10, 212. [Google Scholar] [CrossRef]
- Bloom, H.D.; Perlmutter, A. A Sexual Aggregating Pheromone System in the Zebrafish, Brachydanio Rerio (Hamilton-Buchanan). J. Exp. Zool. 1977, 199, 215–226. [Google Scholar] [CrossRef]
- Al-Imari, L.; Gerlai, R. Sight of Conspecifics as Reward in Associative Learning in Zebrafish (Danio Rerio). Behav. Brain Res. 2008, 189, 216–219. [Google Scholar] [CrossRef]
- Varga, Z.K.; Zsigmond, A.; Pejtsik, D.; Varga, M.; Demeter, K.; Mikics, E.; Haller, J.; Aliczki, M. The Swimming Plus-Maze Test: A Novel High-Throughput Model for Assessment of Anxiety-Related Behaviour in Larval and Juvenile Zebrafish (Danio Rerio). Sci. Rep. 2018, 8, 16590. [Google Scholar] [CrossRef] [PubMed]
- Engeszer, R.E.; Ryan, M.J.; Parichy, D.M. Learned Social Preference in Zebrafish. Curr. Biol. 2004, 14, 881–884. [Google Scholar] [CrossRef] [PubMed]
- Ogi, A.; Licitra, R.; Naef, V.; Marchese, M.; Fronte, B.; Gazzano, A.; Santorelli, F.M. Social Preference Tests in Zebrafish: A Systematic Review. Front. Vet. Sci. 2020, 7, 590057. [Google Scholar] [CrossRef]
- Gomez-Laplaza, L.M.; Gerlai, R. Latent Learning in Zebrafish (Danio Rerio). Behav. Brain Res. 2010, 208, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, F.F.; Gaspary, K.V.; Leite, C.E.; De Paula Cognato, G.; Bonan, C.D. Embryological Exposure to Valproic Acid Induces Social Interaction Deficits in Zebrafish (Danio Rerio): A Developmental Behavior Analysis. Neurotoxicol. Teratol. 2015, 52 Pt A, 36–41. [Google Scholar] [CrossRef]
- Dwivedi, S.; Medishetti, R.; Rani, R.; Sevilimedu, A.; Kulkarni, P.; Yogeeswari, P. Larval Zebrafish Model for Studying the Effects of Valproic Acid on Neurodevelopment: An Approach towards Modeling Autism. J. Pharm. Toxicol. Methods 2019, 95, 56–65. [Google Scholar] [CrossRef]
- Fulcher, N.; Tran, S.; Shams, S.; Chatterjee, D.; Gerlai, R. Neurochemical and Behavioral Responses to Unpredictable Chronic Mild Stress Following Developmental Isolation: The Zebrafish as a Model for Major Depression. Zebrafish 2017, 14, 23–34. [Google Scholar] [CrossRef]
- Kumari, S.; Dhiman, P.; Kumar, R.; Rahmatkar, S.N.; Singh, D. Chemo-Kindling in Adult Zebrafish Alters Spatial Cognition but Not Social Novelty Recognition. Behav. Brain Res. 2023, 438, 114158. [Google Scholar] [CrossRef]
- Jarosova, R.; Niyangoda, S.S.; Hettiarachchi, P.; Johnson, M.A. Impaired Dopamine Release and Latent Learning in Alzheimer’s Disease Model Zebrafish. ACS Chem. Neurosci. 2022, 13, 2924–2931. [Google Scholar] [CrossRef]
- Hettiarachchi, P.; Niyangoda, S.S.; Jarosova, R.; Johnson, M.A. Dopamine Release Impairments Accompany Locomotor and Cognitive Deficiencies in Rotenone-Treated Parkinson’s Disease Model Zebrafish. Chem. Res. Toxicol. 2022, 35, 1974–1982. [Google Scholar] [CrossRef]
- Cleal, M.; Fontana, B.D.; Double, M.; Mezabrovschi, R.; Parcell, L.; Redhead, E.; Parker, M.O. Dopaminergic Modulation of Working Memory and Cognitive Flexibility in a Zebrafish Model of Aging-Related Cognitive Decline. Neurobiol. Aging 2021, 102, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Cachat, J.; Stewart, A.; Utterback, E.; Hart, P.; Gaikwad, S.; Wong, K.; Kyzar, E.; Wu, N.; Kalueff, A.V. Three-Dimensional Neurophenotyping of Adult Zebrafish Behavior. PLoS ONE 2011, 6, e17597. [Google Scholar] [CrossRef] [PubMed]
- Egan, R.J.; Bergner, C.L.; Hart, P.C.; Cachat, J.M.; Canavello, P.R.; Elegante, M.F.; Elkhayat, S.I.; Bartels, B.K.; Tien, A.K.; Tien, D.H.; et al. Understanding Behavioral and Physiological Phenotypes of Stress and Anxiety in Zebrafish. Behav. Brain Res. 2009, 205, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Maaswinkel, H.; Zhu, L.; Weng, W. Assessing Social Engagement in Heterogeneous Groups of Zebrafish: A New Paradigm for Autism-Like Behavioral Responses. PLoS ONE 2013, 8, e75955. [Google Scholar] [CrossRef]
- Kim, L.; He, L.; Maaswinkel, H.; Zhu, L.; Sirotkin, H.; Weng, W. Anxiety, Hyperactivity and Stereotypy in a Zebrafish Model of Fragile X Syndrome and Autism Spectrum Disorder. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 55, 40–49. [Google Scholar] [CrossRef]
- Huisken, J.; Swoger, J.; Del Bene, F.; Wittbrodt, J.; Stelzer, E.H. Optical Sectioning Deep inside Live Embryos by Selective Plane Illumination Microscopy. Science 2004, 305, 1007–1009. [Google Scholar] [CrossRef]
- Fahrbach, F.O.; Voigt, F.F.; Schmid, B.; Helmchen, F.; Huisken, J. Rapid 3d Light-Sheet Microscopy with a Tunable Lens. Opt. Express 2013, 21, 21010–21026. [Google Scholar] [CrossRef]
- Royer, L.A.; Lemon, W.C.; Chhetri, R.K.; Wan, Y.; Coleman, M.; Myers, E.W.; Keller, P.J. Adaptive Light-Sheet Microscopy for Long-Term, High-Resolution Imaging in Living Organisms. Nat. Biotechnol. 2016, 34, 1267–1278. [Google Scholar] [CrossRef]
- Sancataldo, G.; Gavryusev, V.; de Vito, G.; Turrini, L.; Locatelli, M.; Fornetto, C.; Tiso, N.; Vanzi, F.; Silvestri, L.; Pavone, F.S. Flexible Multi-Beam Light-Sheet Fluorescence Microscope for Live Imaging without Striping Artifacts. Front. Neuroanat. 2019, 13, 7. [Google Scholar] [CrossRef]
- Gavryusev, V.; Sancataldo, G.; Ricci, P.; Montalbano, A.; Fornetto, C.; Turrini, L.; Laurino, A.; Pesce, L.; de Vito, G.; Tiso, N.; et al. Dual-Beam Confocal Light-Sheet Microscopy Via Flexible Acousto-Optic Deflector. J. Biomed. Opt. 2019, 24, 1–6. [Google Scholar] [CrossRef]
- de Vito, G.; Ricci, P.; Turrini, L.; Gavryusev, V.; Mullenbroich, M.C.; Tiso, N.; Vanzi, F.; Silvestri, L.; Pavone, F. Effects of Excitation Light Polarization on Fluorescence Emission in Two-Photon Light-Sheet Microscopy. Biomed. Opt. Express 2020, 11, 4651–4665. [Google Scholar] [CrossRef]
- Ricci, P.; Gavryusev, V.; Mullenbroich, C.; Turrini, L.; de Vito, G.; Silvestri, L.; Sancataldo, G.; Pavone, F.S. Removing Striping Artifacts in Light-Sheet Fluorescence Microscopy: A Review. Prog. Biophys. Mol. Biol. 2022, 168, 52–65. [Google Scholar] [CrossRef]
- Tomer, R.; Lovett-Barron, M.; Kauvar, I.; Andalman, A.; Burns, V.M.; Sankaran, S.; Grosenick, L.; Broxton, M.; Yang, S.; Deisseroth, K. Sped Light Sheet Microscopy: Fast Mapping of Biological System Structure and Function. Cell 2015, 163, 1796–1806. [Google Scholar] [CrossRef]
- Randlett, O.; Wee, C.L.; Naumann, E.A.; Nnaemeka, O.; Schoppik, D.; Fitzgerald, J.E.; Portugues, R.; Lacoste, A.M.; Riegler, C.; Engert, F.; et al. Whole-Brain Activity Mapping onto a Zebrafish Brain Atlas. Nat. Methods 2015, 12, 1039–1046. [Google Scholar] [CrossRef]
- Kunst, M.; Laurell, E.; Mokayes, N.; Kramer, A.; Kubo, F.; Fernandes, A.M.; Forster, D.; Maschio, M.D.; Baier, H. A Cellular-Resolution Atlas of the Larval Zebrafish Brain. Neuron 2019, 103, 21–38.e5. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, D.G.C.; Cicconet, M.; Torres, R.M.; Choi, W.; Quan, T.M.; Moon, J.; Wetzel, A.W.; Champion, A.S.; Graham, B.J.; Randlett, O.; et al. Whole-Brain Serial-Section Electron Microscopy in Larval Zebrafish. Nature 2017, 545, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Svara, F.; Forster, D.; Kubo, F.; Januszewski, M.; Maschio, M.D.; Schubert, P.J.; Kornfeld, J.; Wanner, A.A.; Laurell, E.; Denk, W.; et al. Automated Synapse-Level Reconstruction of Neural Circuits in the Larval Zebrafish Brain. Nat. Methods 2022, 19, 1357–1366. [Google Scholar] [CrossRef]
- Johnson, R.E.; Linderman, S.; Panier, T.; Wee, C.L.; Song, E.; Herrera, K.J.; Miller, A.; Engert, F. Probabilistic Models of Larval Zebrafish Behavior Reveal Structure on Many Scales. Curr. Biol. 2020, 30, 70–82.e4. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zhang, C.; Lischinsky, J.E.; Jing, M.; Zhou, J.; Wang, H.; Zhang, Y.; Dong, A.; Wu, Z.; Wu, H.; et al. A Genetically Encoded Fluorescent Sensor for Rapid and Specific in Vivo Detection of Norepinephrine. Neuron 2019, 102, 745–761.e8. [Google Scholar] [CrossRef] [PubMed]
- Marvin, J.S.; Shimoda, Y.; Magloire, V.; Leite, M.; Kawashima, T.; Jensen, T.P.; Kolb, I.; Knott, E.L.; Novak, O.; Podgorski, K.; et al. A Genetically Encoded Fluorescent Sensor for in Vivo Imaging of Gaba. Nat. Methods 2019, 16, 763–770. [Google Scholar] [CrossRef]
- Kubitschke, M.; Muller, M.; Wallhorn, L.; Pulin, M.; Mittag, M.; Pollok, S.; Ziebarth, T.; Bremshey, S.; Gerdey, J.; Claussen, K.C.; et al. Next Generation Genetically Encoded Fluorescent Sensors for Serotonin. Nat. Commun 2022, 13, 7525. [Google Scholar] [CrossRef]
- Yizhar, O.; Fenno, L.E.; Davidson, T.J.; Mogri, M.; Deisseroth, K. Optogenetics in Neural Systems. Neuron 2011, 71, 9–34. [Google Scholar] [CrossRef]
- Boyden, E.S. Optogenetics and the Future of Neuroscience. Nat. Neurosci. 2015, 18, 1200–1201. [Google Scholar] [CrossRef]
- Ricci, P.; Marchetti, M.; Sorelli, M.; Turrini, L.; Resta, F.; Gavryusev, V.; de Vito, G.; Sancataldo, G.; Vanzi, F.; Silvestri, L.; et al. Power-Effective Scanning with Aods for 3d Optogenetic Applications. J. Biophotonics 2022, 15, e202100256. [Google Scholar] [CrossRef]
- Cong, L.; Wang, Z.; Chai, Y.; Hang, W.; Shang, C.; Yang, W.; Bai, L.; Du, J.; Wang, K.; Wen, Q. Rapid Whole Brain Imaging of Neural Activity in Freely Behaving Larval Zebrafish (Danio Rerio). Elife 2017, 6, e28158. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turrini, L.; Roschi, L.; de Vito, G.; Pavone, F.S.; Vanzi, F. Imaging Approaches to Investigate Pathophysiological Mechanisms of Brain Disease in Zebrafish. Int. J. Mol. Sci. 2023, 24, 9833. https://doi.org/10.3390/ijms24129833
Turrini L, Roschi L, de Vito G, Pavone FS, Vanzi F. Imaging Approaches to Investigate Pathophysiological Mechanisms of Brain Disease in Zebrafish. International Journal of Molecular Sciences. 2023; 24(12):9833. https://doi.org/10.3390/ijms24129833
Chicago/Turabian StyleTurrini, Lapo, Lorenzo Roschi, Giuseppe de Vito, Francesco Saverio Pavone, and Francesco Vanzi. 2023. "Imaging Approaches to Investigate Pathophysiological Mechanisms of Brain Disease in Zebrafish" International Journal of Molecular Sciences 24, no. 12: 9833. https://doi.org/10.3390/ijms24129833
APA StyleTurrini, L., Roschi, L., de Vito, G., Pavone, F. S., & Vanzi, F. (2023). Imaging Approaches to Investigate Pathophysiological Mechanisms of Brain Disease in Zebrafish. International Journal of Molecular Sciences, 24(12), 9833. https://doi.org/10.3390/ijms24129833