
Proteome Landscape during Ripening of Solid Endosperm from Two Different Coconut Cultivars Reveals Contrasting Carbohydrate and Fatty Acid Metabolic Pathway Modulation

 ,
,  , , , , , ,
, , , , , ,  ,
, Abstract
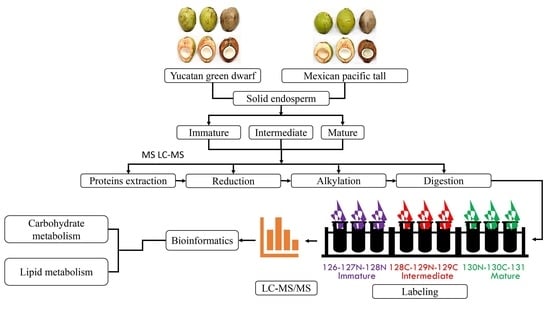
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Share and Cite
Félix, J.W.; Granados-Alegría, M.I.; Gómez-Tah, R.; Tzec-Simá, M.; Ruíz-May, E.; Canto-Canché, B.; Zamora-Briseño, J.A.; Bojórquez-Velázquez, E.; Oropeza-Salín, C.; Islas-Flores, I. Proteome Landscape during Ripening of Solid Endosperm from Two Different Coconut Cultivars Reveals Contrasting Carbohydrate and Fatty Acid Metabolic Pathway Modulation. Int. J. Mol. Sci. 2023, 24, 10431. https://doi.org/10.3390/ijms241310431
Félix JW, Granados-Alegría MI, Gómez-Tah R, Tzec-Simá M, Ruíz-May E, Canto-Canché B, Zamora-Briseño JA, Bojórquez-Velázquez E, Oropeza-Salín C, Islas-Flores I. Proteome Landscape during Ripening of Solid Endosperm from Two Different Coconut Cultivars Reveals Contrasting Carbohydrate and Fatty Acid Metabolic Pathway Modulation. International Journal of Molecular Sciences. 2023; 24(13):10431. https://doi.org/10.3390/ijms241310431
Chicago/Turabian StyleFélix, Jean Wildort, María Inés Granados-Alegría, Rufino Gómez-Tah, Miguel Tzec-Simá, Eliel Ruíz-May, Blondy Canto-Canché, Jesús Alejandro Zamora-Briseño, Esaú Bojórquez-Velázquez, Carlos Oropeza-Salín, and Ignacio Islas-Flores. 2023. "Proteome Landscape during Ripening of Solid Endosperm from Two Different Coconut Cultivars Reveals Contrasting Carbohydrate and Fatty Acid Metabolic Pathway Modulation" International Journal of Molecular Sciences 24, no. 13: 10431. https://doi.org/10.3390/ijms241310431
APA StyleFélix, J. W., Granados-Alegría, M. I., Gómez-Tah, R., Tzec-Simá, M., Ruíz-May, E., Canto-Canché, B., Zamora-Briseño, J. A., Bojórquez-Velázquez, E., Oropeza-Salín, C., & Islas-Flores, I. (2023). Proteome Landscape during Ripening of Solid Endosperm from Two Different Coconut Cultivars Reveals Contrasting Carbohydrate and Fatty Acid Metabolic Pathway Modulation. International Journal of Molecular Sciences, 24(13), 10431. https://doi.org/10.3390/ijms241310431