
Beta-Secretase 1 Recruits Amyloid-Beta Precursor Protein to ROCK2 Kinase, Resulting in Erroneous Phosphorylation and Beta-Amyloid Plaque Formation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
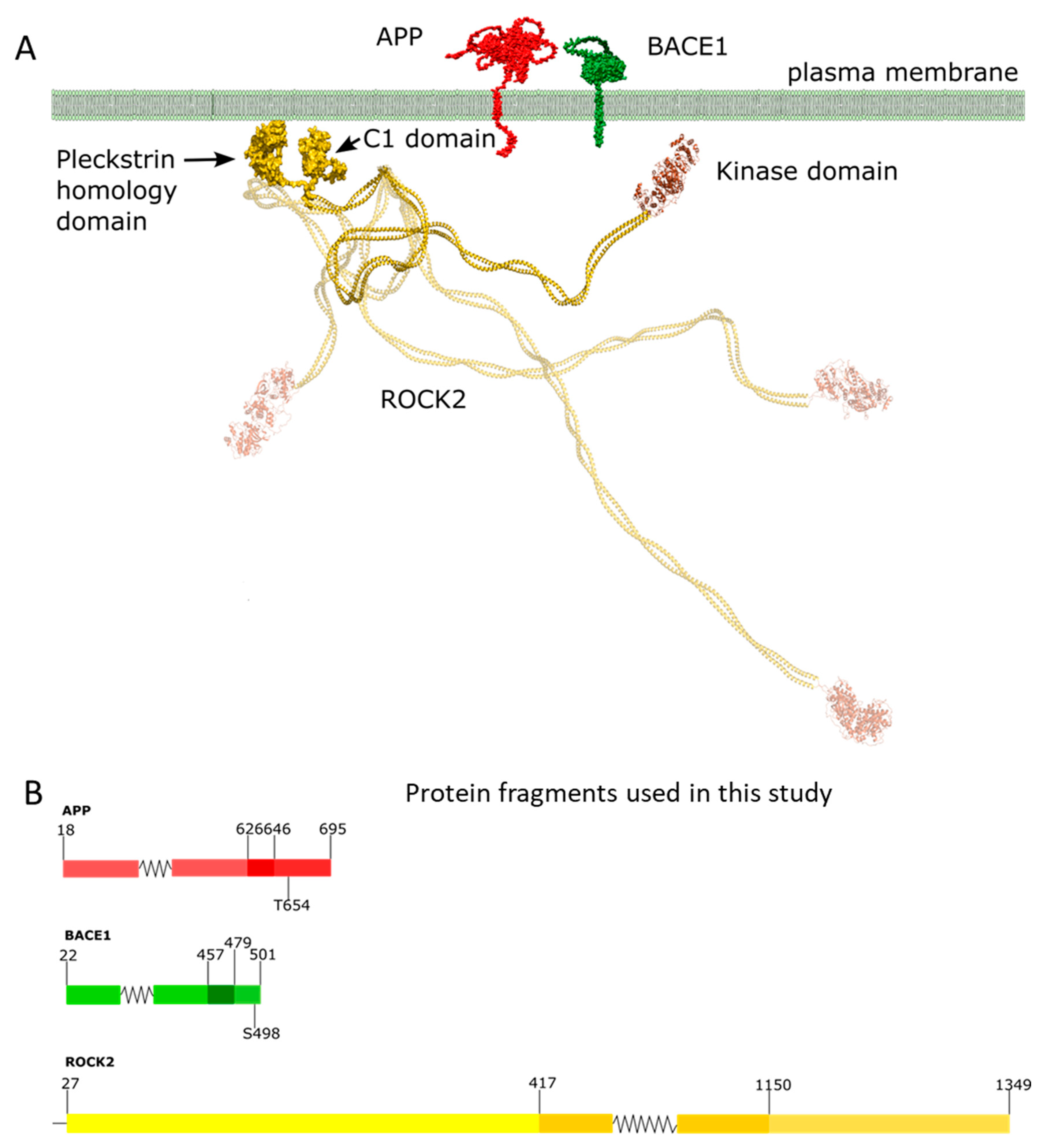
:1. Introduction
2. Results
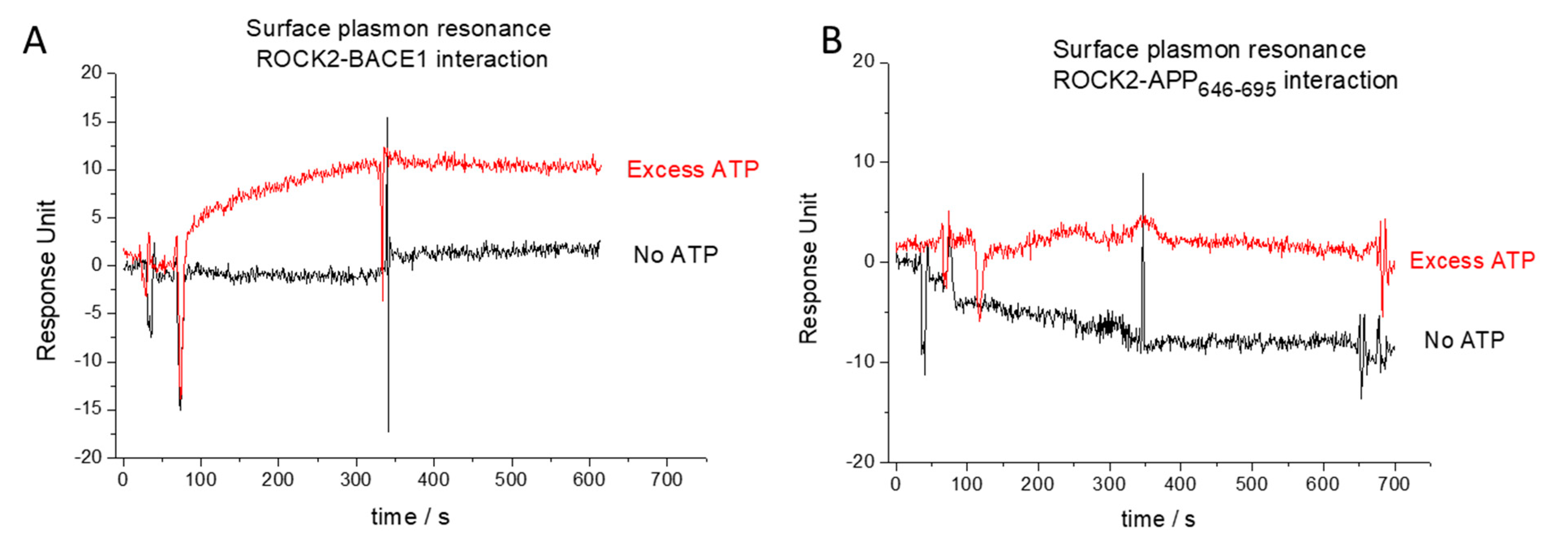
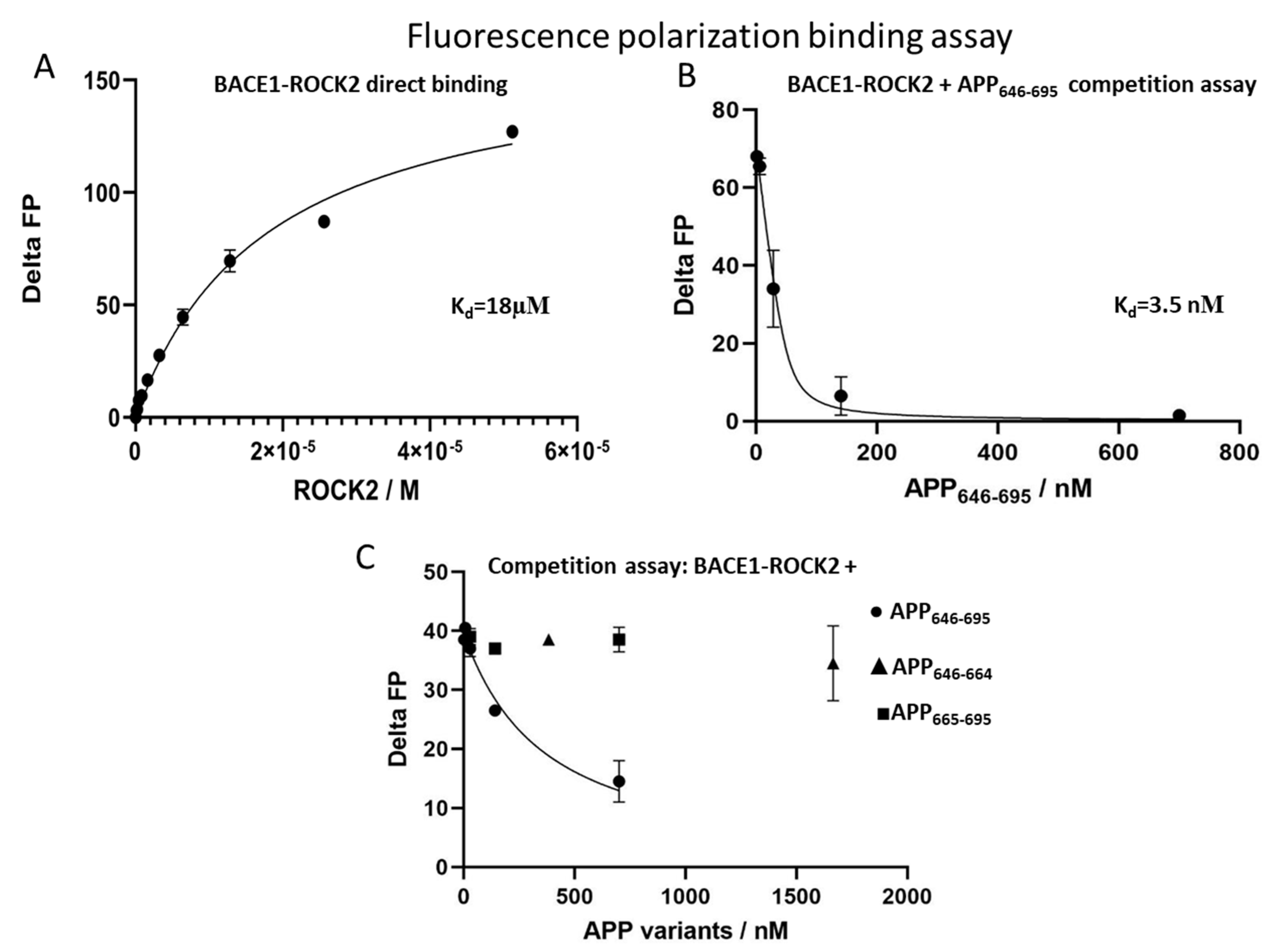
2.1. BACE1-ROCK2 Binary Interaction
2.2. APP-ROCK2 Binary Interaction
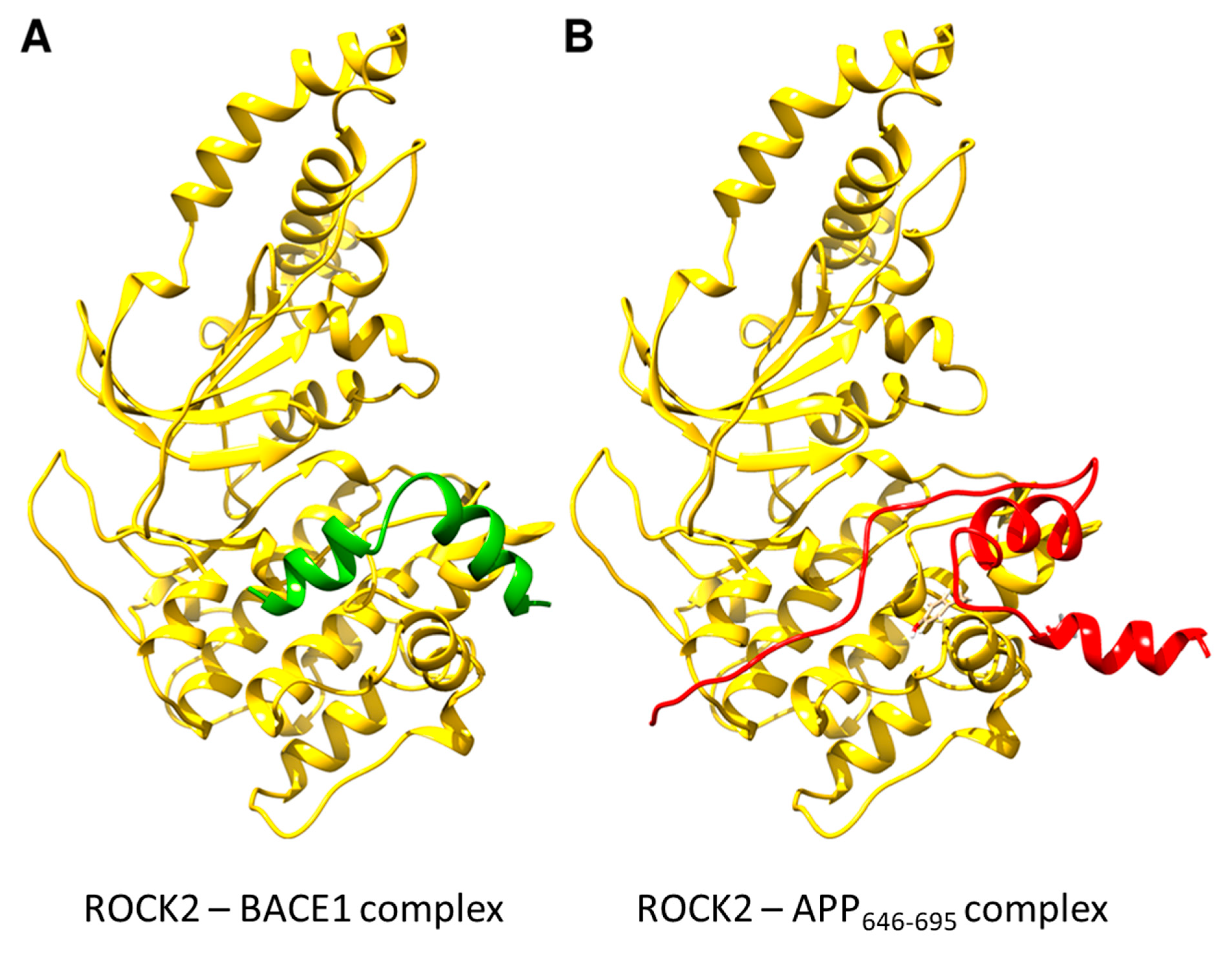
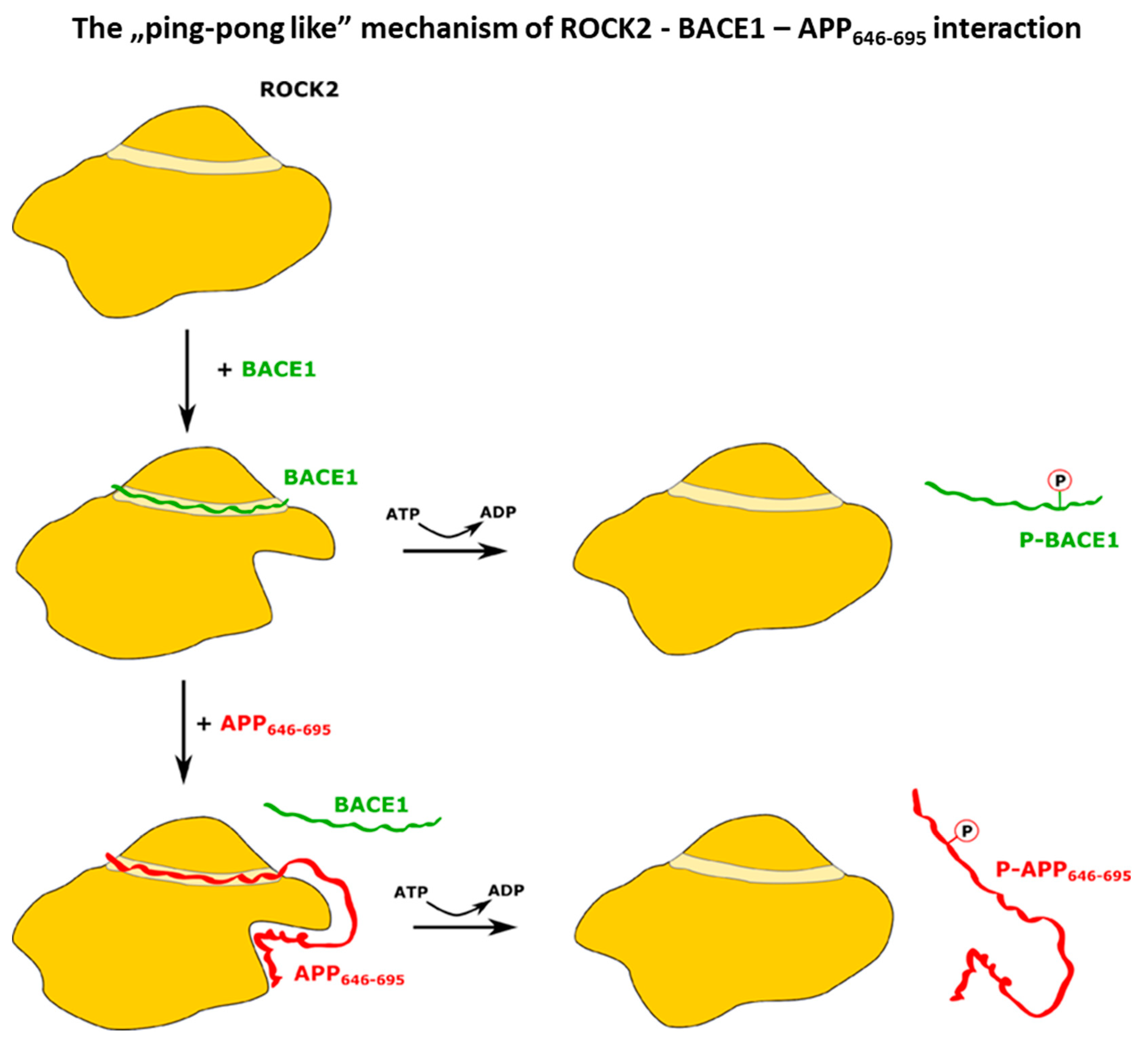
2.3. In Search of a ROCK2-BACE1-APP Ternary Interaction
3. Discussion
4. Materials and Methods
4.1. Protein Expression and Purification
4.2. Peptides
4.3. Fluorescence Labeling
4.4. Fluorescence Polarization
4.5. Surface Plasmon Resonance (SPR)
4.6. Modeling of the Binary Complexes
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hampel, H.; Hardy, J.; Blennow, K.; Chen, C.; Perry, G.; Kim, S.H.; Villemagne, V.L.; Aisen, P.; Vendruscolo, M.; Iwatsubo, T.; et al. The Amyloid-β Pathway in Alzheimer’s Disease. Mol. Psychiatry 2021, 26, 5481–5503. [Google Scholar] [CrossRef]
- Walsh, S.; Merrick, R.; Milne, R.; Brayne, C. Aducanumab for Alzheimer’s Disease? BMJ 2021, 374, n1682. [Google Scholar] [CrossRef] [PubMed]
- Kojro, E.; Fahrenholz, F. The Non-Amyloidogenic Pathway: Structure and Function of Alpha-Secretases. Subcell Biochem. 2005, 38, 105–127. [Google Scholar] [CrossRef]
- Cole, S.L.; Vassar, R. The Alzheimer’s Disease β-Secretase Enzyme, BACE1. Mol. Neurodegener. 2007, 2, 22. [Google Scholar] [CrossRef] [Green Version]
- Bai, X.; Yan, C.; Yang, G.; Lu, P.; Ma, D.; Sun, L.; Zhou, R.; Scheres, S.H.W.; Shi, Y. An Atomic Structure of Human γ-Secretase. Nature 2015, 525, 212–217. [Google Scholar] [CrossRef] [Green Version]
- Choy, R.W.-Y.; Cheng, Z.; Schekman, R. Amyloid Precursor Protein (APP) Traffics from the Cell Surface via Endosomes for Amyloid β (Aβ) Production in the Trans-Golgi Network. Proc. Natl. Acad. Sci. USA 2012, 109, E2077–E2082. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Chen, D.; Lee, T.H. Phosphorylation Signaling in APP Processing in Alzheimer’s Disease. Int. J. Mol. Sci. 2019, 21, E209. [Google Scholar] [CrossRef] [Green Version]
- Herskowitz, J.H.; Feng, Y.; Mattheyses, A.L.; Hales, C.M.; Higginbotham, L.A.; Duong, D.M.; Montine, T.J.; Troncoso, J.C.; Thambisetty, M.; Seyfried, N.T.; et al. Pharmacologic Inhibition of ROCK2 Suppresses Amyloid-β Production in an Alzheimer’s Disease Mouse Model. J. Neurosci. 2013, 33, 19086–19098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Yin, Y.; Weiser, A.; Griffin, E.; Cameron, M.D.; Lin, L.; Ruiz, C.; Schürer, S.C.; Inoue, T.; Rao, P.V.; et al. Discovery of Substituted 4-(Pyrazol-4-Yl)-Phenylbenzodioxane-2-Carboxamides as Potent and Highly Selective Rho Kinase (ROCK-II) Inhibitors. J. Med. Chem. 2008, 51, 6642–6645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loirand, G. Rho Kinases in Health and Disease: From Basic Science to Translational Research. Pharmacol. Rev. 2015, 67, 1074–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, W.; Liu, W.; Yan, J.; Zhang, M. Structure Basis and Unconventional Lipid Membrane Binding Properties of the PH-C1 Tandem of Rho Kinases. J. Biol. Chem. 2008, 283, 26263–26273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truebestein, L.; Elsner, D.J.; Fuchs, E.; Leonard, T.A. A Molecular Ruler Regulates Cytoskeletal Remodelling by the Rho Kinases. Nat. Commun. 2015, 6, 10029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajdú, I.; Szilágyi, A.; Végh, B.M.; Wacha, A.; Györffy, D.; Gráczer, É.; Somogyi, M.; Gál, P.; Závodszky, P. Ligand-Induced Conformational Rearrangements Regulate the Switch between Membrane-Proximal and Distal Functions of Rho Kinase 2. Commun. Biol. 2020, 3, 721. [Google Scholar] [CrossRef]
- Goate, A.; Chartier-Harlin, M.C.; Mullan, M.; Brown, J.; Crawford, F.; Fidani, L.; Giuffra, L.; Haynes, A.; Irving, N.; James, L. Segregation of a Missense Mutation in the Amyloid Precursor Protein Gene with Familial Alzheimer’s Disease. Nature 1991, 349, 704–706. [Google Scholar] [CrossRef] [PubMed]
- Rohan de Silva, H.A.; Jen, A.; Wickenden, C.; Jen, L.S.; Wilkinson, S.L.; Patel, A.J. Cell-Specific Expression of Beta-Amyloid Precursor Protein Isoform MRNAs and Proteins in Neurons and Astrocytes. Brain Res. Mol. Brain Res. 1997, 47, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Vassar, R.; Bennett, B.D.; Babu-Khan, S.; Kahn, S.; Mendiaz, E.A.; Denis, P.; Teplow, D.B.; Ross, S.; Amarante, P.; Loeloff, R.; et al. Beta-Secretase Cleavage of Alzheimer’s Amyloid Precursor Protein by the Transmembrane Aspartic Protease BACE. Science 1999, 286, 735–741. [Google Scholar] [CrossRef] [Green Version]
- Sastre, M.; Steiner, H.; Fuchs, K.; Capell, A.; Multhaup, G.; Condron, M.M.; Teplow, D.B.; Haass, C. Presenilin-Dependent Gamma-Secretase Processing of Beta-Amyloid Precursor Protein at a Site Corresponding to the S3 Cleavage of Notch. EMBO Rep. 2001, 2, 835–841. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Mao, G.; Tan, J.; Dong, Y.; Cui, M.-Z.; Kim, S.-H.; Xu, X. Identification of a New Presenilin-Dependent Zeta-Cleavage Site within the Transmembrane Domain of Amyloid Precursor Protein. J. Biol. Chem. 2004, 279, 50647–50650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Strooper, B.; Saftig, P.; Craessaerts, K.; Vanderstichele, H.; Guhde, G.; Annaert, W.; Von Figura, K.; Van Leuven, F. Deficiency of Presenilin-1 Inhibits the Normal Cleavage of Amyloid Precursor Protein. Nature 1998, 391, 387–390. [Google Scholar] [CrossRef]
- Lee, M.-S.; Kao, S.-C.; Lemere, C.A.; Xia, W.; Tseng, H.-C.; Zhou, Y.; Neve, R.; Ahlijanian, M.K.; Tsai, L.-H. APP Processing Is Regulated by Cytoplasmic Phosphorylation. J. Cell Biol. 2003, 163, 83–95. [Google Scholar] [CrossRef]
- Vieira, S.I.; Rebelo, S.; Domingues, S.C.; da Cruz e Silva, E.F.; da Cruz e Silva, O.A.B. S655 Phosphorylation Enhances APP Secretory Traffic. Mol. Cell. Biochem. 2009, 328, 145–154. [Google Scholar] [CrossRef]
- Hu, Y.-B.; Ren, R.-J.; Zhang, Y.-F.; Huang, Y.; Cui, H.-L.; Ma, C.; Qiu, W.-Y.; Wang, H.; Cui, P.-J.; Chen, H.-Z.; et al. Rho-Associated Coiled-Coil Kinase 1 Activation Mediates Amyloid Precursor Protein Site-Specific Ser655 Phosphorylation and Triggers Amyloid Pathology. Aging Cell 2019, 18, e13001. [Google Scholar] [CrossRef]
- Aplin, A.E.; Gibb, G.M.; Jacobsen, J.S.; Gallo, J.M.; Anderton, B.H. In Vitro Phosphorylation of the Cytoplasmic Domain of the Amyloid Precursor Protein by Glycogen Synthase Kinase-3beta. J. Neurochem. 1996, 67, 699–707. [Google Scholar] [CrossRef]
- Iijima, K.; Ando, K.; Takeda, S.; Satoh, Y.; Seki, T.; Itohara, S.; Greengard, P.; Kirino, Y.; Nairn, A.C.; Suzuki, T. Neuron-Specific Phosphorylation of Alzheimer’s Beta-Amyloid Precursor Protein by Cyclin-Dependent Kinase 5. J. Neurochem. 2000, 75, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Standen, C.L.; Brownlees, J.; Grierson, A.J.; Kesavapany, S.; Lau, K.F.; McLoughlin, D.M.; Miller, C.C. Phosphorylation of Thr(668) in the Cytoplasmic Domain of the Alzheimer’s Disease Amyloid Precursor Protein by Stress-Activated Protein Kinase 1b (Jun N-Terminal Kinase-3). J. Neurochem. 2001, 76, 316–320. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.M.; You, M.-H.; Chen, C.-H.; Suh, J.; Tanzi, R.E.; Ho Lee, T. Inhibition of Death-Associated Protein Kinase 1 Attenuates the Phosphorylation and Amyloidogenic Processing of Amyloid Precursor Protein. Hum. Mol. Genet. 2016, 25, 2498–2513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.-C.; Zhang, W.; Chua, L.-L.; Chai, C.; Li, R.; Lin, L.; Cao, Z.; Angeles, D.C.; Stanton, L.W.; Peng, J.-H.; et al. Phosphorylation of Amyloid Precursor Protein by Mutant LRRK2 Promotes AICD Activity and Neurotoxicity in Parkinson’s Disease. Sci. Signal. 2017, 10, eaam6790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Lee, J.S.; Lee, K.J.; Turner, R.S.; Hoe, H.-S.; Pak, D.T.S. Polo-like Kinase 2 Phosphorylation of Amyloid Precursor Protein Regulates Activity-Dependent Amyloidogenic Processing. Neuropharmacology 2017, 117, 387–400. [Google Scholar] [CrossRef] [Green Version]
- Zambrano, N.; Bruni, P.; Minopoli, G.; Mosca, R.; Molino, D.; Russo, C.; Schettini, G.; Sudol, M.; Russo, T. The Beta-Amyloid Precursor Protein APP Is Tyrosine-Phosphorylated in Cells Expressing a Constitutively Active Form of the Abl Protoncogene. J. Biol. Chem. 2001, 276, 19787–19792. [Google Scholar] [CrossRef] [Green Version]
- Poulsen, E.T.; Iannuzzi, F.; Rasmussen, H.F.; Maier, T.J.; Enghild, J.J.; Jørgensen, A.L.; Matrone, C. An Aberrant Phosphorylation of Amyloid Precursor Protein Tyrosine Regulates Its Trafficking and the Binding to the Clathrin Endocytic Complex in Neural Stem Cells of Alzheimer’s Disease Patients. Front. Mol. Neurosci. 2017, 10, 59. [Google Scholar] [CrossRef] [Green Version]
- Rebelo, S.; Vieira, S.I.; Esselmann, H.; Wiltfang, J.; da Cruz e Silva, E.F.; da Cruz e Silva, O.A.B. Tyrosine 687 Phosphorylated Alzheimer’s Amyloid Precursor Protein Is Retained Intracellularly and Exhibits a Decreased Turnover Rate. Neurodegener. Dis. 2007, 4, 78–87. [Google Scholar] [CrossRef]
- Wang, H.; Li, R.; Shen, Y. β-Secretase: Its Biology as a Therapeutic Target in Diseases. Trends Pharmacol. Sci. 2013, 34, 215–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hampel, H.; Vassar, R.; De Strooper, B.; Hardy, J.; Willem, M.; Singh, N.; Zhou, J.; Yan, R.; Vanmechelen, E.; De Vos, A.; et al. The β-Secretase BACE1 in Alzheimer’s Disease. Biol. Psychiatry 2021, 89, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Pastorino, L.; Ikin, A.F.; Nairn, A.C.; Pursnani, A.; Buxbaum, J.D. The Carboxyl-Terminus of BACE Contains a Sorting Signal That Regulates BACE Trafficking but Not the Formation of Total Aβ. Mol. Cell. Neurosci. 2002, 19, 175–185. [Google Scholar] [CrossRef]
- Walter, J.; Fluhrer, R.; Hartung, B.; Willem, M.; Kaether, C.; Capell, A.; Lammich, S.; Multhaup, G.; Haass, C. Phosphorylation Regulates Intracellular Trafficking of Beta-Secretase. J. Biol. Chem. 2001, 276, 14634–14641. [Google Scholar] [CrossRef] [Green Version]
- von Arnim, C.A.F.; von Einem, B.; Weber, P.; Wagner, M.; Schwanzar, D.; Spoelgen, R.; Strauss, W.L.S.; Schneckenburger, H. Impact of Cholesterol Level upon APP and BACE Proximity and APP Cleavage. Biochem. Biophys. Res. Commun. 2008, 370, 207–212. [Google Scholar] [CrossRef]
- Radzimanowski, J.; Simon, B.; Sattler, M.; Beyreuther, K.; Sinning, I.; Wild, K. Structure of the Intracellular Domain of the Amyloid Precursor Protein in Complex with Fe65-PTB2. EMBO Rep. 2008, 9, 1134–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Evans, R.; O’Neill, M.; Pritzel, A.; Antropova, N.; Senior, A.; Green, T.; Žídek, A.; Bates, R.; Blackwell, S.; Yim, J.; et al. Protein Complex Prediction with AlphaFold-Multimer. BioRxiv 2021. [Google Scholar] [CrossRef]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making Protein Folding Accessible to All. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef]
- Das, B.; Yan, R. A Close Look at BACE1 Inhibitors for Alzheimer’s Disease Treatment. CNS Drugs 2019, 33, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Julian, L.; Olson, M.F. Rho-Associated Coiled-Coil Containing Kinases (ROCK): Structure, Regulation, and Functions. Small GTPases 2014, 5, e29846. [Google Scholar] [CrossRef]
- Kümper, S.; Mardakheh, F.K.; McCarthy, A.; Yeo, M.; Stamp, G.W.; Paul, A.; Worboys, J.; Sadok, A.; Jørgensen, C.; Guichard, S.; et al. Rho-Associated Kinase (ROCK) Function Is Essential for Cell Cycle Progression, Senescence and Tumorigenesis. Elife 2016, 5, e12994. [Google Scholar] [CrossRef] [PubMed]
- Cutler, C.; Lee, S.J.; Arai, S.; Rotta, M.; Zoghi, B.; Lazaryan, A.; Ramakrishnan, A.; DeFilipp, Z.; Salhotra, A.; Chai-Ho, W.; et al. Belumosudil for Chronic Graft-versus-Host Disease after 2 or More Prior Lines of Therapy: The ROCKstar Study. Blood 2021, 138, 2278–2289. [Google Scholar] [CrossRef]
- Brear, P.; Hyvönen, M. Crystal Structure of the Rho-Associated Coiled-Coil Kinase 2 Inhibitor Belumosudil Bound to CK2α. Acta Crystallogr. F Struct. Biol. Commun. 2022, 78, 348–353. [Google Scholar] [CrossRef]
- Akama, T.; Dong, C.; Virtucio, C.; Sullivan, D.; Zhou, Y.; Zhang, Y.-K.; Rock, F.; Freund, Y.; Liu, L.; Bu, W.; et al. Linking Phenotype to Kinase: Identification of a Novel Benzoxaborole Hinge-Binding Motif for Kinase Inhibition and Development of High-Potency Rho Kinase Inhibitors. J. Pharmacol. Exp. Ther. 2013, 347, 615–625. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, A.; Fukumoto, H.; Shah, T.; Whelan, C.M.; Irizarry, M.C.; Hyman, B.T. Demonstration by FRET of BACE Interaction with the Amyloid Precursor Protein at the Cell Surface and in Early Endosomes. J. Cell Sci. 2003, 116, 3339–3346. [Google Scholar] [CrossRef] [Green Version]
- Das, U.; Scott, D.A.; Ganguly, A.; Koo, E.H.; Tang, Y.; Roy, S. Activity-Induced Convergence of APP and BACE-1 in Acidic Microdomains via an Endocytosis-Dependent Pathway. Neuron 2013, 79, 447–460. [Google Scholar] [CrossRef] [Green Version]
- Somogyi, M.; Szimler, T.; Baksa, A.; Végh, B.M.; Bakos, T.; Paréj, K.; Ádám, C.; Zsigmond, Á.; Megyeri, M.; Flachner, B.; et al. A Versatile Modular Vector Set for Optimizing Protein Expression among Bacterial, Yeast, Insect and Mammalian Hosts. PLoS ONE 2019, 14, e0227110. [Google Scholar] [CrossRef] [Green Version]
- Maina, C.V.; Riggs, P.D.; Grandea, A.G.; Slatko, B.E.; Moran, L.S.; Tagliamonte, J.A.; McReynolds, L.A.; Guan, C.D. An Escherichia Coli Vector to Express and Purify Foreign Proteins by Fusion to and Separation from Maltose-Binding Protein. Gene 1988, 74, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Morrison, J.F. Kinetics of the Reversible Inhibition of Enzyme-Catalysed Reactions by Tight-Binding Inhibitors. Biochim. Biophys. Acta (BBA) Enzymol. 1969, 185, 269–286. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hajdú, I.; Végh, B.M.; Szilágyi, A.; Závodszky, P. Beta-Secretase 1 Recruits Amyloid-Beta Precursor Protein to ROCK2 Kinase, Resulting in Erroneous Phosphorylation and Beta-Amyloid Plaque Formation. Int. J. Mol. Sci. 2023, 24, 10416. https://doi.org/10.3390/ijms241310416
Hajdú I, Végh BM, Szilágyi A, Závodszky P. Beta-Secretase 1 Recruits Amyloid-Beta Precursor Protein to ROCK2 Kinase, Resulting in Erroneous Phosphorylation and Beta-Amyloid Plaque Formation. International Journal of Molecular Sciences. 2023; 24(13):10416. https://doi.org/10.3390/ijms241310416
Chicago/Turabian StyleHajdú, István, Barbara M. Végh, András Szilágyi, and Péter Závodszky. 2023. "Beta-Secretase 1 Recruits Amyloid-Beta Precursor Protein to ROCK2 Kinase, Resulting in Erroneous Phosphorylation and Beta-Amyloid Plaque Formation" International Journal of Molecular Sciences 24, no. 13: 10416. https://doi.org/10.3390/ijms241310416
APA StyleHajdú, I., Végh, B. M., Szilágyi, A., & Závodszky, P. (2023). Beta-Secretase 1 Recruits Amyloid-Beta Precursor Protein to ROCK2 Kinase, Resulting in Erroneous Phosphorylation and Beta-Amyloid Plaque Formation. International Journal of Molecular Sciences, 24(13), 10416. https://doi.org/10.3390/ijms241310416