
Elevation of Arginase-II in Podocytes Contributes to Age-Associated Albuminuria in Male Mice



Abstract
:1. Introduction
2. Results
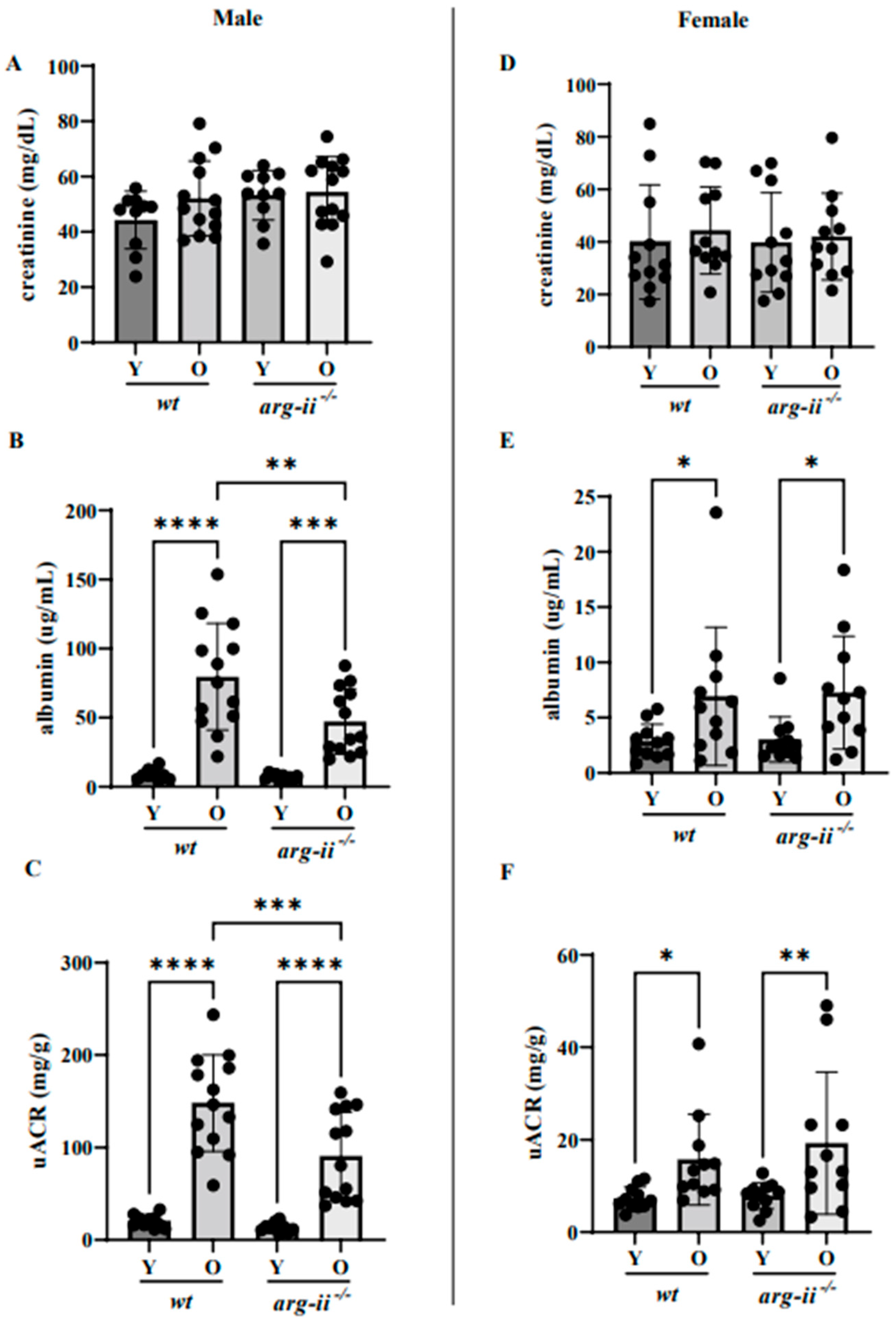
2.1. arg-ii Ablation Decreases Age-Associated Albuminuria in Male but Not Female Mice
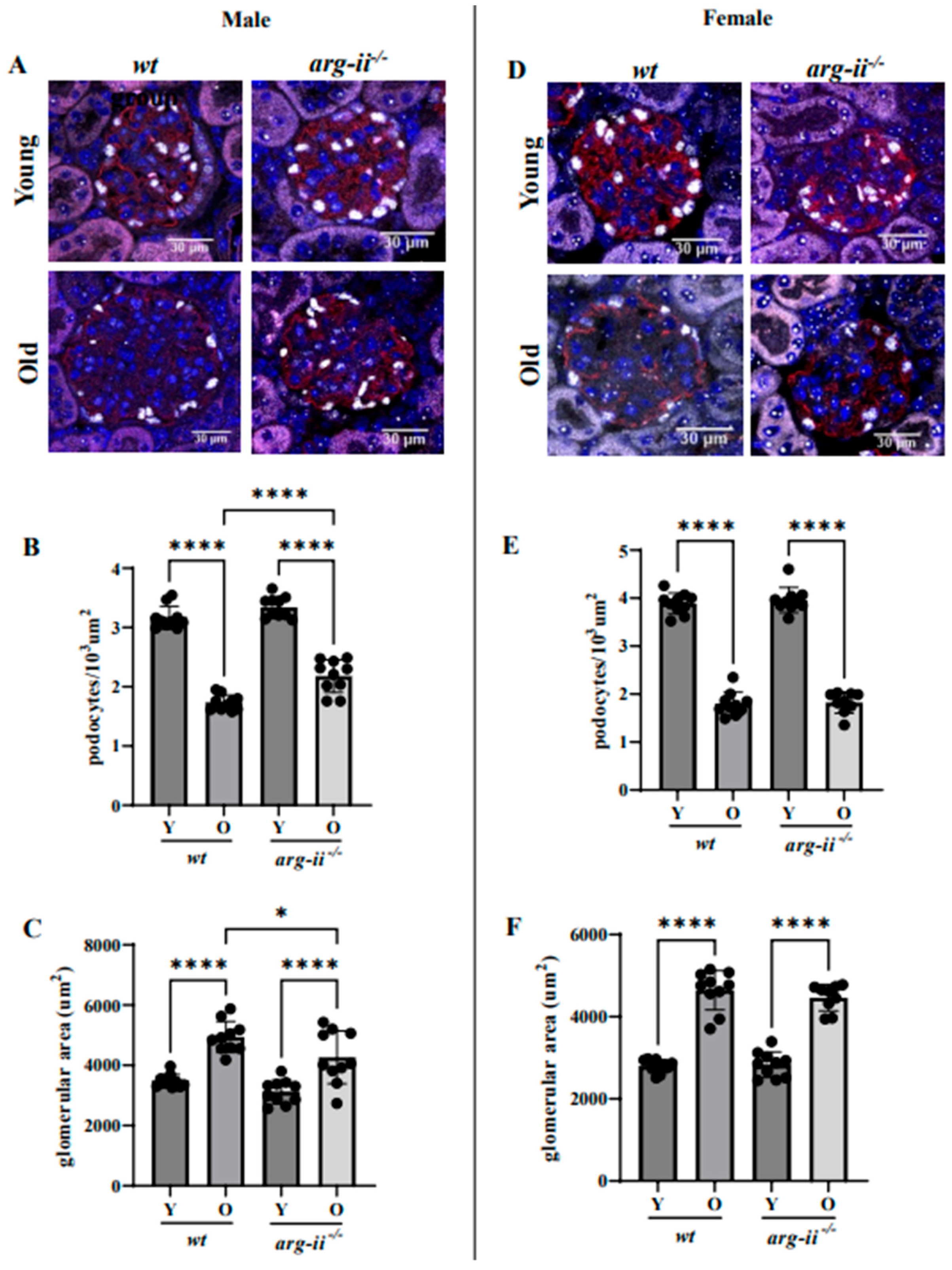
2.2. arg-ii Ablation Protects Male Mice from Age-Associated Decreases in Podocytes
2.3. arg-ii Ablation Has No Effects on Age-Associated Glomerulosclerosis in Both Sexes
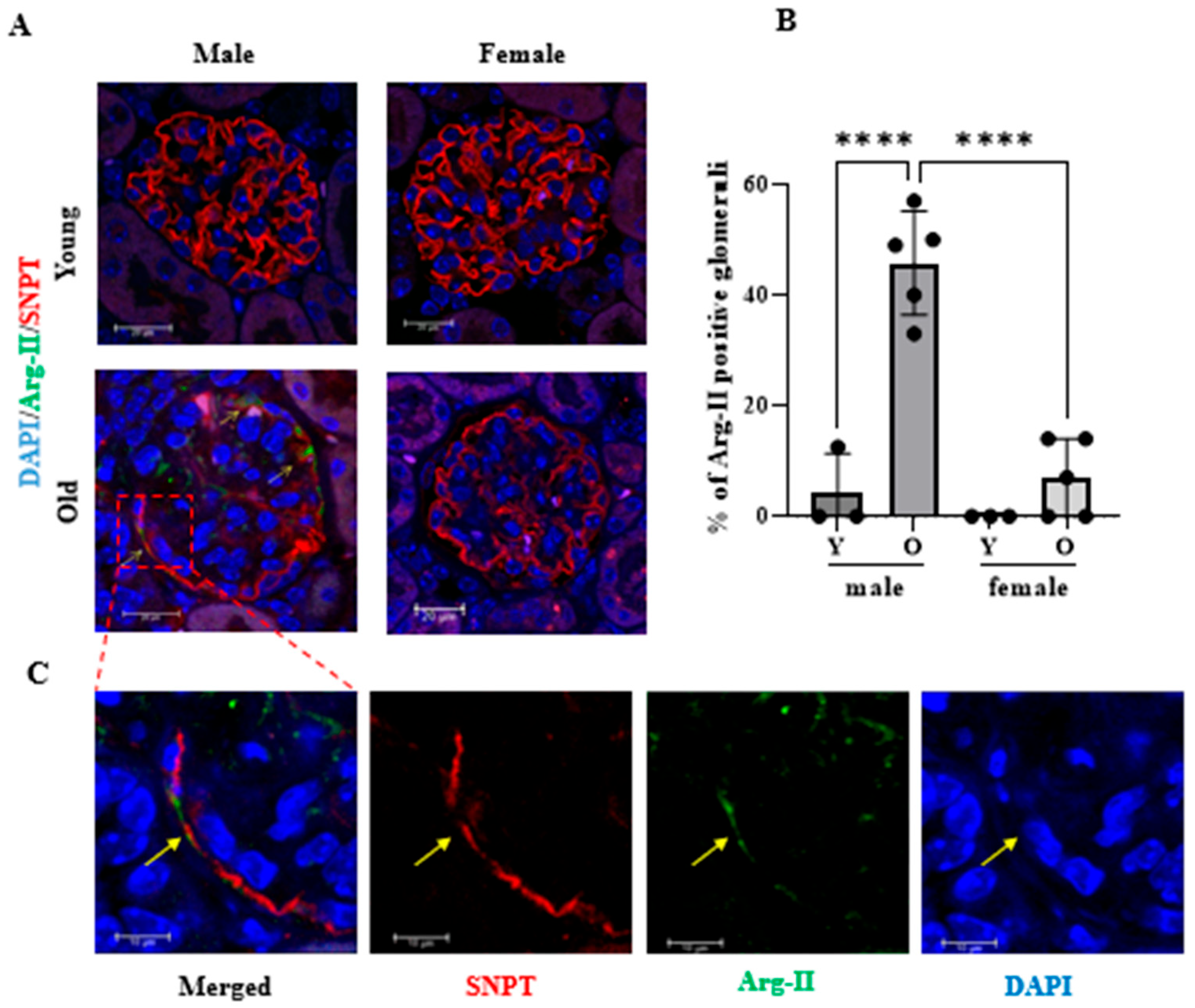
2.4. Age-Dependent Increase in Arg-II Level in Podocytes in Male but Not in Female Mice
3. Discussion
Limitations and Future Perspectives
4. Materials and Methods
4.1. arg-ii−/− Mouse and Sample Preparation
4.2. Quantification of Podocytes
4.3. Immunofluorescence Staining
4.4. Urine Collection, Measurement of Urine Albumin and Creatinine
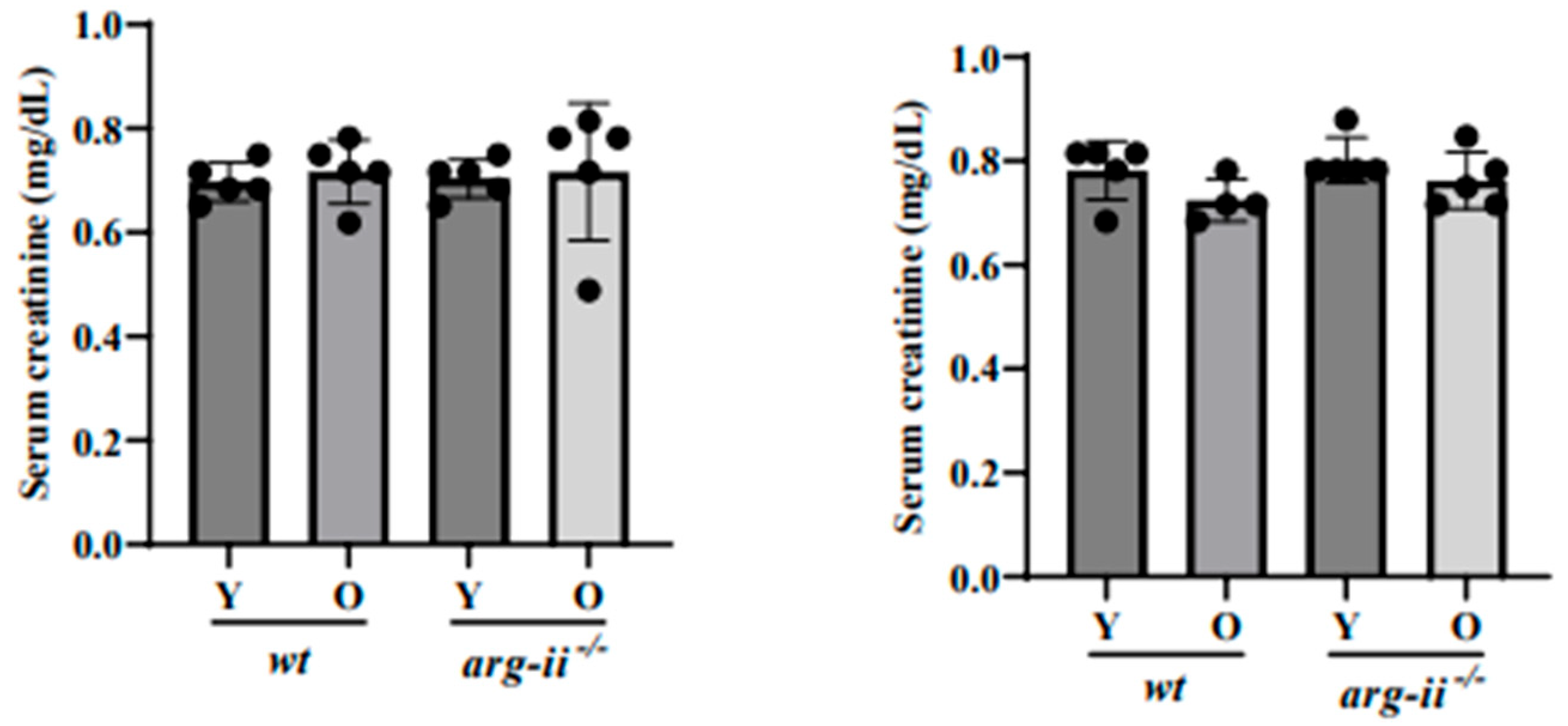
4.5. Blood Collection and Measurement of Serum Creatinine
4.6. Quantification of Glomerulosclerosis
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Arg-II | Arginase-II |
| PAS | Periodic acid-Schiff |
| PCR | polymerase chain reaction |
| SNPT | synaptopodin |
| WT1 | Wilms tumor protein 1 |
| uACR | Urinary albumin/creatinine ratio |
References
- Marquez-Exposito, L.; Tejedor-Santamaria, L.; Santos-Sanchez, L.; Valentijn, F.A.; Cantero-Navarro, E.; Rayego-Mateos, S.; Rodrigues-Diez, R.R.; Tejera-Munoz, A.; Marchant, V.; Sanz, A.B.; et al. Acute Kidney Injury is Aggravated in Aged Mice by the Exacerbation of Proinflammatory Processes. Front. Pharmacol. 2021, 12, 662020. [Google Scholar] [CrossRef]
- Chou, Y.H.; Lai, T.S.; Lin, Y.C.; Chiang, W.C.; Chu, T.S.; Lin, S.L.; Chen, Y.M. Age-Dependent Effects of Acute Kidney Injury on End-Stage Kidney Disease and Mortality in Patients with Moderate to Severe Chronic Kidney Disease. Nephron 2023, 147, 329–336. [Google Scholar] [CrossRef]
- O’Sullivan, E.D.; Hughes, J.; Ferenbach, D.A. Renal Aging: Causes and Consequences. J. Am. Soc. Nephrol. JASN 2017, 28, 407–420. [Google Scholar] [CrossRef] [Green Version]
- Rule, A.D.; Amer, H.; Cornell, L.D.; Taler, S.J.; Cosio, F.G.; Kremers, W.K.; Textor, S.C.; Stegall, M.D. The association between age and nephrosclerosis on renal biopsy among healthy adults. Ann. Intern. Med. 2010, 152, 561–567. [Google Scholar] [CrossRef] [Green Version]
- Minutolo, R.; Gabbai, F.B.; Chiodini, P.; Provenzano, M.; Borrelli, S.; Garofalo, C.; Bellizzi, V.; Russo, D.; Conte, G.; De Nicola, L.; et al. Sex Differences in the Progression of CKD Among Older Patients: Pooled Analysis of 4 Cohort Studies. Am. J. Kidney Dis. 2020, 75, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Beckwith, H.; Lightstone, L.; McAdoo, S. Sex and Gender in Glomerular Disease. Semin. Nephrol. 2022, 42, 185–196. [Google Scholar] [CrossRef]
- Gotoh, T.; Sonoki, T.; Nagasaki, A.; Terada, K.; Takiguchi, M.; Mori, M. Molecular cloning of cDNA for nonhepatic mitochondrial arginase (arginase II) and comparison of its induction with nitric oxide synthase in a murine macrophage-like cell line. FEBS Lett. 1996, 395, 119–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vockley, J.G.; Jenkinson, C.P.; Shukla, H.; Kern, R.M.; Grody, W.W.; Cederbaum, S.D. Cloning and characterization of the human type II arginase gene. Genomics 1996, 38, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Park, C.; Ahn, M.; Lee, J.H.; Shin, T. Immunohistochemical study of arginase 1 and 2 in various tissues of rats. Acta Histochem. 2012, 114, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Rajapakse, A.; Xiong, Y.; Montani, J.-P.; Verrey, F.; Ming, X.-F.; Yang, Z. Genetic Targeting of Arginase-II in Mouse Prevents Renal Oxidative Stress and Inflammation in Diet-Induced Obesity. Front. Physiol. 2016, 7, 560. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Montani, J.P.; Verrey, F.; Feraille, E.; Ming, X.F.; Yang, Z. Arginase-II negatively regulates renal aquaporin-2 and water reabsorption. FASEB J. 2018, 32, 5520–5531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Z.; Potenza, D.M.; Ma, Y.; Ajalbert, G.; Hoogewijs, D.; Ming, X.F.; Yang, Z. Role of Arginase-II in Podocyte Injury under Hypoxic Conditions. Biomolecules 2022, 12, 1213. [Google Scholar] [CrossRef] [PubMed]
- Wetzel, M.D.; Stanley, K.; Wang, W.W.; Maity, S.; Madesh, M.; Reeves, W.B.; Awad, A.S. Selective inhibition of arginase-2 in endothelial cells but not proximal tubules reduces renal fibrosis. JCI Insight 2020, 5, e142187. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Torisu, K.; Tomita, K.; Kawai, Y.; Tsuruya, K.; Nakano, T.; Kitazono, T. Arginase 2 is a mediator of ischemia-reperfusion injury in the kidney through regulation of nitrosative stress. Kidney Int. 2020, 98, 673–685. [Google Scholar] [CrossRef]
- Liang, X.; Potenza, D.M.; Brenna, A.; Ma, Y.; Ren, Z.; Cheng, X.; Ming, X.F.; Yang, Z. Hypoxia Induces Renal Epithelial Injury and Activates Fibrotic Signaling Through Up-Regulation of Arginase-II. Front. Physiol. 2021, 12, 773719. [Google Scholar] [CrossRef]
- Huang, J.; Liang, X.; Ladeiras, D.; Fellay, B.; Ming, X.F.; Yang, Z. Role of tubular epithelial arginase-II in renal inflammaging. NPJ Aging Mech. Dis. 2021, 7, 5. [Google Scholar] [CrossRef]
- Morris, S.M., Jr.; Gao, T.; Cooper, T.K.; Kepka-Lenhart, D.; Awad, A.S. Arginase-2 mediates diabetic renal injury. Diabetes 2011, 60, 3015–3022. [Google Scholar] [CrossRef] [Green Version]
- Yepuri, G.; Velagapudi, S.; Xiong, Y.Y.; Rajapakse, A.G.; Montani, J.P.; Ming, X.F.; Yang, Z. Positive crosstalk between arginase-II and S6K1 in vascular endothelial inflammation and aging. Aging Cell 2012, 11, 1005–1016. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Yepuri, G.; Montani, J.P.; Ming, X.F.; Yang, Z. Arginase-II Deficiency Extends Lifespan in Mice. Front. Physiol. 2017, 8, 682. [Google Scholar] [CrossRef]
- Xiong, Y.; Yepuri, G.; Necetin, S.; Montani, J.P.; Ming, X.F.; Yang, Z. Arginase-II Promotes Tumor Necrosis Factor-alpha Release From Pancreatic Acinar Cells Causing beta-Cell Apoptosis in Aging. Diabetes 2017, 66, 1636–1649. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Potenza, D.M.; Yang, Y.; Ajalbert, G.; Mertz, K.D.; von Gunten, S.; Ming, X.F.; Yang, Z. Role of pulmonary epithelial arginase-II in activation of fibroblasts and lung inflammaging. Aging Cell 2023, 22, e13790. [Google Scholar] [CrossRef] [PubMed]
- Siwinska, N.; Paslawska, U.; Bachor, R.; Szczepankiewicz, B.; Zak, A.; Grocholska, P.; Szewczuk, Z. Evaluation of podocin in urine in horses using qualitative and quantitative methods. PLoS ONE 2020, 15, e0240586. [Google Scholar] [CrossRef] [PubMed]
- Ichii, O.; Nakamura, T.; Irie, T.; Otani, Y.; Hosotani, M.; Masum, M.A.; Islam, R.M.; Horino, T.; Sunden, Y.; Elewa, Y.H.A.; et al. Age-related glomerular lesions with albuminuria in male cotton rats. Histochem. Cell Biol. 2020, 153, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Balzer, M.S.; Rohacs, T.; Susztak, K. How Many Cell Types Are in the Kidney and What Do They Do? Annu. Rev. Physiol. 2022, 84, 507–531. [Google Scholar] [CrossRef]
- Raup-Konsavage, W.M.; Gao, T.; Cooper, T.K.; Morris, S.M., Jr.; Reeves, W.B.; Awad, A.S. Arginase-2 mediates renal ischemia-reperfusion injury. Am. J. Physiol. Renal. Physiol. 2017, 313, F522–F534. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Long, J.; Mise, K.; Galvan, D.L.; Overbeek, P.A.; Tan, L.; Kumar, S.V.; Chan, W.K.; Lorenzi, P.L.; Chang, B.H.; et al. PGC1alpha is required for the renoprotective effect of lncRNA Tug1 in vivo and links Tug1 with urea cycle metabolites. Cell Rep. 2021, 36, 109510. [Google Scholar] [CrossRef]
- Moeller, M.J.; Kramann, R.; Lammers, T.; Hoppe, B.; Latz, E.; Ludwig-Portugall, I.; Boor, P.; Floege, J.; Kurts, C.; Weiskirchen, R.; et al. New Aspects of Kidney Fibrosis-From Mechanisms of Injury to Modulation of Disease. Front. Med. 2021, 8, 814497. [Google Scholar] [CrossRef]
- Austad, S.N.; Bartke, A. Sex Differences in Longevity and in Responses to Anti-Aging Interventions: A Mini-Review. Gerontology 2015, 62, 40–46. [Google Scholar] [CrossRef]
- Holzenberger, M.; Dupont, J.; Ducos, B.; Leneuve, P.; Geloen, A.; Even, P.C.; Cervera, P.; Le, B.Y. IGF-1 receptor regulates lifespan and resistance to oxidative stress in mice. Nature 2003, 421, 182–187. [Google Scholar] [CrossRef] [Green Version]
- Selman, C.; Lingard, S.; Choudhury, A.I.; Batterham, R.L.; Claret, M.; Clements, M.; Ramadani, F.; Okkenhaug, K.; Schuster, E.; Blanc, E.; et al. Evidence for lifespan extension and delayed age-related biomarkers in insulin receptor substrate 1 null mice. FASEB J. 2008, 22, 807–818. [Google Scholar] [CrossRef] [Green Version]
- Selman, C.; Tullet, J.M.; Wieser, D.; Irvine, E.; Lingard, S.J.; Choudhury, A.I.; Claret, M.; Al-Qassab, H.; Carmignac, D.; Ramadani, F.; et al. Ribosomal protein S6 kinase 1 signaling regulates mammalian life span. Science 2009, 326, 140–144, Erratum in Science 2011, 334, 39. [Google Scholar] [CrossRef] [Green Version]
- Lamming, D.W.; Mihaylova, M.M.; Katajisto, P.; Baar, E.L.; Yilmaz, O.H.; Hutchins, A.; Gultekin, Y.; Gaither, R.; Sabatini, D.M. Depletion of Rictor, an essential protein component of mTORC2, decreases male lifespan. Aging Cell 2014, 13, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Kanfi, Y.; Naiman, S.; Amir, G.; Peshti, V.; Zinman, G.; Nahum, L.; Bar-Joseph, Z.; Cohen, H.Y. The sirtuin SIRT6 regulates lifespan in male mice. Nature 2012, 483, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Doublier, S.; Lupia, E.; Catanuto, P.; Periera-Simon, S.; Xia, X.; Korach, K.; Berho, M.; Elliot, S.J.; Karl, M. Testosterone and 17beta-estradiol have opposite effects on podocyte apoptosis that precedes glomerulosclerosis in female estrogen receptor knockout mice. Kidney Int. 2011, 79, 404–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, T.; Esaki, T.; Sumi, D.; Mukherjee, T.; Iguchi, A.; Chaudhuri, G. Modulating role of estradiol on arginase II expression in hyperlipidemic rabbits as an atheroprotective mechanism. Proc. Natl. Acad. Sci. USA 2006, 103, 10485–10490. [Google Scholar] [CrossRef]
- Levillain, O.; Diaz, J.J.; Blanchard, O.; Dechaud, H. Testosterone down-regulates ornithine aminotransferase gene and up-regulates arginase II and ornithine decarboxylase genes for polyamines synthesis in the murine kidney. Endocrinology 2005, 146, 950–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricardo, A.C.; Yang, W.; Sha, D.; Appel, L.J.; Chen, J.; Krousel-Wood, M.; Manoharan, A.; Steigerwalt, S.; Wright, J.; Rahman, M.; et al. Sex-Related Disparities in CKD Progression. J. Am. Soc. Nephrol. JASN 2019, 30, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Shi, O.; Morris, S.M.; Zoghbi, H.; Porter, C.W.; O’Brien, W.E. Generation of a Mouse Model for Arginase II Deficiency by Targeted Disruption of the Arginase II Gene. Mol. Cell. Biol. 2001, 21, 811–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Percie du Sert, N.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; Emerson, M.; et al. Reporting animal research: Explanation and elaboration for the ARRIVE guidelines 2.0. PLoS Biol. 2020, 18, e3000411. [Google Scholar] [CrossRef] [PubMed]
- Vashistha, H.; Meggs, L. Diabetic nephropathy: Lessons from the mouse. Ochsner. J. 2013, 13, 140–146. [Google Scholar] [PubMed]
- Ma, Y.; Potenza, D.M.; Ajalbert, G.; Brenna, A.; Zhu, C.; Ming, X.F.; Yang, Z. Paracrine Effects of Renal Proximal Tubular Epithelial Cells on Podocyte Injury under Hypoxic Conditions Are Mediated by Arginase-II and TGF-beta1. Int. J. Mol. Sci. 2023, 24, 3587. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, J.F.; Vergara, V.B.; Mog, S.R.; Kalinich, J.F. Hydrophobic Sand Is a Non-Toxic Method of Urine Collection, Appropriate for Urinary Metal Analysis in the Rat. Toxics 2017, 5, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckerman, P.; Bi-Karchin, J.; Park, A.S.; Qiu, C.; Dummer, P.D.; Soomro, I.; Boustany-Kari, C.M.; Pullen, S.S.; Miner, J.H.; Hu, C.A.; et al. Transgenic expression of human APOL1 risk variants in podocytes induces kidney disease in mice. Nat. Med. 2017, 23, 429–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parasuraman, S.; Raveendran, R.; Kesavan, R. Blood sample collection in small laboratory animals. J. Pharmacol. Pharmacother. 2010, 1, 87–93, Erratum in J. Pharmacol. Pharmacother. 2017, 8, 153. [Google Scholar] [CrossRef] [Green Version]
- Menendez-Castro, C.; Nitz, D.; Cordasic, N.; Jordan, J.; Bauerle, T.; Fahlbusch, F.B.; Rascher, W.; Hilgers, K.F.; Hartner, A. Neonatal nephron loss during active nephrogenesis—detrimental impact with long-term renal consequences. Sci. Rep. 2018, 8, 4542. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody Target | Dilution |
|---|---|
| Arg-II (#55003, Cell Signaling; Danvers, MA, USA) | IF 1:100 |
| Synaptopodin (sc-515842, Santa Cruz Biotechnol; Dallas, TX, USA) | IF 1:50 |
| WT1 (ab89901, Abcam; Cambridge, UK) | IF 1:100 |
| Alexa Fluor 488-conjugated goat anti-rabbit IgG (H+L) secondary Ab (A-11008, Thermo Fisher Scientific; Waltham, MA, USA) | IF 1:400 |
| Alexa Fluor 568-conjugated goat anti-mouse IgG (H+L) secondary Ab (A-11031, Thermo Fisher Scientific; Waltham, MA, USA) | IF 1:400 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ajalbert, G.; Brenna, A.; Ming, X.-F.; Yang, Z.; Potenza, D.M. Elevation of Arginase-II in Podocytes Contributes to Age-Associated Albuminuria in Male Mice. Int. J. Mol. Sci. 2023, 24, 11228. https://doi.org/10.3390/ijms241311228
Ajalbert G, Brenna A, Ming X-F, Yang Z, Potenza DM. Elevation of Arginase-II in Podocytes Contributes to Age-Associated Albuminuria in Male Mice. International Journal of Molecular Sciences. 2023; 24(13):11228. https://doi.org/10.3390/ijms241311228
Chicago/Turabian StyleAjalbert, Guillaume, Andrea Brenna, Xiu-Fen Ming, Zhihong Yang, and Duilio M. Potenza. 2023. "Elevation of Arginase-II in Podocytes Contributes to Age-Associated Albuminuria in Male Mice" International Journal of Molecular Sciences 24, no. 13: 11228. https://doi.org/10.3390/ijms241311228