1. Introduction
Degeneration of the intervertebral disc (IVD) and the associated chronic lower back pain are serious health and economic problems. The costs of treating patients are very high. At the same time, there are no effective therapies to counteract degenerative changes and their effects. The disease is caused by disturbances in metabolic processes, which lead to structural disorders. The intervertebral disc consists of a gel-like nucleus pulpous (NP) and a surrounding fibrous ring (AF). The NP of a healthy adult disc is low in cells and rich in proteoglycans. As a result, the NP contains a large amount of water and thus fulfills its mechanical function. Resident cells are responsible for the biochemical composition of the NP through the production of proteoglycans, mainly aggrecan and collagen. IVD degeneration is characterized by an early decrease in cellularity of the NP region and associated extracellular matrix changes, reduced hydration, and progressive degeneration. As a result of the disease process, the NP matrix is destroyed due to increased expression of matrix metalloproteinases and inflammatory factors. This leads to disturbances in the production of NP matrices. This process gradually covers the AF and leads to its microdamage. All these changes are accompanied by disturbances in the function of cells found in the NP, especially progenitor cells. Progenitor cells are defined as tissue-specific stem cells responsible for regenerative processes. They have a limited self-renewal capacity but a high proliferative potential. Progenitor cells maintain homeostasis in the tissue in which they reside and play a major role in regeneration following injury. Notably, intervertebral discs are the largest avascular structures in the human body. This is certainly an additional factor determining the development of degenerative processes. The aim of this report is to analyze metabolic disorders in the degeneration process [
1,
2].
Nuclear magnetic resonance spectroscopy allows the study of biochemistry and cellular metabolism, not only in the whole organism in vivo (MRS) but also (at higher magnetic field strengths) in vitro (NMR). NMR methodologies have found wide application in biochemistry and medicine. In vitro NMR of cells, body fluids, and tissues is used in medical biochemistry to study pathophysiological processes, and recently, the technique has also been used to verify pathophysiological abnormalities in vivo. NMR is becoming a very important tool in precision medicine and metabolomics. The aim of this report is to present the results of in vitro NMR examinations characteristic of normal (in the T2-weighted image) and degenerate NPs.
In our research on the NP, we used NMR spectroscopy of extracts of hydrophilic and hydrophobic compounds of the tissue. To date, the NP has been tested in MAS NMR, but NP extracts have not been tested. There are no comparative data on NP extracts, hence the idea to determine if and what differences exist in aqueous and chloroform extracts in control and degenerate NPs. In addition, the identification of compounds that distinguish degenerate NPs from control may allow the in vivo spectroscopic measurements to be modified so that these compounds are visible. This will allow the translation from NMR to MRS disc studies and precise determination of degenerate disc localization.
2. Results
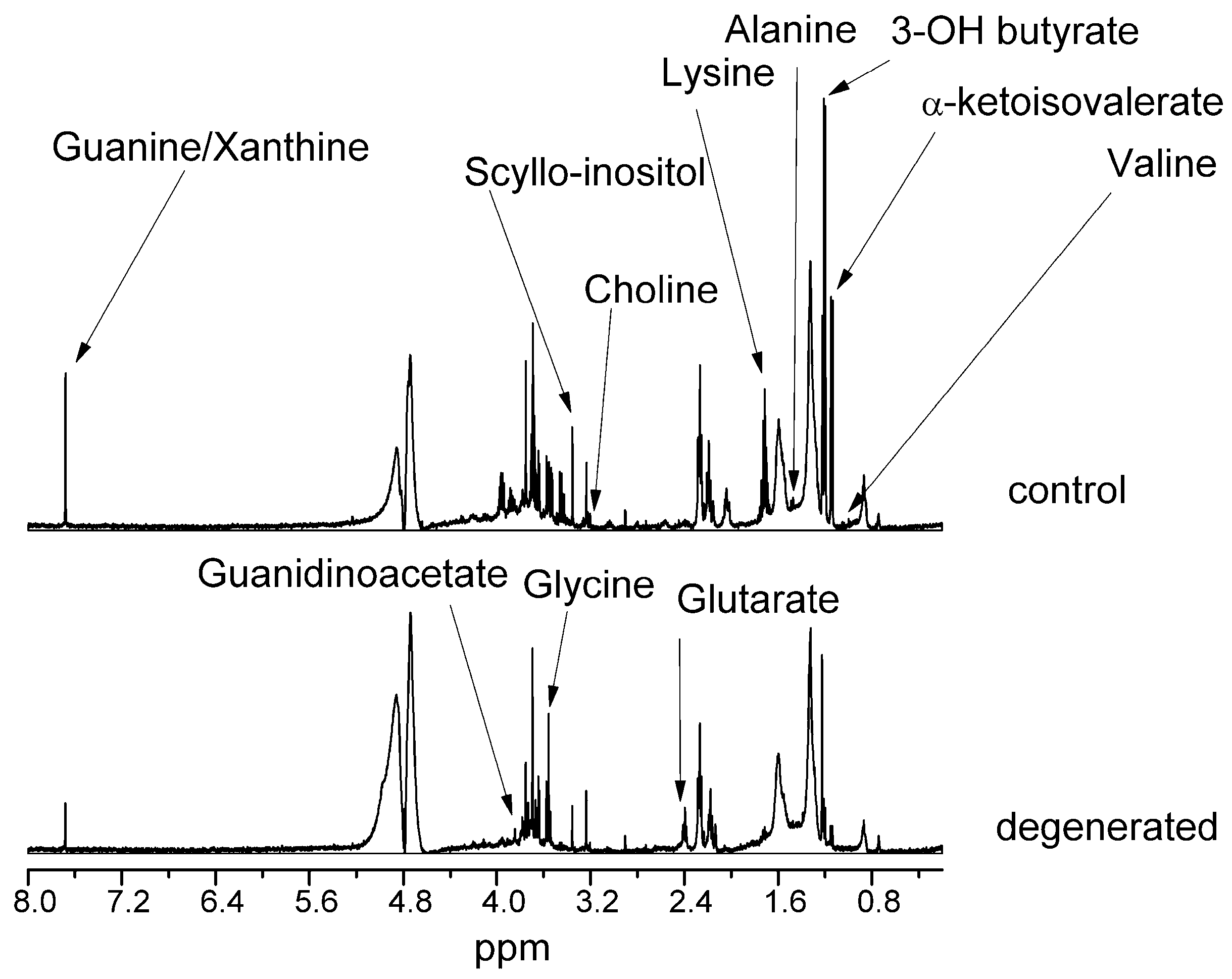
Figure 1 and
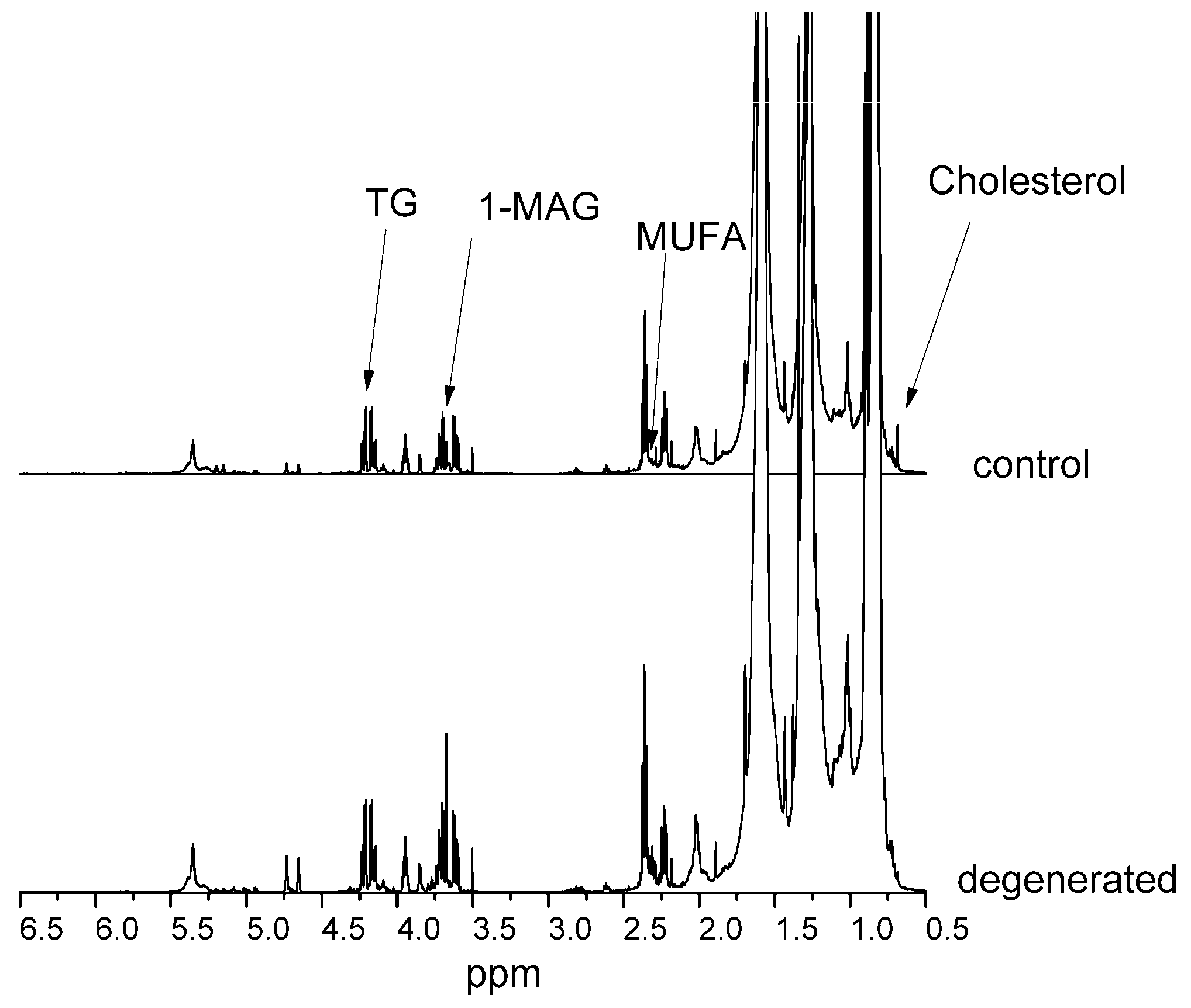
Figure 2 present NMR spectra for control and diseased NPs of hydrophilic and hydrophobic compounds, respectively. Spectra were scaled to a 1 mM TSP internal standard and a CHCl
3 signal for hydrophilic and hydrophobic compounds, respectively. Spectra were also normalized to the milligram of wet weight of the sample.
In
Table 1 and
Table 2, we present changes in selected compound signal intensities of the degenerated NP compared to the control NP.
The most changed compounds were valine, guanine/xanthine, 3-hydroxybutyric acid, α-ketoisovaleric acid, glycine, guanidinoacetate, scyllo-inositol, lysine, alanine, and glutaric acid. In the lipid fraction, almost all compounds increased their levels in dehydrated NP, except for free cholesterol, squalene, and fatty acids in all saturated polyunsaturated (PUFA) and monounsaturated (MUFA) forms. There was also a 65% decrease in the unassigned compound signal.
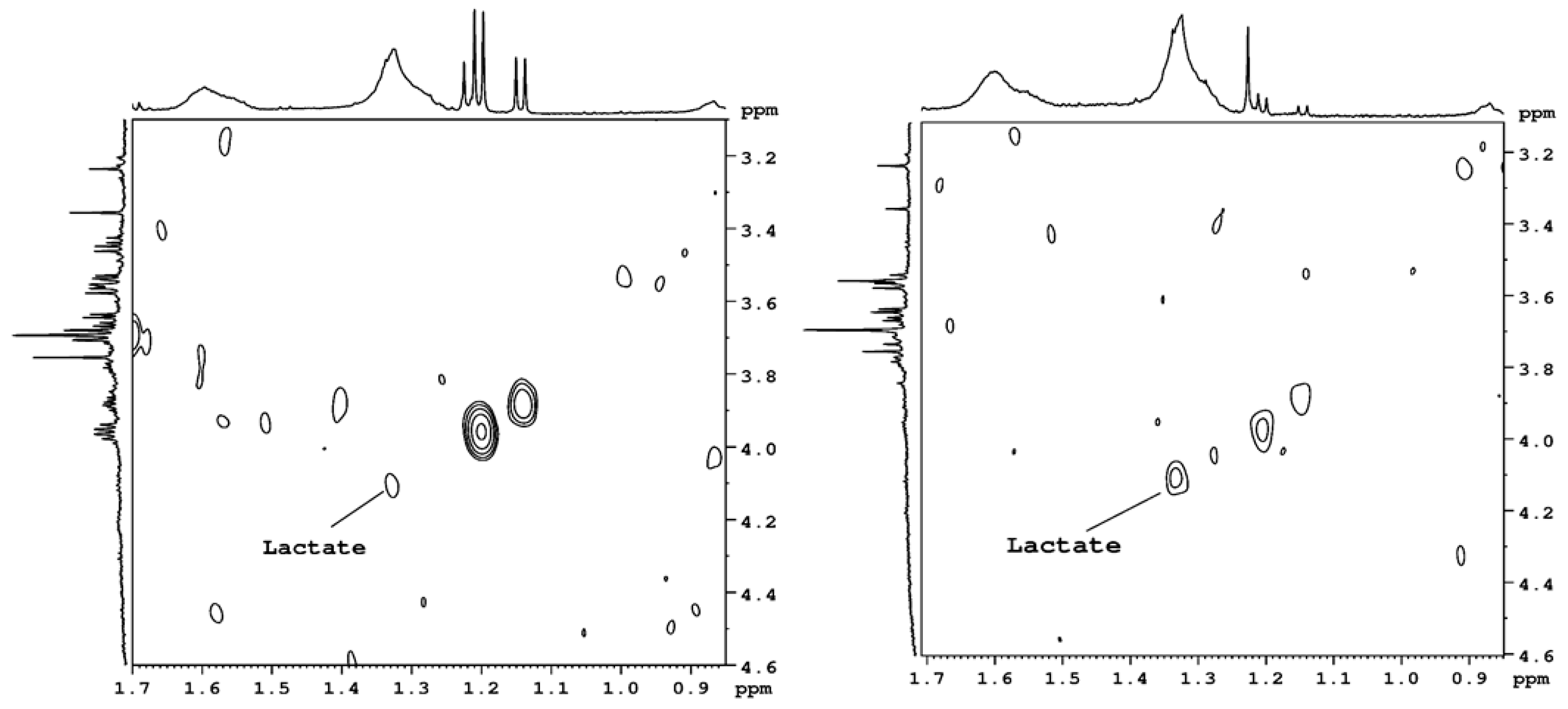
Figure 3 presents the 2D spectra measured in the homonuclear COSY sequence. The lactic acid cross-peak is more visible in the dehydrated NP than in the control NP. However, in the 1D NMR spectra, the level of lactic acid is small in the control and degenerated NP; therefore, it could not be measured with high accuracy.
3. Discussion
Disc degeneration can result from an imbalance between anabolic and catabolic processes or from a loss of balance in the state of metabolism that is maintained in a normal disc. The NMR spectroscopy method is sensitive enough to distinguish between control and diseased intervertebral discs based on metabolite studies. The fact that the study of both discs was performed in one patient allowed us to eliminate the influence of other factors, such as demographic, genetic, or environmental factors. In vitro spectroscopy, due to the high resolution of the spectrum, allowed us to identify compounds that indicate pathology. NMR spectroscopy can also be used in vivo, but the spectral resolution is much lower. On the basis of in vitro tests, it is possible to identify some of the indicated compounds in in vivo tests, e.g., glycine, by appropriately modifying the measurement sequences.
The results of our research collected in
Table 3 indicate disturbances in the biosynthesis and metabolism of cholesterol, the biosynthesis and degradation of various FA groups, ketone bodies, or lysine, and the metabolism of glycerophospholipids, purines, glycine, inositol, galactose, alanine, glutamate, and pyruvate in the biosynthesis of valine and isoleucine, leucine. All these disorders indicate pathomechanisms related to oxidative stress, energy, and neurotransmission disturbances and disturbances in the structure and functioning of cell membranes, inflammation, or chronic pain generators.
Mitochondria are a major site of reactive oxygen species (ROS) generation. Mitochondria-dependent ROS production has been reported in various disc cells derived from different species, including human NP and AF cells [
3]. Multiple studies have proven that extracellular uric acid at physiological concentrations is a strong antioxidant that can reduce the negative effects of oxidative stress by scavenging approximately 60% of the total-body ROS [
4]. Uric acid (UA) is the end product of purine metabolism. In our study, we could not measure UA because it is released outside of the cells, but we could measure earlier products from the purine metabolism cycle—guanine or xanthine. The levels of those compounds drastically decreased in degenerated discs to 30% of normal levels. IVD cells can generate ROS, and ROS overexpression has also been observed in degenerated IVDs. ROS are important mediators in the signal network of IVD cells. They regulate the matrix metabolism, pro-inflammatory phenotype, apoptosis, autophagy, and aging of IVD cells. The oxidative stress involved in ROS not only exacerbates extracellular matrix degradation but also promotes a reduction in the number of living cells in the IVD. When the serum UA level is lower, the antioxidant effect provided by UA is weakened, which may accelerate the progression of intervertebral disc degeneration [
5].
Squalene is a cholesterol precursor. In our research, we observed an increase in squalene concentration and a decrease in cholesterol in a damaged disc, which may indicate a defect in the enzymes mediating the conversion of squalene to cholesterol. Due to the pathway from squalene to cholesterol being multistage (KEGG metabolic pathways—Kyoto Encyclopedia of Genes and Genomes—databases), we cannot indicate the disturbance point because we did not see intermediate products due to low concentrations. Increased concentrations of monoacylglycerol (1-MAG), diglycerides, and triglycerides (TG) can lead to increased synthesis and subsequent degradation of various FA groups. Disturbances in FA degradation (KEGG) are also indicated by a 7-fold increased level of glutaric acid in a damaged disc. The accumulation of glutaric acid occurs in the case of glutaric acidosis type I (GA I; OMIM # 231670), which is a disorder of the systemic and brain metabolism of organic acids caused by biallelic variants of glutaryl-CoA dehydrogenase (GCDH), which encodes the mitochondrial, flavin-dependent GCDH that mediates the degradation of lysine, hydroxylysine, and tryptophan [
6]. A similarly high level of glutarate is found in type II glutaric acidosis (GA II) or multiple acyl-CoA dehydrogenase deficiency (MADD, OMIM # 231680), which is an inherited autosomal recessive disease affecting the metabolism of fatty acids, amino acids, and choline. In our research, with a high level of glutaric acid, we observed decreases in lysine (by approximately 90%) and choline (by approximately 50%). It has been observed that some people with GA I develop serious complications of the musculoskeletal system. In our research method, we were unable to identify a specific defective enzyme causing disturbances in the metabolism of lysine, choline, or FA, but perhaps the mechanism is similar to that of GA I and II [
6,
7].
Inflammation is another mechanism that can damage the intervertebral disc. In our research, an indication that this may be the case is increased levels of FA, MUFA, and PUFA. These compounds may be precursors of pro-inflammatory factors in NP cells. In NS cells, there is a balance between pro-inflammatory and anti-inflammatory factors that can be produced biochemically from arachidonic acid and PUFAs. If this state of equilibrium is disturbed, there may be an overproduction of inflammatory factors leading to damage to the intervertebral disc. The explanation of all details of biochemical pathways is in the publication of Das [
8].
A tissue or organ can generate pain only if it is innervated. In a healthy disc, nerve endings can be found only in the periphery of the outer annulus fibrosus (AF) and throughout the end plate with varying densities: no nerves are normally present in the inner AF or NP. These nerves are mainly nociceptive, responding to neurotrophins, such as nerve growth factor, and are implicated in disc pain transmission. However, in highly degenerated discs, these nerves can penetrate into the NP. All the tissues that surround the spinal motion segment are innervated and thus have the potential to cause pain [
1]. The increased level of glycine in the degenerate disc we observed is particularly important. The glycine receptor (GlyR) is the receptor for the neurotransmitter of the amino acid glycine. GlyR is an ionotropic receptor that acts through a chloride current. It is one of the most widespread inhibitory receptors in the central nervous system and plays an important role in a variety of physiological processes, especially mediating inhibitory neurotransmission in the spinal cord and brainstem. However, the GlyR subtypes contribute differently to nociception and chronic pain. The present evidence strongly suggests that post-translational modifications of synaptic α1β and α3β GlyR subunits are involved in spinal glycinergic disinhibition and chronic inflammatory pain [
9]. However, the roles of the α2 and β subunits remain largely unknown. A variety of molecular mechanisms suggest the highly dynamic plasticity of GlyR and other proteins in the glycinergic system. Signal pathways that control GlyR phosphorylation appear to be critical for the fine-tuning of inhibitory synaptic force. The data available thus far suggest that GlyR phosphorylation acts as a molecular switch regulating cell membrane expression, synaptic localization, and ion channel function. Whether these mechanisms work in parallel or independently at glycinergic synapses affected by pain signals remains to be determined. Despite the increasing number of recent reports, it should be noted that characterization of the roles of GlyR subtypes in nociception and chronic pain is far from complete, as many open questions in this area have yet to be resolved. A significant concern is the importance of tonic glycinergic and presynaptic GlyR inhibition in pain control. Another question is whether the supraspinal subtypes of GlyR or GlyR expressed in non-neuronal cells also participate in pain regulation, which is almost completely unexplored. Similarly, the expression and potential importance of GlyR on major afferents remain controversial. Finally, the role of GlyR-targeting autoantibodies in pain control in humans is largely unknown. The identification and characterization of novel protein kinases, protein‒protein interactions, and post-translational modifications involved in GlyR regulation could open exciting new avenues in pain research [
9,
10,
11].
In cell metabolism, intracellular oxygen levels are directly connected with changes in gene expression through the regulation of hypoxia-inducible factor (HIF) and its degradation by the von Hippel–Lindau axis. In the IVD, including the NP, these processes also occur, and proper HIF function is necessary for IVD integrity preservation. Dysregulation of HIF signaling is considered a trigger for degenerative cascade processes, and hypoxia-inducible factor 1α (HIF1α) is indicated as the guardian of hypoxic cells. In NP cells, HIF1α is regulated by both oxygen-dependent and oxygen-independent mechanisms involving prolyl hydroxylase domain-containing proteins (PHDs) and circadian rhythm. These cells have functional mitochondria that undergo HIF-dependent mitophagy and fragmentation. By adjusting cellular metabolism to living within low oxygen levels, the functioning of HIF1α maintains constant energy production levels, intracellular pH, and redox homeostasis and indirectly controls critical cellular processes, such as matrix biosynthesis. HIF1α influences glycolytic flux, in part, by maintaining H+/lactate efflux from NP cells at the final stage of glycolysis. In addition, it controls intracellular pH levels. High lactate levels act as a hypoxia mimetic factor by initiating the biosynthesis of TCA cycle intermediates. Studies in NP cells show a dynamic relationship between metabolic flux and HIF1α activity that is mediated by intracellular lactate levels, where intracellular lactate accumulation increases HIF1α activity [
12]. Our research indicates that the mechanism associated with HIV family proteins in the damaged intervertebral disc is disturbed. This is evidenced by lowered levels of lactate and ketone bodies.
Wang et al. [
13] proposed a new model of disc metabolism: lactate-dependent metabolic symbiosis. They proposed a working model of disc lactate-dependent metabolic symbiosis whereby hypoxic NP cells anaerobically convert glucose into lactate, which is then secreted and imported into neighboring cells of the more oxygenated AF tissue to be metabolized via oxidative phosphorylation. Their data suggest that lactate has been overlooked as an important biofuel in disc metabolism. Importantly, their findings also indicate the existence of lactate-dependent metabolic symbiosis between the NP and AF of the IVD as a metabolic adaptation to efficiently recycle lactate. Such metabolic adaptation would not only prevent the accumulation of lactate as a waste product but also simultaneously generate the much-needed energy as well as precursors for biosynthesis to cells residing within the nutrient-poor disc environment. One important implication of their model is that compromised or disrupted lactate symbiosis could contribute to metabolic disturbance and intervertebral disc degeneration. In our studies, the lactate level was decreased to 60% in degenerated discs compared to control discs. This may support the theory described by Wang et al. [
13].
Reduced disc height associated with disc degeneration can lead to facet overloading and subsequent facet degeneration and pain, which can be exacerbated by increased compression [
14]. Although discography has been used widely to localize painful discs and is regarded as the gold standard in the diagnosis of discogenic pain, this procedure is invasive and may even enhance the progression of disc degeneration itself. Unfortunately, none of the standard clinical tests can differentiate discogenic from nondiscogenic pain. For the past two decades, magnetic resonance imaging (MRI) has been the gold standard in assessing disc degeneration severity. In the T2-weighted MRI sequences, water content is correlated positively with signal intensity; therefore, the healthy NP is bright on T2 MRI because it contains a large amount of proteoglycan (PG) that attracts water. In disc degeneration, PG degradation results in a secondary reduction in water content, and consequently, the NP signal darkens on T2 MRI. Various novel MRI approaches have been introduced to detect subtle chemical changes in the disc matrix that may have important clinical value (e.g., T1rho, CEST, UTE, and sodium CSI). However, indications for use are currently not agreed on because prospective studies that validate the utility of these techniques, both in terms of LBP diagnosis and improved clinical outcomes, have not been extensively performed [
14].







{kind=link}
{kind=link}
{kind=link}
{kind=link}