
The Influence of Nicotine on Trophoblast-Derived Exosomes in a Mouse Model of Pathogenic Preeclampsia

Abstract
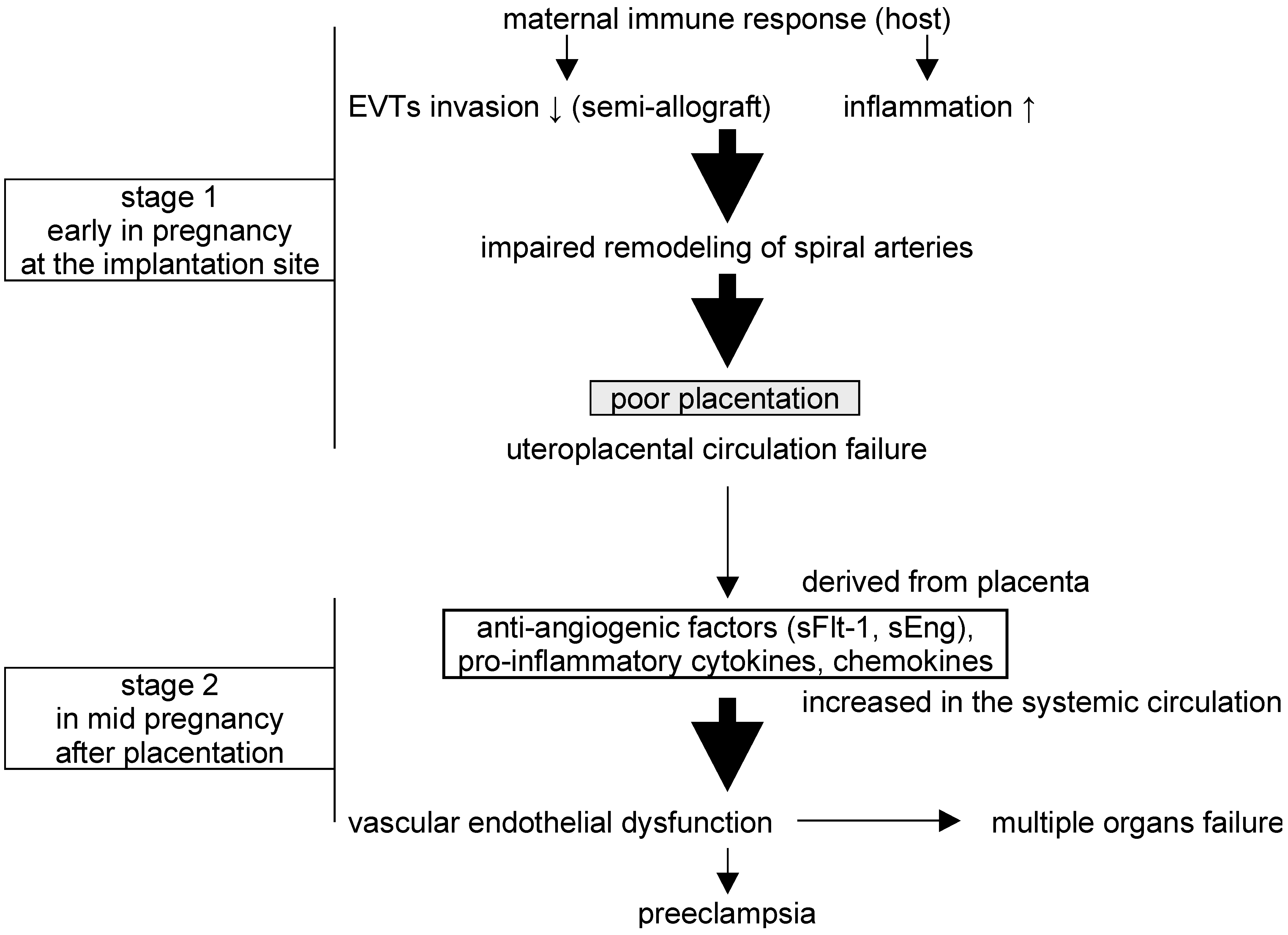
:1. Introduction
2. Results
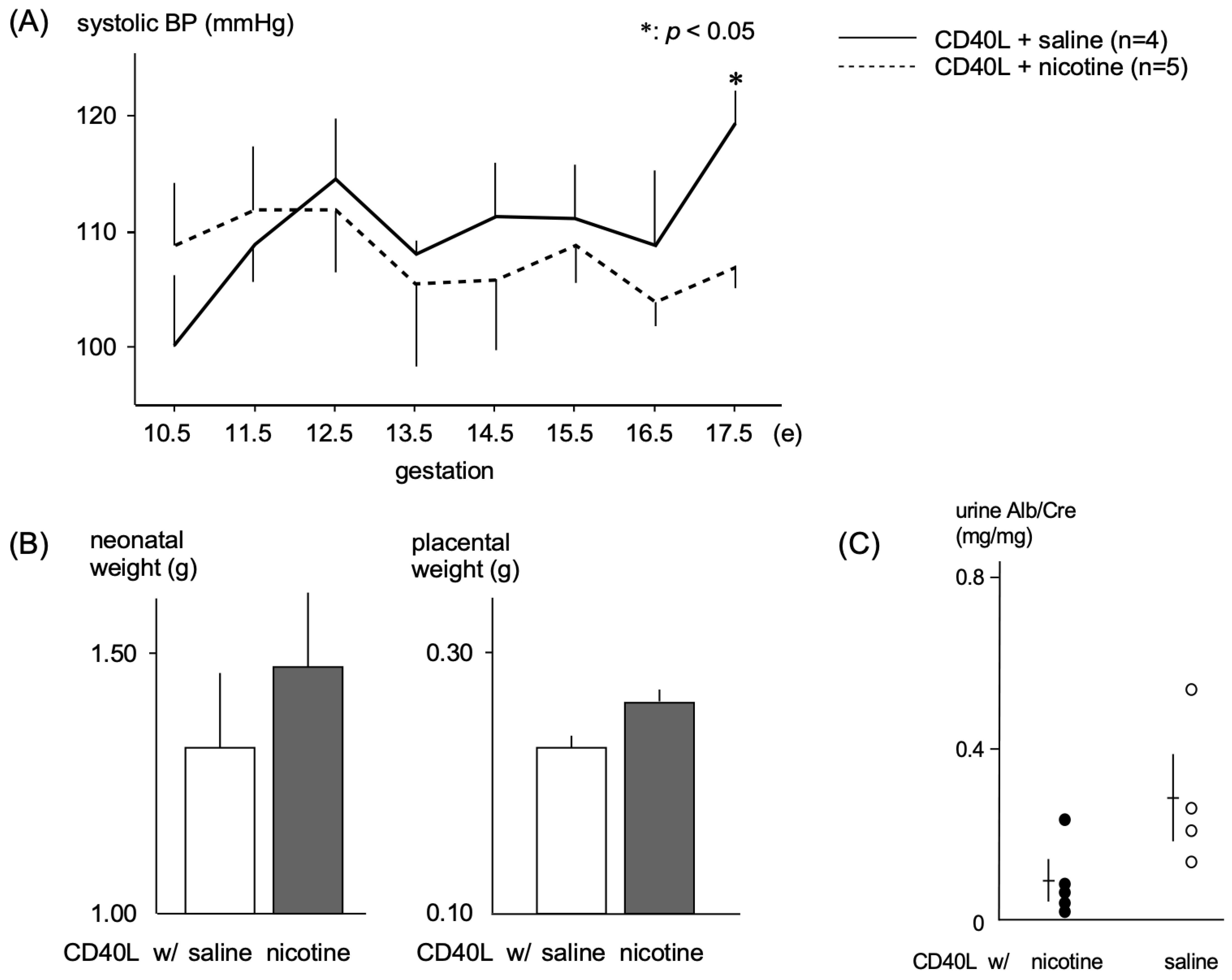
2.1. Effects of Nicotine in a PE Mouse Model
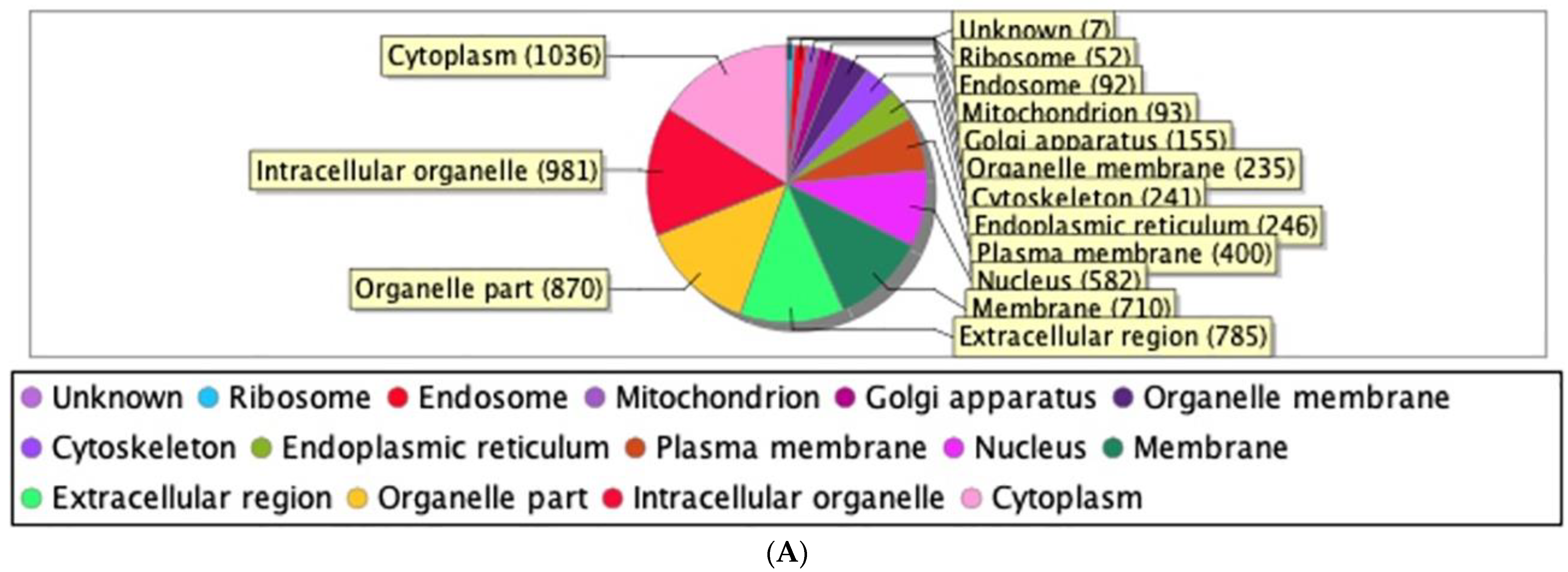
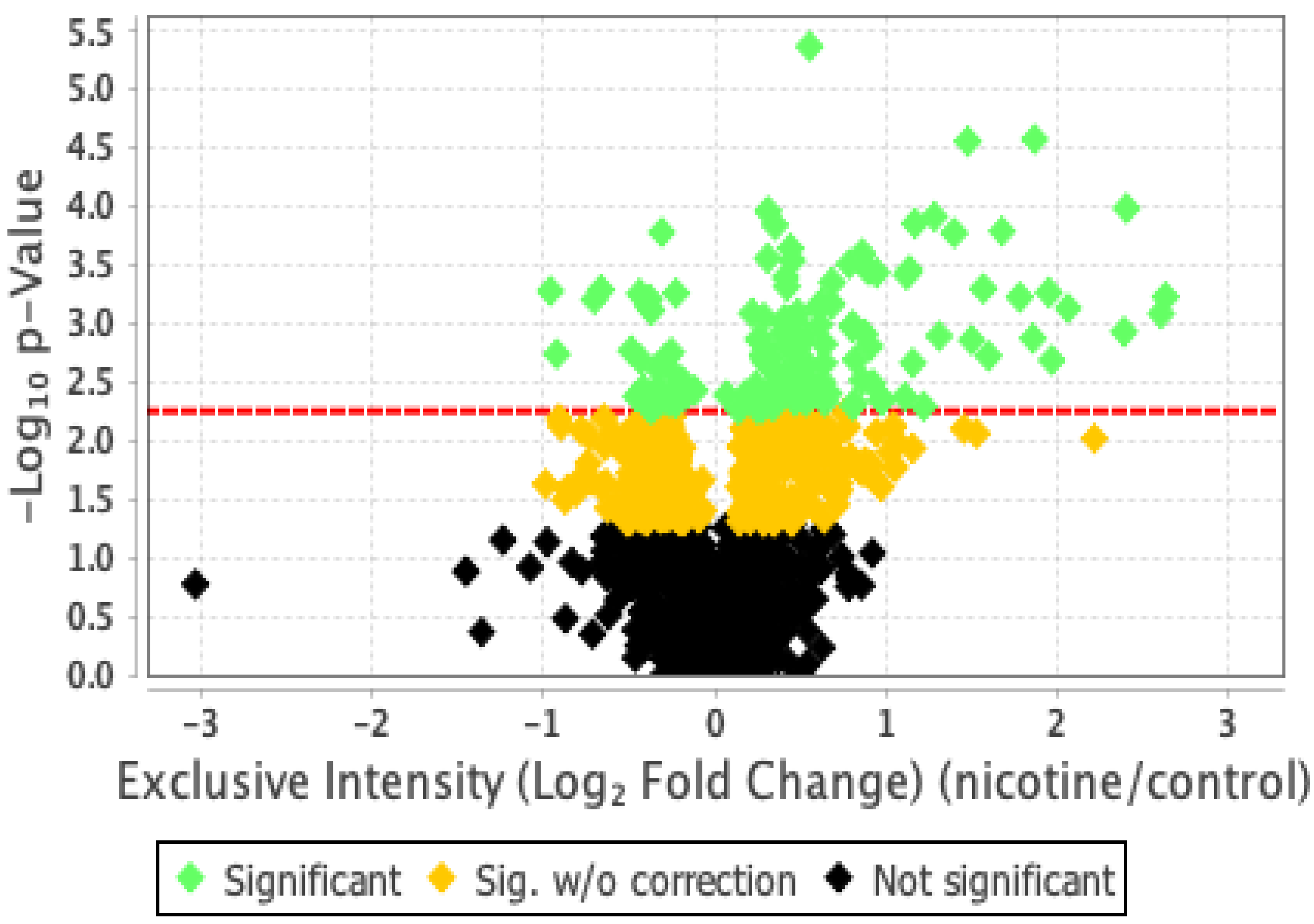
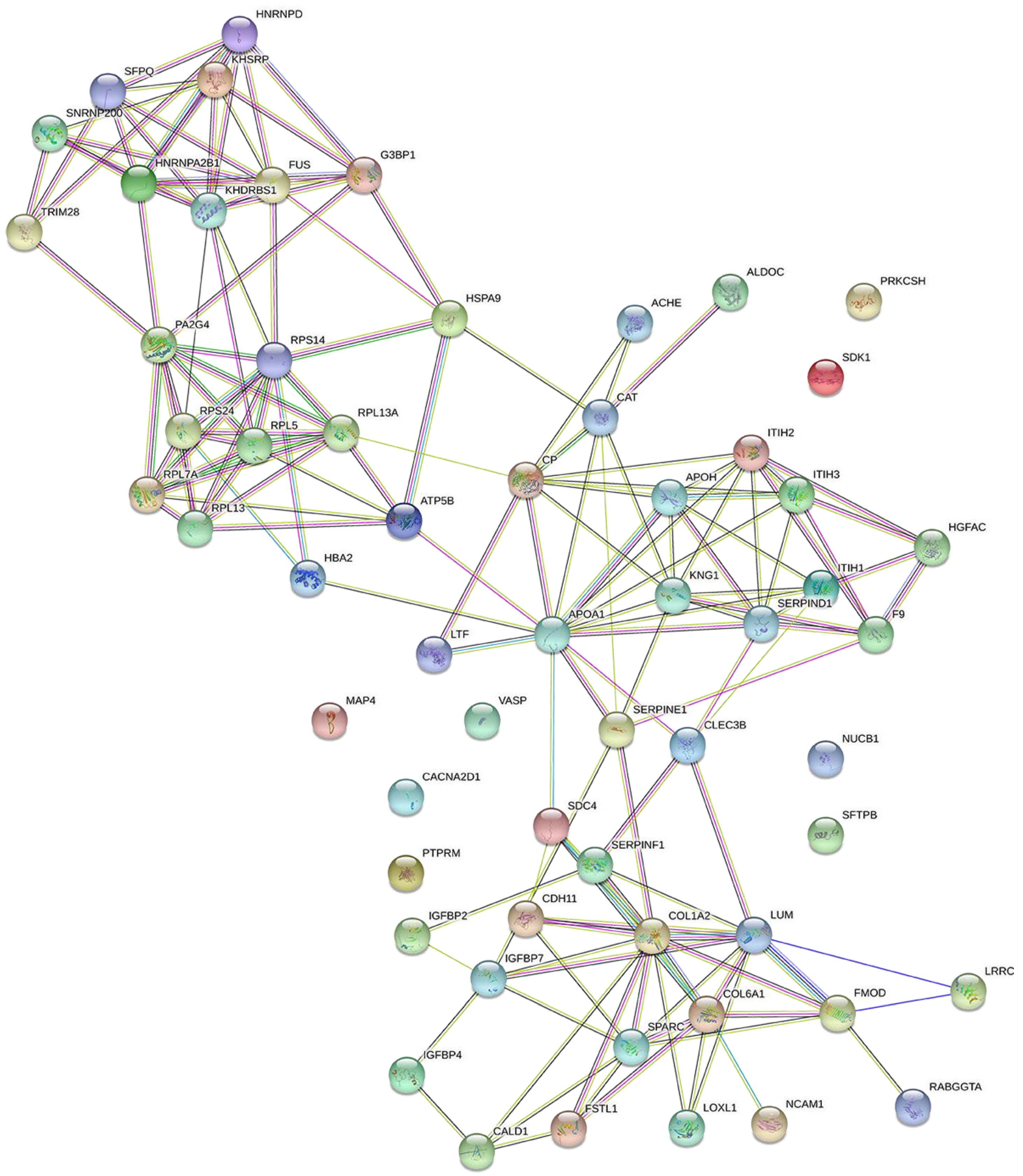
2.2. Proteomic Analysis and Bioinformatic Characterization
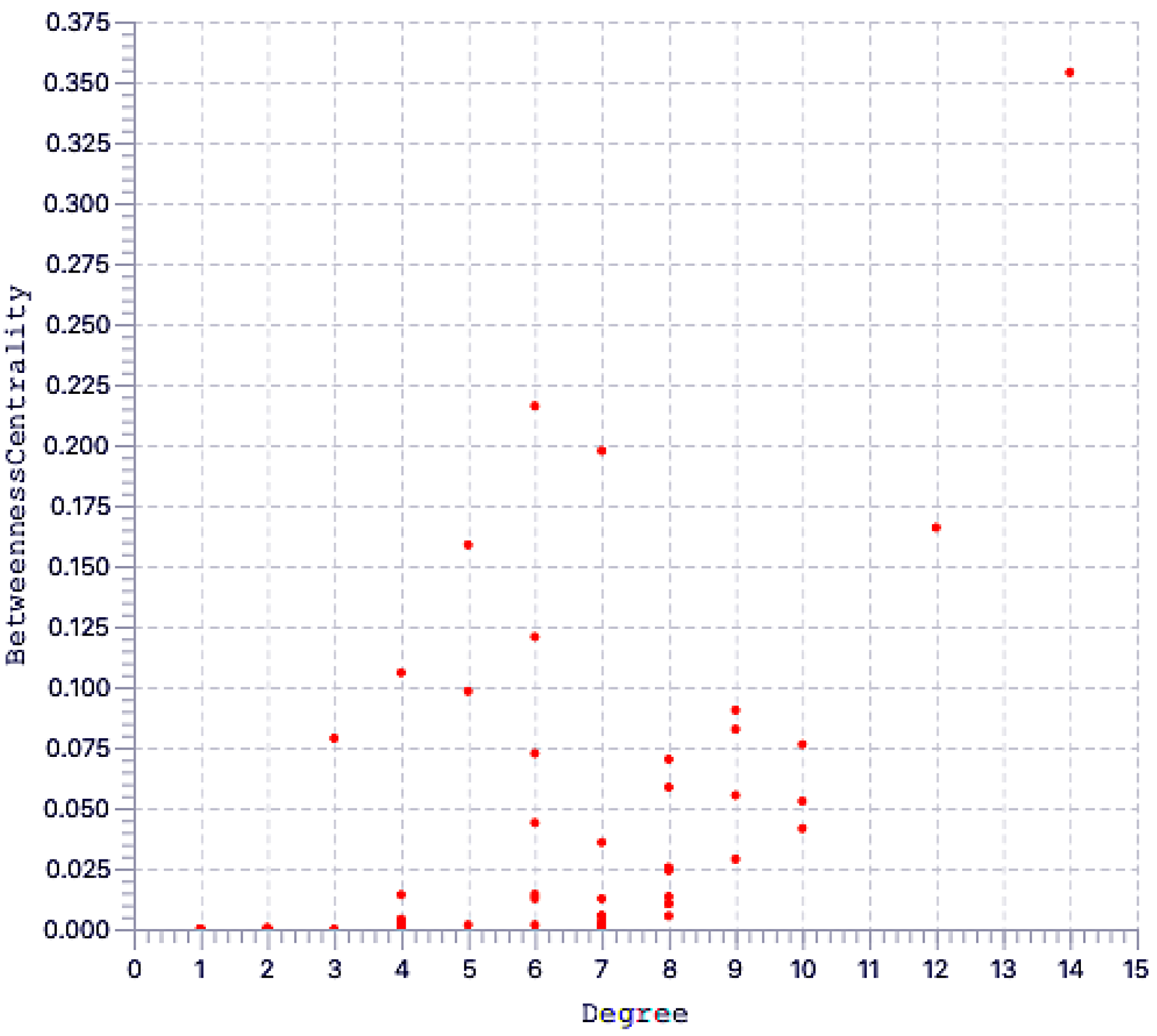
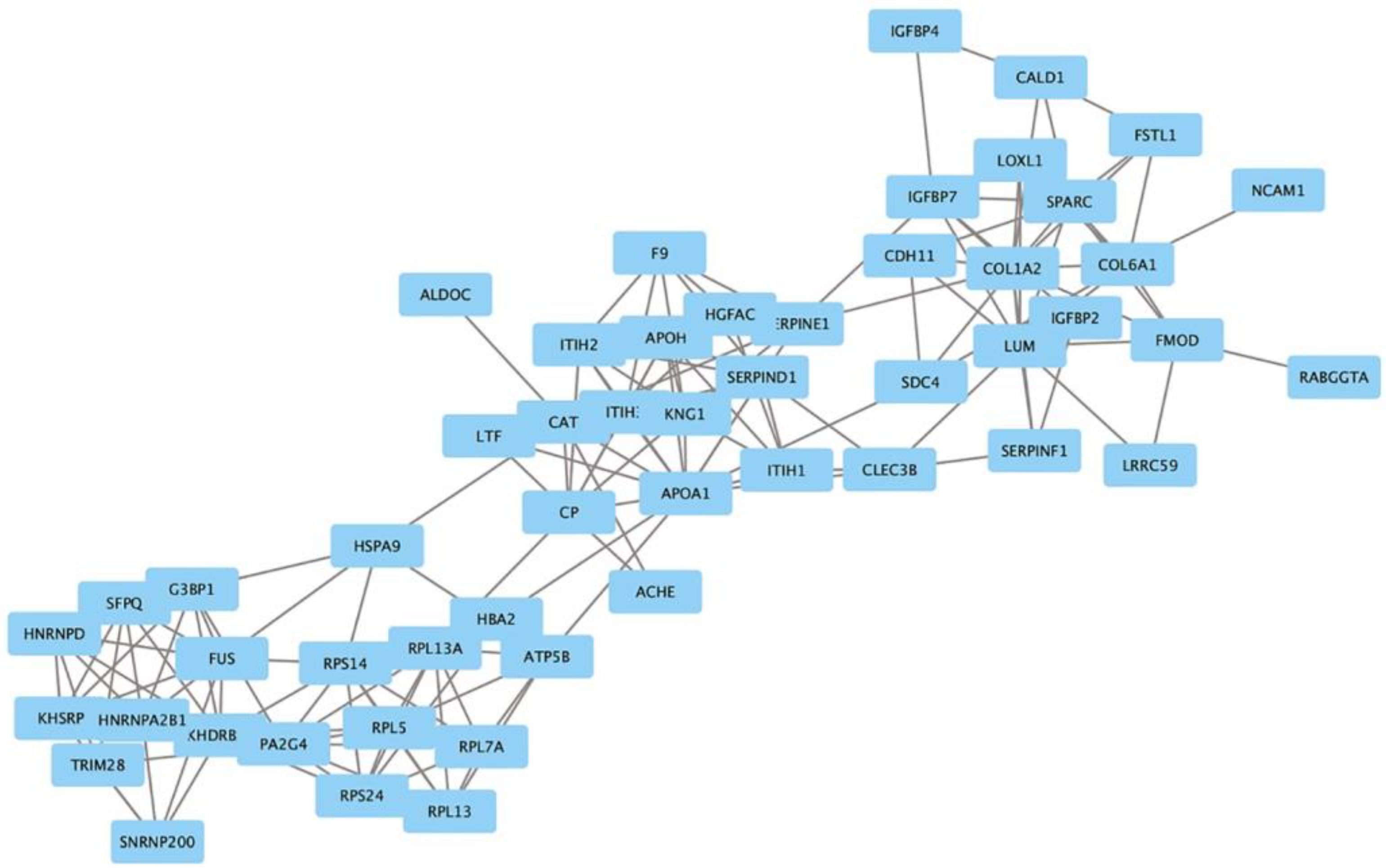
2.3. Protein Network Analysis
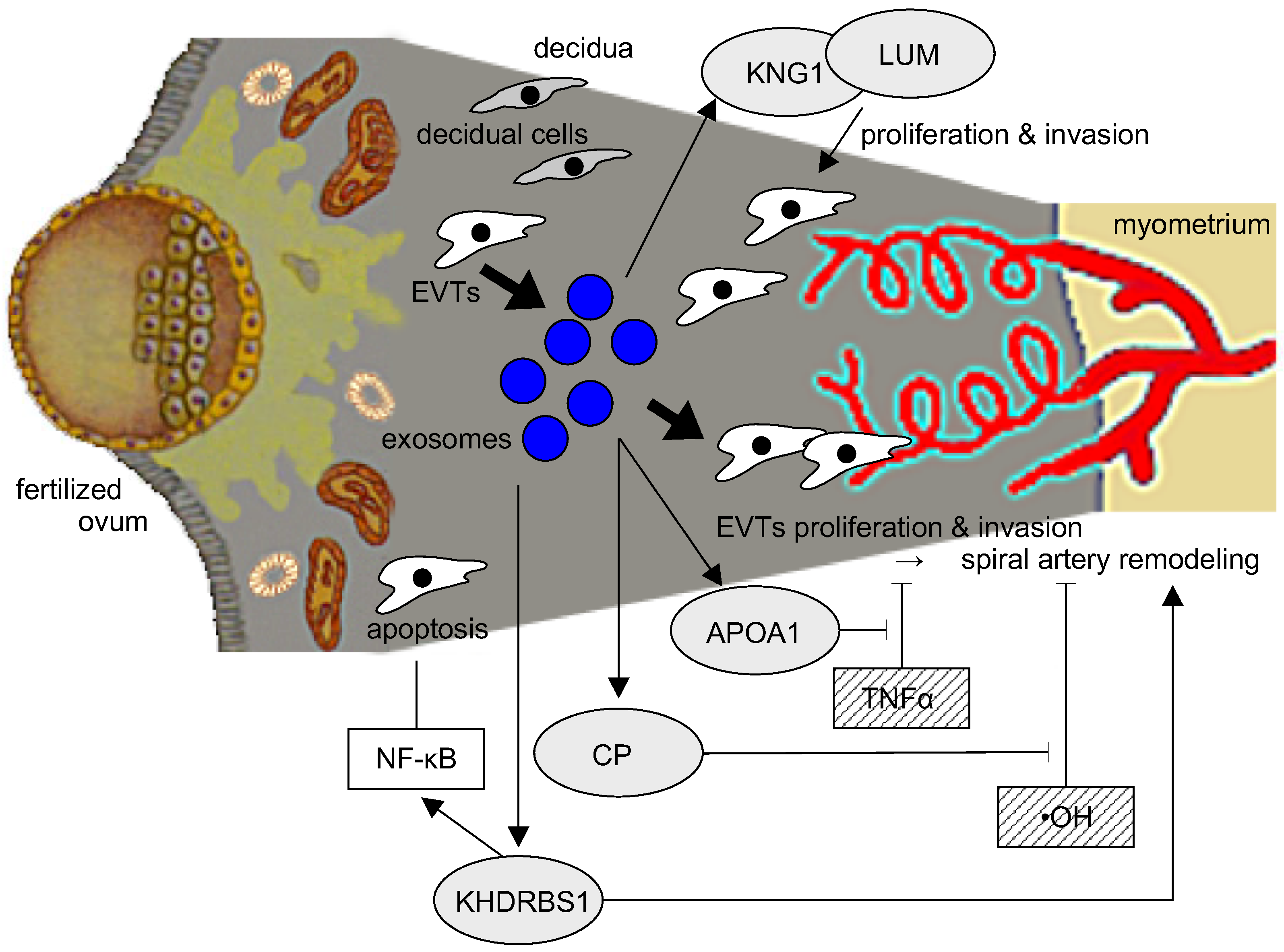
3. Discussion
4. Materials and Methods
4.1. Animal Experiments with a PE Mouse Model
4.2. Cell Culture
4.3. Extraction of Exosomes
4.4. Proteomics Sample Preparation
4.5. Nano LC-MS Analysis
4.6. Statistical Analysis
4.7. DIA (Data Independent Acquisition) Proteomic Analysis
4.8. Bioinformatic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Redman, C.W.; Sargent, I.L. Placental stress and pre-eclampsia: A revised view. Placenta 2009, 30 (Suppl. A), S38–S42. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.M.; Hubel, C.A. The two stage model of preeclampsia: Variations on the theme. Placenta 2009, 30 (Suppl. A), S32–S37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granger, J.P.; Spradley, F.T.; Bakrania, B.A. The Endothelin System: A Critical Player in the Pathophysiology of Preeclampsia. Curr. Hypertens. Rep. 2018, 20, 32. [Google Scholar] [CrossRef] [PubMed]
- Kilbourn, R.G.; Traber, D.L.; Szabó, C. Nitric oxide and shock. Dis. Mon. 1997, 43, 277–348. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K.; Matsubara, Y.; Hyodo, S.; Katayama, T.; Ito, M. Role of nitric oxide and reactive oxygen species in the pathogenesis of preeclampsia. J. Obstet. Gynaecol. Res. 2010, 36, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.W.; Wang, Y. Deficient glutathione peroxidase activity in preeclampsia is associated with increased placental production of thromboxane and lipid peroxides. Am. J. Obstet. Gynecol. 1993, 169, 1456–1461. [Google Scholar] [CrossRef]
- Salomon, C.; Yee, S.W.; Mitchell, M.D.; Rice, G.E. The possible role of extravillous trophoblast-derived exosomes on the uterine spiral arterial remodeling under both normal and pathological conditions. BioMed Res. Int. 2014, 2014, 693157. [Google Scholar] [CrossRef]
- Biró, O.; Alasztics, B.; Molvarec, A.; Joó, J.; Nagy, B.; Rigó, J., Jr. Various levels of circulating exosomal total-miRNA and miR-210 hypoxamiR in different forms of pregnancy hypertension. Pregnancy Hypertens. 2017, 10, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Teng, X.; Chen, L.; Chen, W.; Yang, J.; Yang, Z.; Shen, Z. Mesenchymal Stem Cell-Derived Exosomes Improve the Microenvironment of Infarcted Myocardium Contributing to Angiogenesis and Anti-Inflammation. Cell. Physiol. Biochem. 2015, 37, 2415–2424. [Google Scholar] [CrossRef]
- England, L.; Zhang, J. Smoking and risk of preeclampsia: A systematic review. Front. Biosci. 2007, 12, 2471–2483. [Google Scholar] [CrossRef] [Green Version]
- Hammoud, A.O.; Bujold, E.; Sorokin, Y.; Schild, C.; Krapp, M.; Baumann, P. Smoking in pregnancy revisited: Findings from a large population-based study. Am. J. Obstet. Gynecol. 2005, 192, 1856–1862; discussion 1862–1853. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, S.; Maeda, T.; Kume, T.; Kochiyama, H.; Akaike, A.; Shimohama, S.; Kimura, J. Nicotine protects cultured cortical neurons against glutamate-induced cytotoxicity via alpha7-neuronal receptors and neuronal CNS receptors. Brain Res. 1997, 765, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Michalak, E.; Halko-Gąsior, A.; Chomyszyn-Gajewska, M. The impact of tobacco on oral health—Based on literature. Prz. Lek. 2016, 73, 516–519. [Google Scholar]
- West, R. Tobacco smoking: Health impact, prevalence, correlates and interventions. Psychol. Health 2017, 32, 1018–1036. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, A.; Wu, S.Y.; Sharma, S.; Wu, K.; Zhao, D.; Deshpande, R.; Singh, R.; Li, W.; Topaloglu, U.; Ruiz, J.; et al. Exosomal miR-4466 from nicotine-activated neutrophils promotes tumor cell stemness and metabolism in lung cancer metastasis. Oncogene 2022, 41, 3079–3092. [Google Scholar] [CrossRef]
- Zhu, J.; Liu, B.; Wang, Z.; Wang, D.; Ni, H.; Zhang, L.; Wang, Y. Exosomes from nicotine-stimulated macrophages accelerate atherosclerosis through miR-21-3p/PTEN-mediated VSMC migration and proliferation. Theranostics 2019, 9, 6901–6919. [Google Scholar] [CrossRef]
- Conde-Agudelo, A.; Althabe, F.; Belizán, J.M.; Kafury-Goeta, A.C. Cigarette smoking during pregnancy and risk of preeclampsia: A systematic review. Am. J. Obstet. Gynecol. 1999, 181, 1026–1035. [Google Scholar] [CrossRef]
- Boswijk, E.; Bauwens, M.; Mottaghy, F.M.; Wildberger, J.E.; Bucerius, J. Potential of α7 nicotinic acetylcholine receptor PET imaging in atherosclerosis. Methods 2017, 130, 90–104. [Google Scholar] [CrossRef]
- Moccia, F.; Frost, C.; Berra-Romani, R.; Tanzi, F.; Adams, D.J. Expression and function of neuronal nicotinic ACh receptors in rat microvascular endothelial cells. Am. J. Physiol. Heart Circ. Physiol. 2004, 286, H486–H491. [Google Scholar] [CrossRef] [Green Version]
- Conklin, B.S.; Zhao, W.; Zhong, D.S.; Chen, C. Nicotine and cotinine up-regulate vascular endothelial growth factor expression in endothelial cells. Am. J. Pathol. 2002, 160, 413–418. [Google Scholar] [CrossRef] [Green Version]
- Heeschen, C.; Weis, M.; Aicher, A.; Dimmeler, S.; Cooke, J.P. A novel angiogenic pathway mediated by non-neuronal nicotinic acetylcholine receptors. J. Clin. Investig. 2002, 110, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.O.; Wright, S.M. Production of endothelin by cultured human endothelial cells following exposure to nicotine or caffeine. Metabolism 1999, 48, 845–848. [Google Scholar] [CrossRef]
- Mayhan, W.G.; Sharpe, G.M.; Anding, P. Agonist-induced release of nitric oxide during acute exposure to nicotine. Life Sci. 1999, 65, 1829–1837. [Google Scholar] [CrossRef] [PubMed]
- Nadler, J.L.; Velasco, J.S.; Horton, R. Cigarette smoking inhibits prostacyclin formation. Lancet 1983, 1, 1248–1250. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Lodhi, S.; Sahota, P.; Thakkar, M.M. Nicotine administration in the wake-promoting basal forebrain attenuates sleep-promoting effects of alcohol. J. Neurochem. 2015, 135, 323–331. [Google Scholar] [CrossRef] [Green Version]
- Mimura, K.; Tomimatsu, T.; Sharentuya, N.; Tskitishvili, E.; Kinugasa-Taniguchi, Y.; Kanagawa, T.; Kimura, T. Nicotine restores endothelial dysfunction caused by excess sFlt1 and sEng in an in vitro model of preeclamptic vascular endothelium: A possible therapeutic role of nicotinic acetylcholine receptor (nAChR) agonists for preeclampsia. Am. J. Obstet. Gynecol. 2010, 202, 4464.e1-6. [Google Scholar] [CrossRef]
- Klisch, K.; Schraner, E.M. Intraluminal vesicles of binucleate trophoblast cell granules are a possible source of placental exosomes in ruminants. Placenta 2020, 90, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Li, Q.; Zhang, Q.; Li, Z.; Yao, X.; Guo, Y.; Xiao, L.; Wang, X.; Ni, H. Exosomes derived from placental trophoblast cells regulate endometrial epithelial receptivity in dairy cows during pregnancy. J. Reprod. Dev. 2022, 68, 21–29. [Google Scholar] [CrossRef]
- Erices, R.; Corthorn, J.; Lisboa, F.; Valdés, G. Bradykinin promotes migration and invasion of human immortalized trophoblasts. Reprod. Biol. Endocrinol. 2011, 9, 97. [Google Scholar] [CrossRef] [Green Version]
- Maurer, M.; Bader, M.; Bas, M.; Bossi, F.; Cicardi, M.; Cugno, M.; Howarth, P.; Kaplan, A.; Kojda, G.; Leeb-Lundberg, F.; et al. New topics in bradykinin research. Allergy 2011, 66, 1397–1406. [Google Scholar] [CrossRef]
- Appunni, S.; Rubens, M.; Ramamoorthy, V.; Anand, V.; Khandelwal, M.; Saxena, A.; McGranaghan, P.; Odia, Y.; Kotecha, R.; Sharma, A. Lumican, pro-tumorigenic or anti-tumorigenic: A conundrum. Clin. Chim. Acta 2021, 514, 1–7. [Google Scholar] [CrossRef]
- Giatagana, E.M.; Berdiaki, A.; Tsatsakis, A.; Tzanakakis, G.N.; Nikitovic, D. Lumican in Carcinogenesis-Revisited. Biomolecules 2021, 11, 1319. [Google Scholar] [CrossRef]
- Cooke, A.L.; Morris, J.; Melchior, J.T.; Street, S.E.; Jerome, W.G.; Huang, R.; Herr, A.B.; Smith, L.E.; Segrest, J.P.; Remaley, A.T.; et al. A thumbwheel mechanism for APOA1 activation of LCAT activity in HDL. J. Lipid Res. 2018, 59, 1244–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charlton, F.; Bobek, G.; Stait-Gardner, T.; Price, W.S.; Mirabito Colafella, K.M.; Xu, B.; Makris, A.; Rye, K.A.; Hennessy, A. The protective effect of apolipoprotein in models of trophoblast invasion and preeclampsia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2017, 312, R40–R48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, W.M.; Li, L.J.; Chiu, I.A.; Lai, T.C.; Chang, Y.C.; Tsai, H.F.; Yang, C.J.; Huang, M.S.; Su, C.Y.; Lai, T.L.; et al. The aberrant cancer metabolic gene carbohydrate sulfotransferase 11 promotes non-small cell lung cancer cell metastasis via dysregulation of ceruloplasmin and intracellular iron balance. Transl. Oncol. 2022, 25, 101508. [Google Scholar] [CrossRef] [PubMed]
- Fu, K.; Sun, X.; Wier, E.M.; Hodgson, A.; Liu, Y.; Sears, C.L.; Wan, F. Sam68/KHDRBS1 is critical for colon tumorigenesis by regulating genotoxic stress-induced NF-κB activation. Elife 2016, 5, e15018. [Google Scholar] [CrossRef] [PubMed]
- Pillay, P.; Moodley, K.; Moodley, J.; Mackraj, I. Placenta-derived exosomes: Potential biomarkers of preeclampsia. Int. J. Nanomed. 2017, 12, 8009–8023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, Y.; Zhang, S.T.; Zhu, S.T. Nicotine enhances migration and invasion of human esophageal squamous carcinoma cells which is inhibited by nimesulide. World J. Gastroenterol. 2009, 15, 2500–2505. [Google Scholar] [CrossRef] [PubMed]
- Knight, M.; Redman, C.W.; Linton, E.A.; Sargent, I.L. Shedding of syncytiotrophoblast microvilli into the maternal circulation in pre-eclamptic pregnancies. Br. J. Obstet. Gynaecol. 1998, 105, 632–640. [Google Scholar] [CrossRef]
- Redman, C.W.; Sargent, I.L. Placental debris, oxidative stress and pre-eclampsia. Placenta 2000, 21, 597–602. [Google Scholar] [CrossRef]
- Germain, S.J.; Sacks, G.P.; Sooranna, S.R.; Sargent, I.L.; Redman, C.W. Systemic inflammatory priming in normal pregnancy and preeclampsia: The role of circulating syncytiotrophoblast microparticles. J. Immunol. 2007, 178, 5949–5956. [Google Scholar] [CrossRef] [Green Version]
- Redman, C.W.; Sargent, I.L. Microparticles and immunomodulation in pregnancy and pre-eclampsia. J. Reprod. Immunol. 2007, 76, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Li, W.; Li, P.; Zheng, Z.; Lin, B.; Zhou, B.; Guo, K.; He, P.; Yang, J. Stimulation of α7 Nicotinic Acetylcholine Receptor by Nicotine Suppresses Decidual M1 Macrophage Polarization Against Inflammation in Lipopolysaccharide-Induced Preeclampsia-Like Mouse Model. Front. Immunol. 2021, 12, 642071. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, J.; Bao, J.; Li, X.; Ye, A.; Zhang, G.; Liu, H. Activation of the cholinergic anti-inflammatory pathway by nicotine ameliorates lipopolysaccharide-induced preeclampsia-like symptoms in pregnant rats. Placenta 2017, 49, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, A.; Koide, K.; Hasegawa, J.; Arakaki, T.; Takenaka, S.; Maruyama, D.; Matsuoka, R.; Sekizawa, A. Maternal Smoking History Enhances the Expression of Placental Growth Factor in Invasive Trophoblasts at Early Gestation Despite Cessation of Smoking. PLoS ONE 2015, 10, e0134181. [Google Scholar] [CrossRef]
- Levine, R.J.; Maynard, S.E.; Qian, C.; Lim, K.H.; England, L.J.; Yu, K.F.; Schisterman, E.F.; Thadhani, R.; Sachs, B.P.; Epstein, F.H.; et al. Circulating angiogenic factors and the risk of preeclampsia. N. Engl. J. Med. 2004, 350, 672–683. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Klebanoff, M.A.; Levine, R.J.; Puri, M.; Moyer, P. The puzzling association between smoking and hypertension during pregnancy. Am. J. Obstet. Gynecol. 1999, 181, 1407–1413. [Google Scholar] [CrossRef]
- Morisaki, N.; Obara, T.; Piedvache, A.; Kobayashi, S.; Miyashita, C.; Nishimura, T.; Ishikuro, M.; Sata, F.; Horikawa, R.; Mori, C.; et al. Association between smoking and hypertension in pregnancy among Japanese women: A meta-analysis of birth cohort studies in the Japan Birth Cohort Consortium (JBiCC) and JECS. J. Epidemiol. 2022. [Google Scholar] [CrossRef]
- Chen, J.; Qiu, M.; Huang, Z.; Chen, J.; Zhou, C.; Han, F.; Qu, Y.; Wang, S.; Zhuang, J.; Li, X. Nicotine Suppresses the Invasiveness of Human Trophoblasts by Downregulation of CXCL12 Expression through the Alpha-7 Subunit of the Nicotinic Acetylcholine Receptor. Reprod. Sci. 2020, 27, 916–924. [Google Scholar] [CrossRef] [Green Version]
- Matsubara, K.; Abe, E.; Ochi, H.; Kusanagi, Y.; Ito, M. Changes in serum concentrations of tumor necrosis factor alpha and adhesion molecules in normal pregnant women and those with pregnancy-induced hypertension. J. Obstet. Gynaecol. Res. 2003, 29, 422–426. [Google Scholar] [CrossRef]
- Matsubara, K.; Matsubara, Y.; Mori, M.; Uchikura, Y.; Hamada, K.; Fujioka, T.; Hashimoto, H.; Matsumoto, T. Immune activation during the implantation phase causes preeclampsia-like symptoms via the CD40-CD40 ligand pathway in pregnant mice. Hypertens. Res. 2016, 39, 407–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cione, E.; Cannataro, R.; Gallelli, L.; De Sarro, G.; Caroleo, M.C. Exosome microRNAs in Metabolic Syndrome as Tools for the Early Monitoring of Diabetes and Possible Therapeutic Options. Pharmaceuticals 2021, 14, 1257. [Google Scholar] [CrossRef] [PubMed]
- Leuchtmann, A.B.; Adak, V.; Dilbaz, S.; Handschin, C. The Role of the Skeletal Muscle Secretome in Mediating Endurance and Resistance Training Adaptations. Front. Physiol. 2021, 12, 709807. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Downregulated | |||
|---|---|---|---|
| Accession Number | Protein Name | log2FC | p Value |
| Q9UGI8 | Testin | 2.67 | p < 0.05 |
| Q02818 | Nucleobindin-1 | 2.63 | p < 0.001 |
| P13591 | Neural cell adhesion molecule 1 | 2.60 | p < 0.001 |
| P02788 | Lactotransferrin | 2.40 | p < 0.0005 |
| P28827 | Receptor-type tyrosine-protein phosphatase mu | 2.39 | p < 0.005 |
| P05114 | Non-histone chromosomal protein HMG-14 | 2.31 | p < 0.01 |
| P24592 | Insulin-like growth factor-binding protein 6 | 2.23 | p < 0.05 |
| P55285 | Cadherin-6 | 2.21 | p < 0.01 |
| Q05682 | Caldesmon | 2.06 | p < 0.001 |
| O00533 | Neural cell adhesion molecule L1-like protein | 2.00 | p < 0.005 |
| Upregulated | |||
| Accession Number | Protein Name | log2FC | p Value |
| Q6GTS8 | N-fatty-acyl-amino acid synthase/hydrolase | −2.08 | p < 0.0005 |
| Q7Z7L7 | Protein zer-1 homolog | −2.00 | p < 0.05 |
| Q14139 | Serine/threonine-protein phosphatase 6 catalytic subunit | −1.78 | p < 0.0005 |
| Q14997 | Proteasome activator complex subunit 4 | −1.66 | p < 0.05 |
| Q03001 | Dystonin | −1.51 | p < 0.05 |
| O60341 | Lysine-specific histone demethylase 1A | −1.42 | p < 0.0001 |
| P68871 | Hemoglobin subunit beta | −1.35 | p < 0.05 |
| P40306 | Proteasome subunit beta type-10 | −1.34 | p < 0.05 |
| Q9ULHO | Kinase D-interacting substrate of 220 kDa | −1.26 | p < 0.001 |
| Q13033 | Striatin-3 | −1.15 | p < 0.005 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kubo, A.; Matsubara, K.; Matsubara, Y.; Nakaoka, H.; Sugiyama, T. The Influence of Nicotine on Trophoblast-Derived Exosomes in a Mouse Model of Pathogenic Preeclampsia. Int. J. Mol. Sci. 2023, 24, 11126. https://doi.org/10.3390/ijms241311126
Kubo A, Matsubara K, Matsubara Y, Nakaoka H, Sugiyama T. The Influence of Nicotine on Trophoblast-Derived Exosomes in a Mouse Model of Pathogenic Preeclampsia. International Journal of Molecular Sciences. 2023; 24(13):11126. https://doi.org/10.3390/ijms241311126
Chicago/Turabian StyleKubo, Ayane, Keiichi Matsubara, Yuko Matsubara, Hirotomo Nakaoka, and Takashi Sugiyama. 2023. "The Influence of Nicotine on Trophoblast-Derived Exosomes in a Mouse Model of Pathogenic Preeclampsia" International Journal of Molecular Sciences 24, no. 13: 11126. https://doi.org/10.3390/ijms241311126