
Harnessing Autophagy to Overcome Antigen-Specific T-Cell Dysfunction: Implication for People Living with HIV-1
 , , , and
, , , and {kind=link}
{kind=link}
Abstract
:1. Introduction: Autophagy, a Key Degradation Program for Nutrient Recycling
2. Thanks to Nutrient Diversification, Autophagy Becomes a New Facet of Ag-Specific T-Cell Metabolism
3. High Autophagy Is Key for Optimal Ag-Specific T-Cell Immunity in PLWH
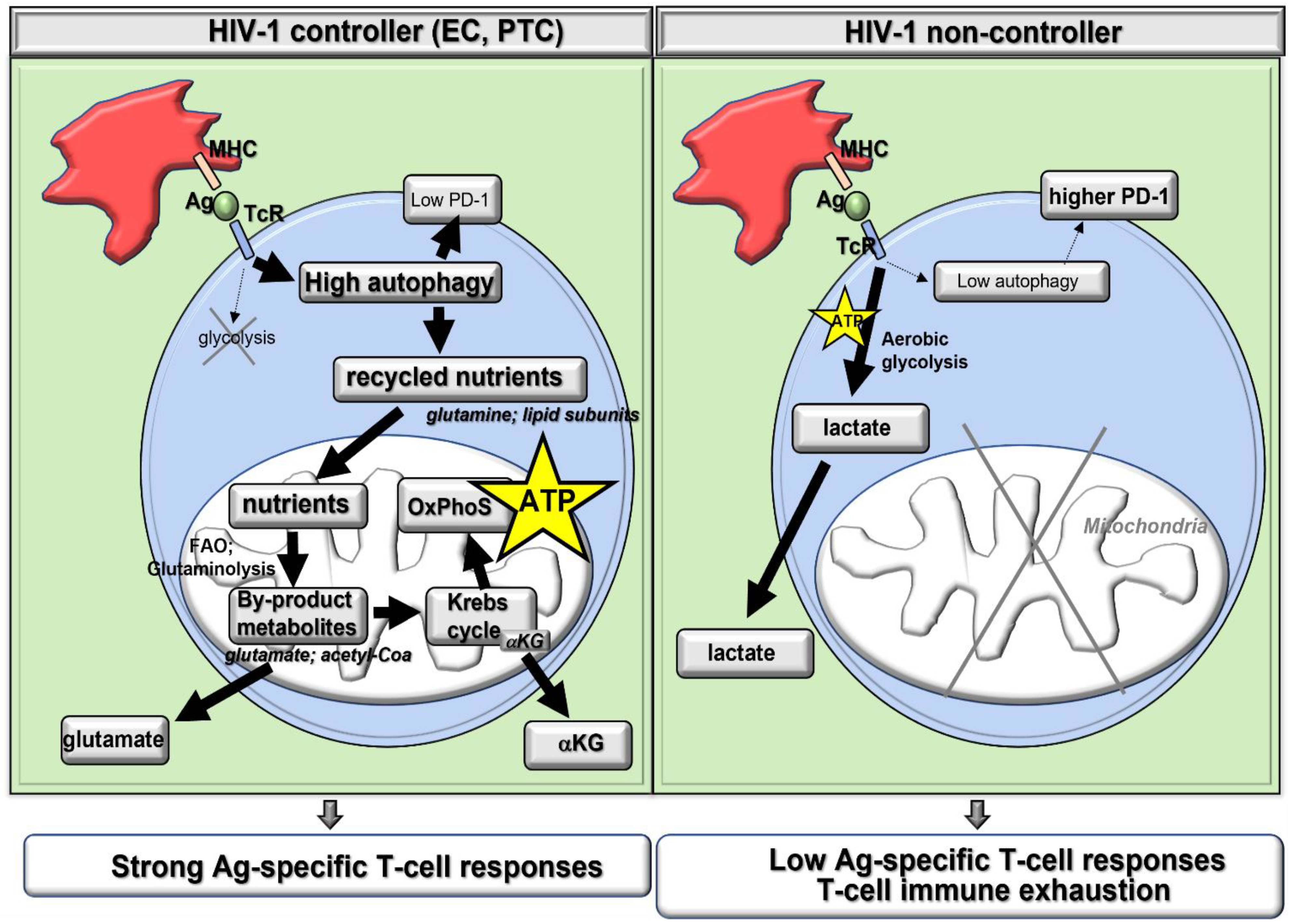
3.1. Autophagy Grants the Metabolic Plasticity-Dependent T-Cell Protection Found in Elite Controllers (EC)
3.2. Autophagy Improves Antiviral T-Cell Generation by Supporting Major Histocompatibility Complex (MHC) Restricted Ag Presentation
4. Autophagy in Ag-Specific T-Cells from PLWH Must Be Induced Together with ART (Figure 2)
4.1. As HIV-1 Proteins Hijack Autophagy to Block Lysosomal Degradation
4.2. As Autophagy-Mediated Metabolism Favors Cell Infectivity and Viral Replication
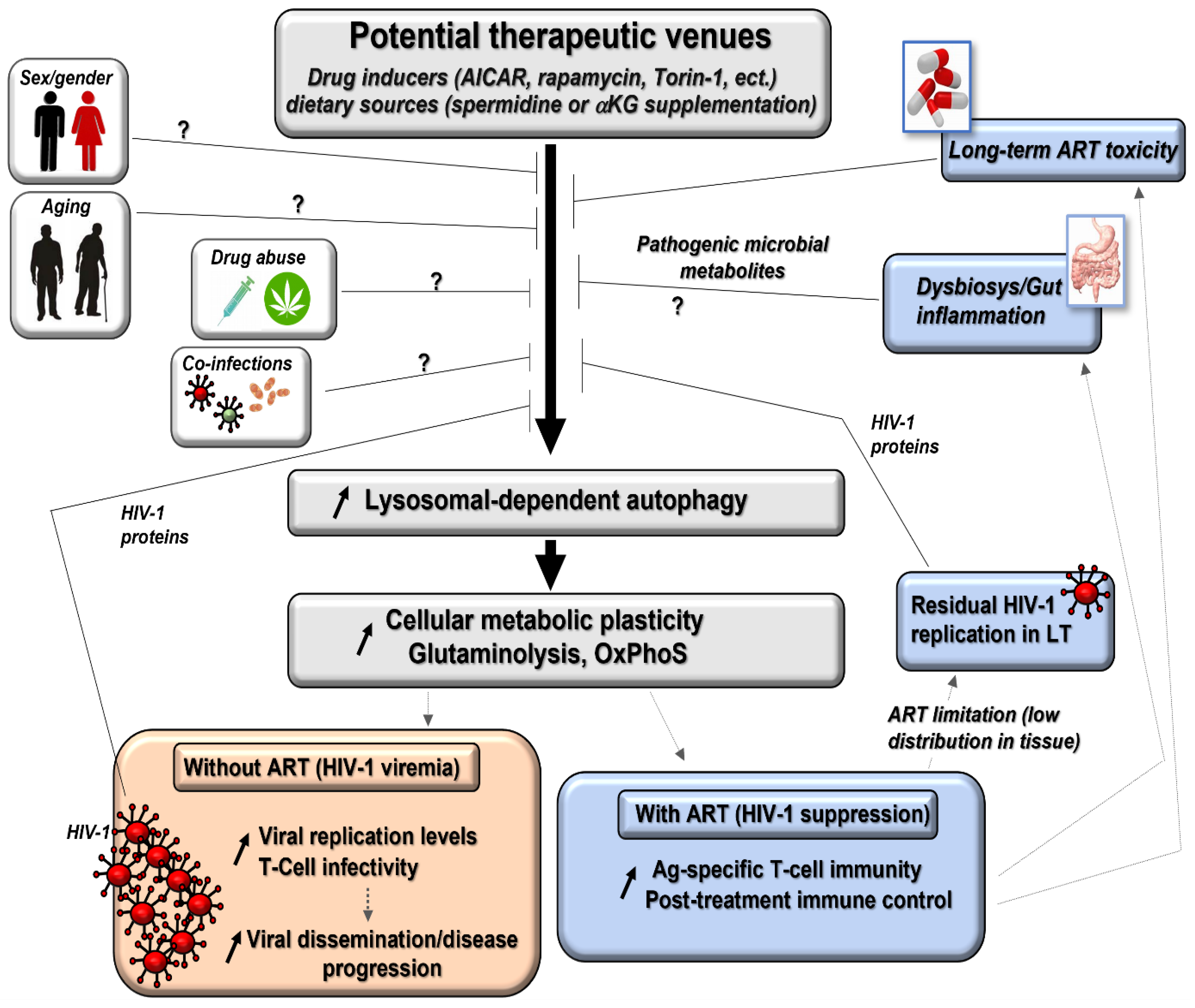
5. Clinical Parameters in PLWH May Impact the Efficacy of Autophagy Induction with ART Co-Treatment (Figure 2)
5.1. Biological Sex and Age of the Individuals
5.2. The Duration and Nature of Antiretroviral Drug (ARV) Regimens
5.3. Low Distribution of ARV into Lymphoid Tissues
5.4. Changes in the Gut Microbiota
6. Therapeutic Tools to Induce Autophagy in T-Cells of PLWH
6.1. Autophagy Activator Drugs
6.2. Pro-Autophagy Diets
7. Final Remarks
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Safaroghli-Azar, A.; Sanaei, M.-J.; Pourbagheri-Sigaroodi, A.; Bashash, D. Phosphoinositide 3-kinase (PI3K) classes: From cell signaling to endocytic recycling and autophagy. Eur. J. Pharmacol. 2023, 953, 175827. [Google Scholar] [CrossRef] [PubMed]
- Ren, A.; Gong, F.; Liu, G.; Fan, W. NR1H4-mediated circRHOBTB3 modulates the proliferation, metastasis, and Warburg effects of cervical cancer through interacting with IGF2BP3. Mol. Cell Biochem. 2023, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wan, K.; Shao, J.; Liu, X.; Cai, Y.; Xu, Y.; Li, L.; Xiong, L.; Liang, S. HOXD9 contributes to the Warburg effect and tumor metastasis in non-small cell lung cancer via transcriptional activation of PFKFB3. Exp. Cell Res. 2023, 427, 113583. [Google Scholar] [CrossRef]
- Xie, F.; Zhang, H.; Zhu, K.; Jiang, C.; Zhang, X.; Chang, H.; Qiao, Y.; Sun, M.; Wang, J.; Wang, M.; et al. PRMT5 promotes ovarian cancer growth through enhancing Warburg effect by methylating ENO1. Medcomm 2023, 4, e245. [Google Scholar] [CrossRef]
- Kim, J.; Kang, H.; Son, B.; Kim, M.-J.; Kang, J.; Park, K.H.; Jeon, J.; Jo, S.; Kim, H.Y.; Youn, H.; et al. NRBF2-mediated autophagy contributes to metabolite replenishment and radioresistance in glioblastoma. Exp. Mol. Med. 2022, 54, 1872–1885. [Google Scholar] [CrossRef]
- Kim, S.; Oh, M.; Kang, M.; Ko, J. Small leucine zipper protein functions as a modulator for metabolic reprogramming of colo-rectal cancer cells by inducing nutrient stress-mediated autophagy. Cell Mol. Life Sci. 2022, 79, 505. [Google Scholar] [CrossRef]
- Papadaki, S.; Magklara, A. Regulation of Metabolic Plasticity in Cancer Stem Cells and Implications in Cancer Therapy. Cancers 2022, 14, 5912. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, H.; Chen, K.; Guo, Y.; Mo, T.; Gao, T. Inhibition of autophagy impairs free fatty acid-induced excessive lipid accu-mulation in hepatocellular carcinoma and hepatic cells. J. Biosci. 2022, 47, 27. [Google Scholar] [CrossRef]
- Katheder, N.S.; Khezri, R.; O’farrell, F.; Schultz, S.W.; Jain, A.; Rahman, M.M.; Schink, K.O.; Theodossiou, T.A.; Johansen, T.; Juhász, G.; et al. Microenvironmental autophagy promotes tumour growth. Nature 2017, 541, 417–420. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Tautenhahn, H.-M.; Dirsch, O.; Dahmen, U. Blocking autophagy with chloroquine aggravates lipid accumulation and reduces intracellular energy synthesis in hepatocellular carcinoma cells, both contributing to its anti-proliferative effect. J. Cancer Res. Clin. Oncol. 2022, 148, 3243–3256. [Google Scholar] [CrossRef]
- Xu, G.; Jiang, Y.; Xiao, Y.; Liu, X.-D.; Yue, F.; Li, W.; Li, X.; He, Y.; Jiang, X.; Huang, H.; et al. Fast clearance of lipid droplets through MAP1S-activated autophagy suppresses clear cell renal cell carcinomas and promotes patient survival. Oncotarget 2015, 7, 6255–6265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Ding, J.; Li, B.; Sun, T.; You, X.; He, Q.; Sheng, W. Effects of icariin and curcumol on autophagy, ferroptosis, and lipid metabolism based on miR-7/m-TOR/SREBP1 pathway on prostate cancer. Biofactors 2023, 49, 438–456. [Google Scholar]
- Yin, H.; Shan, Y.; Xia, T.; Ji, Y.; Yuan, L.; You, Y.; You, B. Emerging Roles of Lipophagy in Cancer Metastasis. Cancers 2022, 14, 4526. [Google Scholar] [CrossRef] [PubMed]
- Botbol, Y.; Patel, B.; Macian, F. Common gamma-chain cytokine signaling is required for macroautophagy induction during CD4+ T-cell activation. Autophagy 2015, 11, 1864–1877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loucif, H.; Dagenais-Lussier, X.; Avizonis, D.; Choinière, L.; Beji, C.; Cassin, L.; Routy, J.-P.; Fritz, J.H.; Olagnier, D.; van Grevenynghe, J. Autophagy-dependent glutaminolysis drives superior IL21 production in HIV-1-specific CD4 T cells. Autophagy 2021, 18, 1256–1273. [Google Scholar] [CrossRef] [PubMed]
- Loucif, H.; Dagenais-Lussier, X.; Beji, C.; Cassin, L.; Jrade, H.; Tellitchenko, R.; Routy, J.-P.; Olagnier, D.; van Grevenynghe, J. Lipophagy confers a key metabolic advantage that ensures protective CD8A T-cell responses against HIV-1. Autophagy 2021, 17, 3408–3423. [Google Scholar] [CrossRef] [PubMed]
- Pua, H.H.; He, Y.W. Autophagy and lymphocyte homeostasis. Curr. Top. Microbiol. Immunol. 2009, 335, 85–105. [Google Scholar]
- Alsaleh, G.; Panse, I.; Swadling, L.; Zhang, H.; Richter, F.C.; Meyer, A.; Lord, J.; Barnes, E.; Klenerman, P.; Green, C.; et al. Autophagy in T cells from aged donors is maintained by spermidine and correlates with function and vaccine responses. eLife 2020, 9, e57950. [Google Scholar] [CrossRef]
- DeVorkin, L.; Pavey, N.; Carleton, G.; Comber, A.; Ho, C.; Lim, J.; McNamara, E.; Huang, H.; Kim, P.; Zacharias, L.G.; et al. Autophagy Regulation of Metabolism Is Required for CD8+ T Cell Anti-tumor Immunity. Cell Rep. 2019, 27, 502–513.e5. [Google Scholar] [CrossRef] [Green Version]
- Dowling, S.D.; Macian, F. Autophagy and T cell metabolism. Cancer Lett. 2018, 419, 20–26. [Google Scholar] [CrossRef]
- Mocholi, E.; Dowling, S.D.; Botbol, Y.; Gruber, R.C.; Ray, A.K.; Vastert, S.; Shafit-Zagardo, B.; Coffer, P.J.; Macian, F. Autophagy Is a Tolerance-Avoidance Mechanism that Modulates TCR-Mediated Signaling and Cell Metabolism to Prevent Induction of T Cell Anergy. Cell Rep. 2018, 24, 1136–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicoli, F.; Papagno, L.; Frere, J.J.; Cabral-Piccin, M.P.; Clave, E.; Gostick, E.; Toubert, A.; Price, D.A.; Caputo, A.; Appay, V. Naïve CD8+ T-Cells Engage a Versatile Metabolic Program upon Activation in Humans and Differ Energetically from Memory CD8+ T-Cells. Front. Immunol. 2018, 9, 2736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlie, K.; Westerback, A.; DeVorkin, L.; Hughson, L.R.; Brandon, J.M.; MacPherson, S.; Gadawski, I.; Townsend, K.N.; Poon, V.I.; Elrick, M.A.; et al. Survival of effector CD8+ T cells during influenza in-fection is dependent on autophagy. J. Immunol. 2015, 194, 4277–4286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdor, R.; Mocholi, E.; Botbol, Y.; Guerrero-Ros, I.; Chandra, D.; Koga, H.; Gravekamp, C.; Cuervo, A.M.; Macian, F. Chaper-one-mediated autophagy regulates T cell responses through targeted degradation of negative regulators of T cell activation. Nat. Immunol. 2014, 15, 1046–1054. [Google Scholar] [CrossRef] [Green Version]
- Jia, W.; He, M.X.; McLeod, I.X.; Guo, J.; Ji, D.; He, Y.W. Autophagy regulates T lymphocyte proliferation through selective deg-radation of the cell-cycle inhibitor CDKN1B/p27Kip1. Autophagy 2015, 11, 2335–2345. [Google Scholar] [CrossRef] [Green Version]
- Murera, D.; Arbogast, F.; Arnold, J.; Bouis, D.; Muller, S.; Gros, F. CD4 T cell autophagy is integral to memory maintenance. Sci. Rep. 2018, 8, 5951. [Google Scholar] [CrossRef] [Green Version]
- Puleston, D.J.; Zhang, H.; Powell, T.J.; Lipina, E.; Sims, S.; Panse, I.; Watson, A.S.; Cerundolo, V.; Townsend, A.R.; Klenerman, P.; et al. Autophagy is a critical regulator of memory CD8+ T cell formation. eLife 2014, 3, e03706. [Google Scholar] [CrossRef]
- Xu, X.; Araki, K.; Li, S.; Han, J.H.; Ye, L.; Tan, W.G.; Konieczny, B.T.; Bruinsma, M.W.; Martinez, J.; Pearce, E.L.; et al. Faculty Autophagy is essential for effector CD8(+) T cell survival and memory formation. Nat. Immunol. 2014, 15, 1152–1161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wik, J.A.; Skålhegg, B.S. T Cell Metabolism in Infection. Front. Immunol. 2022, 13, 840610. [Google Scholar] [CrossRef]
- Cillo, A.R.; Mellors, J.W. Which therapeutic strategy will achieve a cure for HIV-1? Curr. Opin. Virol. 2016, 18, 14–19. [Google Scholar] [CrossRef]
- de Gea-Grela, A.; Moreno, S. Controversies in the Design of Strategies for the Cure of HIV Infection. Pathogens 2023, 12, 322. [Google Scholar] [CrossRef]
- Capa, L.; Ayala-Suarez, R.; De La Torre Tarazona, H.E.; Gonzalez-Garcia, J.; Del Romero, J.; Alcami, J.; Diez-Fuertes, F. Elite con-trollers long-term non progressors present improved survival and slower disease progression. Sci. Rep. 2022, 12, 16356. [Google Scholar] [CrossRef] [PubMed]
- Casado, C.; Galvez, C.; Pernas, M.; Tarancon-Diez, L.; Rodriguez, C.; Sanchez-Merino, V.; Vera, M.; Olivares, I.; De Pablo-Bernal, R.; Merino-Mansilla, A.; et al. Permanent control of HIV-1 pathogenesis in exceptional elite controllers: A model of spontaneous cure. Sci. Rep. 2020, 10, 16356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loucif, H.; Gouard, S.; Dagenais-Lussier, X.; Murira, A.; Stager, S.; Tremblay, C.; Van Grevenynghe, J. Deciphering natural con-trol of HIV-1: A valuable strategy to achieve antiretroviral therapy termination. Cytokine Growth Factor Rev. 2018, 40, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Rutishauser, R.L.; Trautmann, L. CD8+ T-cell responses in HIV controllers: Potential implications for novel HIV remission strategies. Curr. Opin. HIV AIDS 2022, 17, 315–324. [Google Scholar] [CrossRef]
- Deeks, S.G.; International AIDS Society Towards a Cure Working Group; Lewin, S.R.; Ross, A.L.; Ananworanich, J.; Benkirane, M.; Cannon, P.; Chomont, N.; Douek, D.; Lifson, J.D.; et al. International AIDS Society global scientific strategy: Towards an HIV cure 2016. Nat. Med. 2016, 22, 839–850. [Google Scholar] [CrossRef]
- Blankson, J.N.; Turk, G.; Seiger, K.; Lian, X.; Sun, W.; Parsons, E.M.; Gao, C.; Rassadkina, Y.; Polo, M.L.; Czernikier, A.; et al. The Esperanza Patient: More Hope for a Sterilizing HIV-1 Cure. Ann. Intern. Med. 2022, 175, 135–136. [Google Scholar] [CrossRef]
- Duarte, R.F.; Salgado, M.; Sanchez-Ortega, I.; Arnan, M.; Canals, C.; Domingo-Domenech, E.; Fernandez-de-Sevilla, A.; Gonza-lez-Barca, E.; Moron-Lopez, S.; Nogues, N.; et al. CCR5 Delta32 homozygous cord blood allogeneic transplantation in a patient with HIV: A case report. Lancet HIV 2015, 2, e236–e242. [Google Scholar] [CrossRef]
- Angin, M.; Volant, S.; Passaes, C.; Lecuroux, C.; Monceaux, V.; Dillies, M.-A.; Valle-Casuso, J.C.; Pancino, G.; Vaslin, B.; Le Grand, R.; et al. Metabolic plasticity of HIV-specific CD8+ T cells is associated with enhanced antiviral potential and natural control of HIV-1 infection. Nat. Metab. 2019, 1, 704–716. [Google Scholar] [CrossRef]
- Chowdhury, F.Z.; Ouyang, Z.; Buzon, M.; Walker, B.D.; Lichterfeld, M.; Yu, X.G. Metabolic pathway activation distinguishes transcriptional signatures of CD8+ T cells from HIV-1 elite controllers. AIDS 2018, 32, 2669–2677. [Google Scholar] [CrossRef]
- Tarancon-Diez, L.; Rodríguez-Gallego, E.; Rull, A.; Peraire, J.; Viladés, C.; Portilla, I.; Jimenez-Leon, M.R.; Alba, V.; Herrero, P.; Leal, M.; et al. Immunometabolism is a key factor for the persistent spontaneous elite control of HIV-1 infection. Ebiomedicine 2019, 42, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Nardacci, R.; Amendola, A.; Ciccosanti, F.; Corazzari, M.; Esposito, V.; Vlassi, C.; Taibi, C.; Fimia, G.M.; Del Nonno, F.; Ippolito, G.; et al. Autophagy plays an important role in the containment of HIV-1 in nonprogressor-infected patients. Autophagy 2014, 10, 1167–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loucif, H.; Dagenais-Lussier, X.; Beji, C.; Telittchenko, R.; Routy, J.-P.; van Grevenynghe, J. Plasticity in T-cell mitochondrial metabolism: A necessary peacekeeper during the troubled times of persistent HIV-1 infection. Cytokine Growth Factor Rev. 2020, 55, 26–36. [Google Scholar] [CrossRef]
- Rolf, J.; Zarrouk, M.; Finlay, D.K.; Foretz, M.; Viollet, B.; Cantrell, D.A. AMPKalpha1: A glucose sensor that controls CD8 T-cell memory. Eur. J. Immunol. 2013, 43, 889–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciccosanti, F.; Corazzari, M.; Casetti, R.; Amendola, A.; Collalto, D.; Refolo, G.; Vergori, A.; Taibi, C.; D’offizi, G.; Antinori, A.; et al. High Levels of TRIM5α Are Associated with Xenophagy in HIV-1-Infected Long-Term Nonprogressors. Cells 2021, 10, 1207. [Google Scholar] [CrossRef]
- Sagnier, S.; Daussy, C.F.; Borel, S.; Robert-Hebmann, V.; Faure, M.; Blanchet, F.P.; Beaumelle, B.; Biard-Piechaczyk, M.; Espert, L. Autophagy Restricts HIV-1 Infection by Selectively Degrading Tat in CD4+T Lymphocytes. J. Virol. 2015, 89, 615–625. [Google Scholar] [CrossRef] [Green Version]
- Pedreño-López, S.; García, E.; Guerrero, D.; Gómez-Mora, E.; Mateu, L.M.; Pérez, F.O.; Senserrich, J.; Clotet, B.; Cabrera, C. Modulation of the autophagic pathway inhibits HIV-1 infection in human lymphoid tissue cultured ex vivo. Sci. Rep. 2022, 12, 7439. [Google Scholar] [CrossRef] [PubMed]
- Cloherty, A.P.M.; van Teijlingen, N.H.; Eisden, T.-J.T.H.D.; van Hamme, J.L.; Rader, A.G.; Geijtenbeek, T.B.H.; Schreurs, R.R.C.E.; Ribeiro, C.M.S. Autophagy-enhancing drugs limit mucosal HIV-1 acquisition and suppress viral replication ex vivo. Sci. Rep. 2021, 11, 4767. [Google Scholar] [CrossRef]
- Loi, M.; Müller, A.; Steinbach, K.; Niven, J.; da Silva, R.B.; Paul, P.; Ligeon, L.-A.; Caruso, A.; Albrecht, R.A.; Becker, A.C.; et al. Macroautophagy Proteins Control MHC Class I Levels on Dendritic Cells and Shape Anti-viral CD8 + T Cell Responses. Cell Rep. 2016, 15, 1076–1087. [Google Scholar] [CrossRef] [Green Version]
- Mintern, J.D.; Macri, C.; Chin, W.J.; Panozza, S.E.; Segura, E.; Patterson, N.L.; Zeller, P.; Bourges, D.; Bedoui, S.; McMillan, P.J.; et al. Differential use of autophagy by primary dendritic cells specialized in cross-presentation. Autophagy 2015, 11, 906–917. [Google Scholar] [CrossRef] [Green Version]
- Münz, C. Canonical and Non-Canonical Functions of the Autophagy Machinery in MHC Restricted Antigen Presentation. Front. Immunol. 2022, 13, 868888. [Google Scholar] [CrossRef]
- Paludan, C.; Schmid, D.; Landthaler, M.; Vockerodt, M.; Kube, D.; Tuschl, T.; Munz, C. Endogenous MHC class II processing of a viral nuclear antigen after autophagy. Science 2005, 307, 593–596. [Google Scholar] [CrossRef]
- Sengupta, S.; Zhang, J.; Reed, M.C.; Yu, J.; Kim, A.; Boronina, T.N.; Board, N.L.; Wrabl, J.O.; Shenderov, K.; Welsh, R.A.; et al. A cell-free antigen processing sys-tem informs HIV-1 epitope selection and vaccine design. J. Exp. Med. 2023, 220, e20221654. [Google Scholar] [CrossRef]
- Unanue, E.R. From antigen processing to peptide-MHC binding. Nat. Immunol. 2006, 7, 1277–1279. [Google Scholar] [CrossRef]
- Blanchet, F.P.; Moris, A.; Nikolic, D.S.; Lehmann, M.; Cardinaud, S.; Stalder, R.; Garcia, E.; Dinkins, C.; Leuba, F.; Wu, L.; et al. Human immunodeficiency virus-1 inhibition of immunoamphisomes in dendritic cells impairs early innate and adaptive immune responses. Immunity 2010, 32, 654–669. [Google Scholar] [CrossRef] [Green Version]
- Coulon, P.G.; Richetta, C.; Rouers, A.; Blanchet, F.P.; Urrutia, A.; Guerbois, M.; Piguet, V.; Theodorou, I.; Bet, A.; Schwartz, O.; et al. HIV-Infected Dendritic Cells Present Endogenous MHC Class II-Restricted Anti-gens to HIV-Specific CD4+ T Cells. J. Immunol. 2016, 197, 517–532. [Google Scholar] [CrossRef] [Green Version]
- Sarango, G.; Manoury, B.; Moris, A. TAX1BP1 a novel player in antigen presentation. Autophagy 2022, 19, 2153–2155. [Google Scholar] [CrossRef]
- Sarango, G.; Richetta, C.; Pereira, M.; Kumari, A.; Ghosh, M.; Bertrand, L.; Pionneau, C.; Le Gall, M.; Grégoire, S.; Jeger-Madiot, R.; et al. The Autophagy Receptor TAX1BP1 (T6BP) improves antigen presentation by MHC-II molecules. EMBO Rep. 2022, 23, e55470. [Google Scholar] [CrossRef]
- Jin, Y.; Sun, C.; Feng, L.; Li, P.; Xiao, L.; Ren, Y.; Wang, D.; Li, C.; Chen, L. Regulation of SIV antigen-specific CD4+ T cellular im-munity via autophagosome-mediated MHC II molecule-targeting antigen presentation in mice. PLoS ONE 2014, 9, e93143. [Google Scholar]
- Andersen, A.N.; Landsverk, O.J.; Simonsen, A.; Bogen, B.; Corthay, A.; Oynebraten, I. Coupling of HIV-1 Antigen to the Selec-tive Autophagy Receptor SQSTM1/p62 Promotes T-Cell-Mediated Immunity. Front. Immunol. 2017, 7, 167. [Google Scholar]
- Lanzavecchia, A.; Roosnek, E.; Gregory, T.; Berman, P.; Abrignani, S. T cells can present antigens such as HIV gp120 targeted to their own surface molecules. Nature 1988, 334, 530–532. [Google Scholar] [CrossRef]
- Addison, M.M.; Ellis, G.I.; Leslie, G.J.; Zawadzky, N.B.; Riley, J.L.; Hoxie, J.A.; Eisenlohr, L.C. HIV-1-Infected CD4(+) T Cells Present MHC Class II-Restricted Epitope via Endogenous Processing. J. Immunol. 2022, 209, 864–873. [Google Scholar] [CrossRef]
- Cao, L.; Glazyrin, A.; Kumar, S.; Kumar, A. Role of Autophagy in HIV Pathogenesis and Drug Abuse. Mol. Neurobiol. 2016, 54, 5855–5867. [Google Scholar] [CrossRef]
- Liu, Z.; Xiao, Y.; Torresilla, C.; Rassart, E.; Barbeau, B. Implication of Different HIV-1 Genes in the Modulation of Autophagy. Viruses 2017, 9, 389. [Google Scholar] [CrossRef] [Green Version]
- Nardacci, R.; Ciccosanti, F.; Marsella, C.; Ippolito, G.; Piacentini, M.; Fimia, G.M. Role of autophagy in HIV infection and path-ogenesis. J. Intern. Med. 2017, 281, 422–432. [Google Scholar] [CrossRef] [Green Version]
- Borel, S.; Robert-Hebmann, V.; Alfaisal, J.; Jain, A.; Faure, M.; Espert, L.; Chaloin, L.; Paillart, J.C.; Johansen, T.; Biard-Piechaczyk, M. HIV-1 viral infectivity factor interacts with microtubule-associated protein light chain 3 and inhibits autoph-agy. AIDS 2015, 29, 275–286. [Google Scholar] [CrossRef]
- Campbell, G.R.; Rawat, P.; Bruckman, R.S.; Spector, S.A. Human Immunodeficiency Virus Type 1 Nef Inhibits Autophagy through Transcription Factor EB Sequestration. PLoS Pathog. 2015, 11, e1005018. [Google Scholar] [CrossRef] [Green Version]
- Espert, L.; Varbanov, M.; Robert-Hebmann, V.; Sagnier, S.; Robbins, I.; Sanchez, F.; Lafont, V.; Biard-Piechaczyk, M. Differential Role of Autophagy in CD4 T Cells and Macrophages during X4 and R5 HIV-1 Infection. PLoS ONE 2009, 4, e5787. [Google Scholar] [CrossRef]
- Li, J.C.; Au, K.Y.; Fang, J.W.; Yim, H.C.; Chow, K.H.; Ho, P.L.; Lau, A.S. HIV-1 trans-activator protein dysregulates IFN-gamma signaling and contributes to the suppression of autophagy induction. AIDS 2011, 25, 15–25. [Google Scholar] [CrossRef]
- Sardo, L.; Iordanskiy, S.; Klase, Z.; Kashanchi, F. HIV-1 Nef blocks autophagy in human astrocytes. Cell Cycle 2015, 14, 3781–3782. [Google Scholar] [CrossRef]
- Van Grol, J.; Subauste, C.; Andrade, R.M.; Fujinaga, K.; Nelson, J.; Subauste, C.S. HIV-1 inhibits autophagy in bystander macro-phage/monocytic cells through Src-Akt and STAT3. PLoS ONE 2010, 5, e11733. [Google Scholar] [CrossRef] [Green Version]
- Sáez-Cirión, A.; Sereti, I. Immunometabolism and HIV-1 pathogenesis: Food for thought. Nat. Rev. Immunol. 2020, 21, 5–19. [Google Scholar] [CrossRef]
- Clerc, I.; Moussa, D.A.; Vahlas, Z.; Tardito, S.; Oburoglu, L.; Hope, T.J.; Sitbon, M.; Dardalhon, V.; Mongellaz, C.; Taylor, N. Entry of glucose- and glutamine-derived carbons into the citric acid cycle supports early steps of HIV-1 infection in CD4 T cells. Nat. Metab. 2019, 1, 717–730. [Google Scholar] [CrossRef]
- Guo, H.; Wang, Q.; Ghneim, K.; Wang, L.; Rampanelli, E.; Holley-Guthrie, E.; Cheng, L.; Garrido, C.; Margolis, D.M.; Eller, L.A.; et al. Multi-omics analyses reveal that HIV-1 alters CD4+ T cell immunometabolism to fuel virus replication. Nat. Immunol. 2021, 22, 423–433. [Google Scholar] [CrossRef]
- Cabrera-Rodríguez, R.; Hebmann, V.; Marfil, S.; Pernas, M.; Marrero-Hernández, S.; Cabrera, C.; Urrea, V.; Casado, C.; Olivares, I.; Márquez-Arce, D.; et al. HIV-1 envelope glycoproteins isolated from Viremic Non-Progressor individuals are fully functional and cytopathic. Sci. Rep. 2019, 9, 5544. [Google Scholar] [CrossRef] [Green Version]
- Cabrera-Rodríguez, R.; Pérez-Yanes, S.; Estévez-Herrera, J.; Márquez-Arce, D.; Cabrera, C.; Espert, L.; Blanco, J.; Valenzuela-Fernández, A. The Interplay of HIV and Autophagy in Early Infection. Front. Microbiol. 2021, 12, 661446. [Google Scholar] [CrossRef]
- Denizot, M.; Varbanov, M.; Espert, L.; Robert-Hebmann, V.; Sagnier, S.; Garcia, E.G.E.; Curriu, M.; Mamoun, R.; Blanco, J.; Biard-Piechaczyk, M. HIV-1 gp41 fusogenic function triggers autophagy in uninfected cells. Autophagy 2008, 4, 998–1008. [Google Scholar] [CrossRef] [Green Version]
- Mailler, E.; Waheed, A.A.; Park, S.-Y.; Gershlick, D.; Freed, E.O.; Bonifacino, J.S. The autophagy protein ATG9A promotes HIV-1 infectivity. Retrovirology 2019, 16, 18. [Google Scholar] [CrossRef] [Green Version]
- Lista, P.; Straface, E.; Brunelleschi, S.; Franconi, F.; Malorni, W. On the role of autophagy in human diseases: A gender perspec-tive. J. Cell Mol. Med. 2011, 15, 1443–1457. [Google Scholar] [CrossRef]
- Oliván, S.; Calvo, A.C.; Manzano, R.; Zaragoza, P.; Osta, R. Sex Differences in Constitutive Autophagy. BioMed Res. Int. 2014, 2014, 652817. [Google Scholar] [CrossRef] [Green Version]
- Shang, D.; Wang, L.; Klionsky, D.J.; Cheng, H.; Zhou, R. Sex differences in autophagy-mediated diseases: Toward precision medicine. Autophagy 2021, 17, 1065–1076. [Google Scholar] [CrossRef]
- Wang, T.; Zeng, F.; Li, X.; Wei, Y.; Wang, D.; Zhang, W.; Xie, H.; Wei, L.; Xiong, S.; Liu, C.; et al. Identification of key genes and pathways associated with sex differences in rheumatoid arthritis based on bioinformatics analysis. Clin. Rheumatol. 2023, 42, 399–406. [Google Scholar] [CrossRef]
- Sener, E.F.; Dana, H.; Tahtasakal, R.; Hamurcu, Z.; Taheri, S.; Delibasi, N.; Mehmetbeyoglu, E.; Sukranli, Z.Y.; Dal, F.; Tufan, E.; et al. Heterozygous Cc2d1a mice show sex-dependent changes in the Be-clin-1/p62 ratio with impaired prefrontal cortex and hippocampal autophagy. Prog. Neuropsychopharmacol. Biol. Psychiatry 2023, 125, 110764. [Google Scholar] [CrossRef]
- Triolo, M.; Oliveira, A.N.; Kumari, R.; Hood, D.A. The influence of age, sex, and exercise on autophagy, mitophagy, and lyso-some biogenesis in skeletal muscle. Skelet. Muscle 2022, 12, 13. [Google Scholar] [CrossRef]
- Lu, L.-F.; Jiang, J.-Y.; Du, W.-X.; Wang, X.-L.; Li, Z.-C.; Zhou, X.-Y.; Zhang, C.; Mou, C.-Y.; Chen, D.-D.; Li, Z.; et al. Fish female-biased gene cyp19a1a leads to female antiviral response attenuation between sexes by autophagic degradation of MITA. PLoS Pathog. 2022, 18, e1010626. [Google Scholar] [CrossRef]
- Fang, S.; Zhang, L.; Liu, Y.; Xu, W.; Wu, W.; Huang, Z.; Wang, X.; Liu, H.; Sun, Y.; Zhang, R.; et al. Lyso-some activation in peripheral blood mononuclear cells and prognostic significance of circulating LC3B in COVID-19. Brief Bio-Inform. 2021, 22, 1466–1475. [Google Scholar] [CrossRef]
- Campesi, I.; Occhioni, S.; Capobianco, G.; Fois, M.; Montella, A.; Dessole, S.; Franconi, F. Sex-specific pharmacological modula-tion of autophagic process in human umbilical artery smooth muscle cells. Pharmacol. Res. 2016, 113 Pt A, 166–174. [Google Scholar] [CrossRef]
- Addo, M.M.; Altfeld, M. Sex-Based Differences in HIV Type 1 Pathogenesis. J. Infect. Dis. 2014, 209 (Suppl. S3), S86–S92. [Google Scholar] [CrossRef] [Green Version]
- Schnittman, S.R.; Jung, W.; Fitch, K.V.; Zanni, M.V.; McCallum, S.; Lee, J.S.-L.; Shin, S.; Davis, B.J.; Fulda, E.S.; Diggs, M.R.; et al. Effect of host factors and COVID-19 infection on the humoral immune repertoire in treated HIV. JCI Insight 2023, 8, 166848. [Google Scholar] [CrossRef]
- Shakil, S.S.; Temu, T.M.; Kityo, C.; Nazzinda, R.; Erem, G.; Kentoffio, K.; Bittencourt, M.; Ntusi, N.A.; Zanni, M.V.; Longenecker, C.T. Sex modulates the association between inflammation and coronary atherosclerosis among older Ugandan adults with and without HIV. AIDS 2022, 37, 579–586. [Google Scholar] [CrossRef]
- Streeck, H.; Maestri, A.; Habermann, D.; Crowell, T.A.; Esber, A.L.; Son, G.; Eller, L.A.; Eller, M.A.; Parikh, A.P.; Horn, P.A.; et al. Dissecting drivers of immune activation in chronic HIV-1 infection. Ebiomedicine 2022, 83, 104182. [Google Scholar] [CrossRef]
- Tang, M.J.; Alexander, A.; Hoy, J.F. Gender and sex considerations in HIV and bone health. Curr. Opin. HIV AIDS 2023, 18, 75–80. [Google Scholar] [CrossRef]
- Vanpouille, C.; Wells, A.; Wilkin, T.; Mathad, J.S.; Morris, S.; Margolis, L.; Gianella, S. Sex differences in cytokine profiles during suppressive antiretroviral therapy. AIDS 2022, 36, 1215–1222. [Google Scholar] [CrossRef]
- Zanni, M.V.; Foldyna, B.; McCallum, S.; Burdo, T.H.; Looby, S.E.; Fitch, K.V.; Fulda, E.S.; Autissier, P.; Bloomfield, G.S.; Malvestutto, C.D.; et al. Sex Differences in Subclinical Atherosclerosis and Systemic Immune Activation/Inflammation among People With Human Immunodeficiency Virus in the United States. Clin. Infect. Dis. 2023, 76, 323–334. [Google Scholar] [CrossRef]
- Maskew, M.; Brennan, A.T.; Westreich, D.; McNamara, L.; MacPhail, A.P.; Fox, M.P. Gender Differences in Mortality and CD4 Count Response Among Virally Suppressed HIV-Positive Patients. J. Women’s Health 2013, 22, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Cheney, L.; Barbaro, J.M.; Berman, J.W. Antiretroviral Drugs Impact Autophagy with Toxic Outcomes. Cells 2021, 10, 909. [Google Scholar] [CrossRef]
- Bertrand, L.; Toborek, M. Dysregulation of Endoplasmic Reticulum Stress and Autophagic Responses by the Antiretroviral Drug Efavirenz. Mol. Pharmacol. 2015, 88, 304–315. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Stankov, M.V.; Hegermann, J.; Budida, R.; Panayotova-Dimitrova, D.; Schmidt, R.E.; Behrens, G.M.N. Zidovu-dine-Mediated Autophagy Inhibition Enhances Mitochondrial Toxicity in Muscle Cells. Antimicrob. Agents Chemother. 2019, 63, e01443-18. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Arroyo, O.; Gruevska, A.; Victor, V.M.; González-Polo, R.A.; Yakhine-Diop, S.M.; Fuentes, J.M.; Esplugues, J.V.; Blas-Garcia, A.; Apostolova, N. Mitophagy in human astrocytes treated with the antiretroviral drug Efavirenz: Lack of evidence or evidence of the lack. Antivir. Res. 2019, 168, 36–50. [Google Scholar] [CrossRef]
- Stankov, M.V.; Panayotova-Dimitrova, D.; Leverkus, M.; Schmidt, R.E.; Behrens, G.M. Thymidine analogues suppress autoph-agy and adipogenesis in cultured adipocytes. Antimicrob. Agents Chemother. 2013, 57, 543–551. [Google Scholar] [CrossRef] [Green Version]
- Stankov, M.V.; Panayotova-Dimitrova, D.; Leverkus, M.; Vondran, F.W.; Bauerfeind, R.; Binz, A.; Behrens, G.M. Autophagy inhibition due to thymidine analogues as novel mechanism leading to hepatocyte dysfunction and lipid accumulation. AIDS 2012, 26, 1995–2006. [Google Scholar] [CrossRef] [PubMed]
- Zha, B.S.; Wan, X.; Zhang, X.; Zha, W.; Zhou, J.; Wabitsch, M.; Wang, G.; Lyall, V.; Hylemon, P.B.; Zhou, H. HIV protease inhibi-tors disrupt lipid metabolism by activating endoplasmic reticulum stress and inhibiting autophagy activity in adipocytes. PLoS ONE 2013, 8, e59514. [Google Scholar] [CrossRef] [Green Version]
- Cheney, L.; Guzik, H.; Macaluso, F.P.; Macian, F.; Cuervo, A.M.; Berman, J.W. HIV Nef and Antiretroviral Therapy Have an Inhibitory Effect on Autophagy in Human Astrocytes that May Contribute to HIV-Associated Neurocognitive Disorders. Cells 2020, 9, 1426. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.; Thangaraj, A.; Chivero, E.T.; Periyasamy, P.; Burkovetskaya, M.E.; Niu, F.; Guo, M.-L.; Buch, S. N-Acetylcysteine Reverses Antiretroviral-Mediated Microglial Activation by Attenuating Autophagy-Lysosomal Dysfunction. Front. Neurol. 2020, 11, 840. [Google Scholar] [CrossRef]
- Tripathi, A.; Thangaraj, A.; Chivero, E.T.; Periyasamy, P.; Callen, S.; Burkovetskaya, M.E.; Guo, M.L.; Buch, S. Antiretrovi-ral-Mediated Microglial Activation Involves Dysregulated Autophagy and Lysosomal Dysfunction. Cells 2019, 8, 1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blagov, A.V.; Sukhorukov, V.N.; Guo, S.; Zhang, D.; Popov, M.A.; Orekhov, A.N. Impaired Mitochondrial Function in T-Lymphocytes as a Result of Exposure to HIV and ART. Cells 2023, 12, 1072. [Google Scholar] [CrossRef]
- Korencak, M.; Byrne, M.; Richter, E.; Schultz, B.T.; Juszczak, P.; Ake, J.A.; Ganesan, A.; Okulicz, J.F.; Robb, M.L.; Reyes, B.D.L.; et al. Effect of HIV infection and antiretroviral therapy on immune cellular functions. JCI Insight 2019, 4, 126675. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, L.; Ryom, L.; Neesgaard, B.; Wandeler, G.; Staub, T.; Gisinger, M.; Skoll, M.; Günthard, H.F.; Scherrer, A.; Mussini, C.; et al. Clinical Outcomes of 2-Drug Regimens vs 3-Drug Regimens in Antiretroviral Treatment–Experienced People Living with Human Immunodeficiency Virus. Clin. Infect. Dis. 2020, 73, e2323–e2333. [Google Scholar] [CrossRef]
- Maagaard, A.; Holberg-Petersen, M.; Lovgarden, G.; Holm, M.; Pettersen, F.O.; Kvale, D. Distinct mechanisms for mitochon-drial DNA loss in T and B lymphocytes from HIV-infected patients exposed to nucleoside reverse-transcriptase inhibitors and those naive to antiretroviral treatment. J. Infect. Dis. 2008, 198, 1474–1481. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.; Qin, C.; Feng, W.; Sheriston, C.; Khor, Y.J.; Medrano-Padial, C.; Watson, B.E.; Chan, T.; Ling, B.; Stocks, M.J.; et al. Oral administration of tipranavir with long-chain triglyceride results in moderate intestinal lymph targeting but no efficient delivery to HIV-1 reservoir in mesenteric lymph nodes. Int. J. Pharm. 2021, 602, 120621. [Google Scholar] [CrossRef]
- Fletcher, C.V.; Kroon, E.; Schacker, T.; Pinyakorn, S.; Chomont, N.; Chottanapund, S.; Prueksakaew, P.; Benjapornpong, K.; Buranapraditkun, S.; Phanuphak, N.; et al. Persistent HIV transcription and variable an-tiretroviral drug penetration in lymph nodes during plasma viral suppression. AIDS 2022, 36, 985–990. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.P.; Deleage, C.; White, N.; Sykes, C.; Brands, C.; Adamson, L.; Luciw, P.; Estes, J.D.; Kashuba, A.D.M. Antiretroviral drug exposure in lymph nodes is heterogeneous and drug dependent. J. Int. AIDS Soc. 2022, 25, 25895. [Google Scholar] [CrossRef] [PubMed]
- Rabezanahary, H.; Moukambi, F.; Palesch, D.; Clain, J.; Racine, G.; Andreani, G.; Benmadid-Laktout, G.; Zghidi-Abouzid, O.; Soundaramourty, C.; Tremblay, C.; et al. Despite early antiretroviral therapy effector memory and follicular helper CD4 T cells are major reservoirs in visceral lymphoid tissues of SIV-infected macaques. Mucosal Immunol. 2020, 13, 149–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqui, S.; Perez, S.; Gao, Y.; Doyle-Meyers, L.; Foley, B.T.; Li, Q.; Ling, B. Persistent Viral Reservoirs in Lymphoid Tissues in SIV-Infected Rhesus Macaques of Chinese-Origin on Suppressive Antiretroviral Therapy. Viruses 2019, 11, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.; Zhou, X.; Li, X.; Zhu, Q.; Peng, J.; Zhu, B.; Zheng, X.; Lu, Y.; Yang, D.; Wang, B.; et al. Depletion of Gut Microbiota Impairs Gut Barrier Function and Antiviral Immune Defense in the Liver. Front. Immunol. 2021, 12, 636803. [Google Scholar] [CrossRef]
- Annavajhala, M.K.; Khan, S.D.; Sullivan, S.B.; Shah, J.; Pass, L.; Kister, K.; Kunen, H.; Chiang, V.; Monnot, G.C.; Ricupero, C.L.; et al. Oral and Gut Microbial Diversity and Immune Regulation in Patients with HIV on Antiretroviral Therapy. Msphere 2020, 5, e798-19. [Google Scholar] [CrossRef] [Green Version]
- Ishizaka, A.; Koga, M.; Mizutani, T.; Lim, L.A.; Adachi, E.; Ikeuchi, K.; Ueda, R.; Aoyagi, H.; Tanaka, S.; Kiyono, H.; et al. Prolonged Gut Dysbiosis and Fecal Excretion of Hepatitis A Virus in Patients Infected with Human Immunodeficiency Virus. Viruses 2021, 13, 2101. [Google Scholar] [CrossRef]
- Ishizaka, A.; Koga, M.; Mizutani, T.; Parbie, P.K.; Prawisuda, D.; Yusa, N.; Sedohara, A.; Kikuchi, T.; Ikeuchi, K.; Adachi, E.; et al. Unique Gut Microbi-ome in HIV Patients on Antiretroviral Therapy (ART) Suggests Association with Chronic Inflammation. Microbiol. Spectr. 2021, 9, e0070821. [Google Scholar] [CrossRef]
- Mikaeloff, F.; Gelpi, M.; Benfeitas, R.; Knudsen, A.D.; Vestad, B.; Hogh, J.; Hov, J.R.; Benfield, T.; Murray, D.; Giske, C.G.; et al. Network-based multi-omics integration reveals metabolic at-risk profile within treated HIV-infection. eLife 2023, 12, e82785. [Google Scholar] [CrossRef]
- Wang, Z.; Qi, Q. Gut microbial metabolites associated with HIV infection. Futur. Virol. 2019, 14, 335–347. [Google Scholar] [CrossRef]
- Vesterbacka, J.; Rivera, J.; Noyan, K.; Parera, M.; Neogi, U.; Calle, M.; Paredes, R.; Sönnerborg, A.; Noguera-Julian, M.; Nowak, P. Richer gut microbiota with distinct metabolic profile in HIV infected Elite Controllers. Sci. Rep. 2017, 7, 6269. [Google Scholar] [CrossRef] [PubMed]
- Lapaquette, P.; Bizeau, J.-B.; Acar, N.; Bringer, M.-A. Reciprocal interactions between gut microbiota and autophagy. World J. Gastroenterol. 2021, 27, 8283–8301. [Google Scholar] [CrossRef] [PubMed]
- Nabavi-Rad, A.; Yadegar, A.; Sadeghi, A.; Aghdaei, H.A.; Zali, M.R.; Klionsky, D.J.; Yamaoka, Y. The interaction between autophagy, Helicobacter pylori, and gut microbiota in gastric carcinogenesis. Trends Microbiol. 2023, 27, S0966-842X(23)00113-0. [Google Scholar] [CrossRef]
- Planas, D.; Zhang, Y.; Monteiro, P.; Goulet, J.P.; Gosselin, A.; Grandvaux, N.; Hope, T.J.; Fassati, A.; Routy, J.P.; Ancuta, P. HIV-1 selectively targets gut-homing CCR6+CD4+ T cells via mTOR-dependent mechanisms. JCI Insight 2017, 2, e93230. [Google Scholar] [CrossRef] [Green Version]
- Mu, W.; Rezek, V.; Martin, H.; Carrillo, M.A.; Tomer, S.; Hamid, P.; Lizarraga, M.A.; Tibbe, T.D.; Yang, O.O.; Jamieson, B.D.; et al. Autophagy inducer rapamycin treatment reduces IFN-I–mediated Inflammation and improves anti–HIV-1 T cell response in vivo. JCI Insight 2022, 7, e159136. [Google Scholar] [CrossRef]
- Effros, R.B.; Walford, R.L.; Weindruch, R.; Mitcheltree, C. Influences of Dietary Restriction on Immunity to Influenza in Aged Mice. J. Gerontol. 2011, 46, B142–B147. [Google Scholar] [CrossRef]
- Qian, J.; Fang, Y.; Yuan, N.; Gao, X.; Lv, Y.; Zhao, C.; Zhang, S.; Li, Q.; Li, L.; Xu, L.; et al. Innate immune remodeling by short-term intensive fasting. Aging Cell 2021, 20, 13507. [Google Scholar] [CrossRef]
- Karagiannis, F.; Peukert, K.; Surace, L.; Michla, M.; Nikolka, F.; Fox, M.; Weiss, P.; Feuerborn, C.; Maier, P.; Schulz, S.; et al. Impaired ketogenesis ties metabolism to T cell dysfunction in COVID-19. Nature 2022, 609, 801–807. [Google Scholar] [CrossRef]
- Wang, Y.; Chi, H. Fasting as key tone for COVID immunity. Nat. Metab. 2022, 4, 1229–1231. [Google Scholar] [CrossRef] [PubMed]
- Hofer, S.J.; Kroemer, G.; Kepp, O. Autophagy-inducing nutritional interventions in experimental and clinical oncology. Int. Rev. Cell Mol. Biol. 2022, 373, 125–158. [Google Scholar] [PubMed]
- Hofer, S.J.; Simon, A.K.; Bergmann, M.; Eisenberg, T.; Kroemer, G.; Madeo, F. Mechanisms of spermidine-induced autophagy and geroprotection. Nat. Aging 2022, 2, 1112–1129. [Google Scholar] [CrossRef]
- Madeo, F.; Bauer, M.A.; Carmona-Gutierrez, D.; Kroemer, G. Spermidine: A physiological autophagy inducer acting as an anti-aging vitamin in humans? Autophagy 2019, 15, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, A.; De Wet, S.; Hofmeyr, J.-H.S.; Müller-Nedebock, K.K.; Loos, B. The Precision Control of Autophagic Flux and Vesicle Dynamics—A Micropattern Approach. Cells 2018, 7, 94. [Google Scholar] [CrossRef] [Green Version]
- Ito, D.; Ito, H.; Ando, T.; Sakai, Y.; Ideta, T.; Ishii, K.J.; Ishikawa, T.; Shimizu, M. Spermidine enhances the efficacy of adjuvant in HBV vaccination in mice. Hepatol. Commun. 2023, 7, e0104. [Google Scholar] [CrossRef] [PubMed]
- Laubert, M.; Bonifacius, A.; Dragon, A.C.; Mangare, C.; Blasczyk, R.; Huehn, J.; Eiz-Vesper, B. Enhancement of Antiviral T-Cell Responses by Vitamin C Suggests New Strategies to Improve Manufacturing of Virus-Specific T Cells for Adoptive Immuno-therapy. Biology 2022, 11, 536. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Wu, Y.; Zhang, Y.; Wang, Y.; Wu, H.; Zhang, T.; Chen, G.; Huang, X. Factors associated with post-treatment control of viral load in HIV-infected patients: A systematic review and meta-analysis. Int. J. Infect. Dis. 2023, 129, 216–227. [Google Scholar] [CrossRef] [PubMed]
- Giron, L.B.; Palmer, C.S.; Liu, Q.; Yin, X.; Papasavvas, E.; Sharaf, R.; Etemad, B.; Damra, M.; Goldman, A.R.; Tang, H.Y.; et al. Non-invasive plas-ma glycomic and metabolic biomarkers of post-treatment control of HIV. Nat. Commun. 2021, 12, 3922. [Google Scholar] [CrossRef]
- Churchill, M.J.; Deeks, S.G.; Margolis, D.M.; Siliciano, R.F.; Swanstrom, R. HIV reservoirs: What, where and how to target them. Nat. Rev. Microbiol. 2016, 14, 55–60. [Google Scholar] [CrossRef]
- Mutascio, S.; Mota, T.; Franchitti, L.; Sharma, A.A.; Willemse, A.; Bergstresser, S.N.; Wang, H.; Statzu, M.; Tharp, G.K.; Weiler, J.; et al. CD8(+) T cells promote HIV latency by remodeling CD4(+) T cell metabolism to enhance their survival, quiescence, and stemness. Immunity 2023, 56, 1132–1147.e6. [Google Scholar] [CrossRef]
- Valle-Casuso, J.C.; Angin, M.; Volant, S.; Passaes, C.; Monceaux, V.; Mikhailova, A.; Bourdic, K.; Avettand-Fenoel, V.; Boufassa, F.; Sitbon, M.; et al. Cellular Metabolism Is a Major Determinant of HIV-1 Reservoir Seeding in CD4+ T Cells and Offers an Opportunity to Tackle Infection. Cell Metab. 2019, 29, 611–626.e5. [Google Scholar] [CrossRef] [Green Version]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghahari, N.; Telittchenko, R.; Loucif, H.; Isnard, S.; Routy, J.-P.; Olagnier, D.; van Grevenynghe, J. Harnessing Autophagy to Overcome Antigen-Specific T-Cell Dysfunction: Implication for People Living with HIV-1. Int. J. Mol. Sci. 2023, 24, 11018. https://doi.org/10.3390/ijms241311018
Ghahari N, Telittchenko R, Loucif H, Isnard S, Routy J-P, Olagnier D, van Grevenynghe J. Harnessing Autophagy to Overcome Antigen-Specific T-Cell Dysfunction: Implication for People Living with HIV-1. International Journal of Molecular Sciences. 2023; 24(13):11018. https://doi.org/10.3390/ijms241311018
Chicago/Turabian StyleGhahari, Nazanin, Roman Telittchenko, Hamza Loucif, Stephane Isnard, Jean-Pierre Routy, David Olagnier, and Julien van Grevenynghe. 2023. "Harnessing Autophagy to Overcome Antigen-Specific T-Cell Dysfunction: Implication for People Living with HIV-1" International Journal of Molecular Sciences 24, no. 13: 11018. https://doi.org/10.3390/ijms241311018