
κO-SrVIA Conopeptide, a Novel Inhibitor Peptide for Two Members of the Human EAG Potassium Channel Family
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
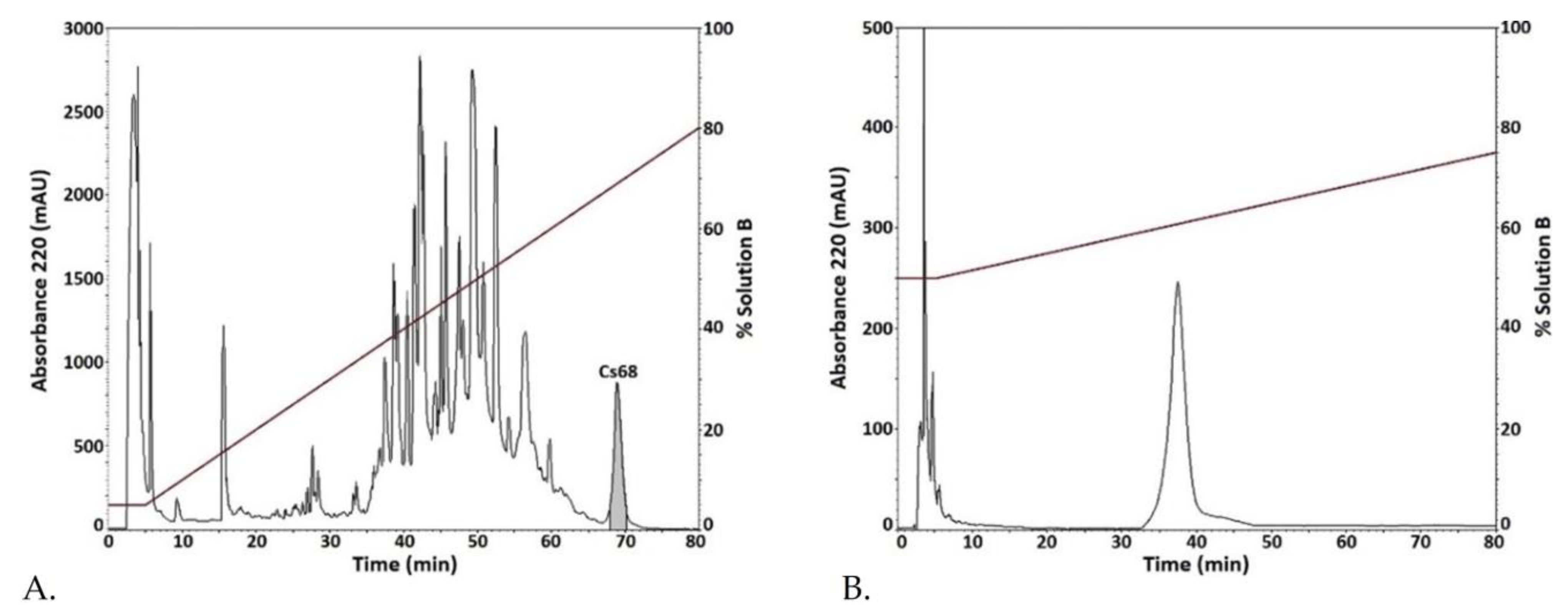
2.1. Conus spurius Venom Fractionation and Purification of Fraction Cs68 Subsection
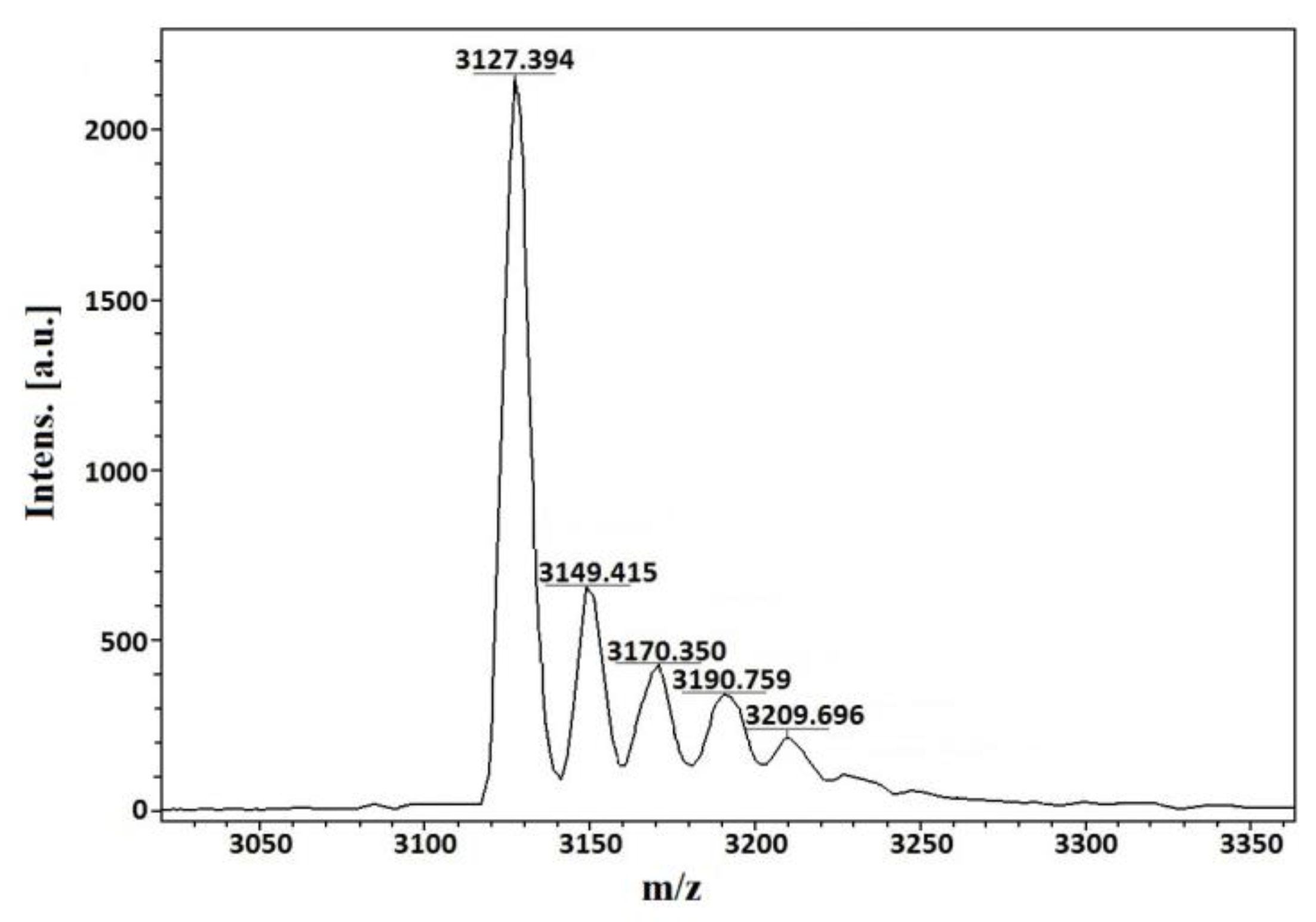
2.2. Molecular Mass and Amino Acid Sequence Analysis
2.3. Similarity Search
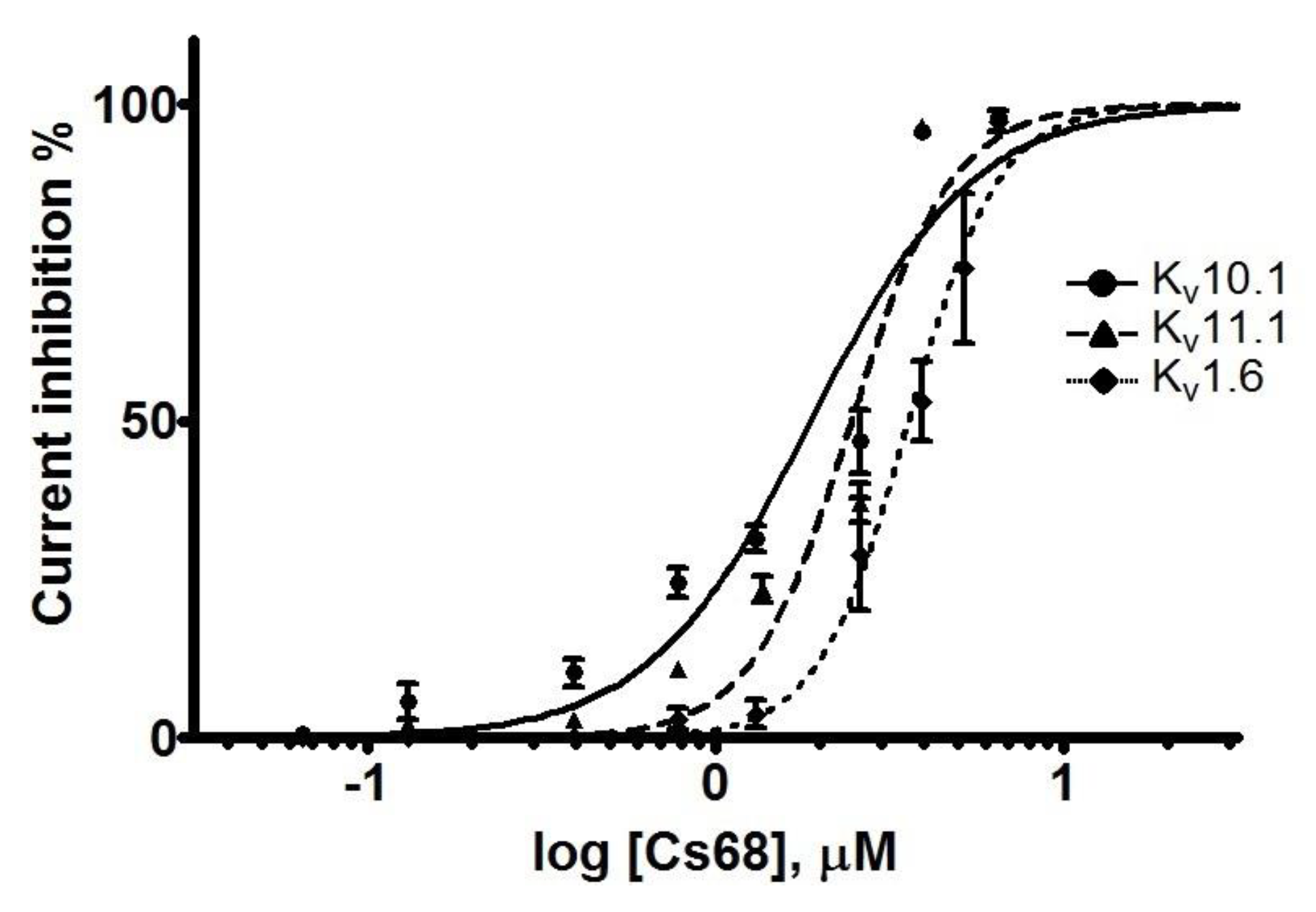
2.4. Electrophysiological Evaluation of Cs68 on Voltage-Gated Potassium Channels
3. Discussion
4. Materials and Methods
4.1. Isolation of Crude Venom Extract from Conus spurius
4.2. Crude Venom Extract Fractionation by RP-HPLC
4.3. Amino Acid Sequencing and Mass Spectrometry Analysis
4.4. Preparation of Vectors Encoding Potassium Channels
4.5. Expression of Voltage-Gated Potassium Channels
4.6. Electrophysiological Recording
4.7. Similarity Search and Sequence Comparison
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hermitte, L.C.D. Venomous marine molluscs of the genus. Conus. Trans. Roy. Soc. Trop. Med. Hyg. 1946, 39, 485–512. [Google Scholar] [CrossRef] [PubMed]
- Endean, R.; Rudkin, C. Further studies of the venoms of Conidae. Toxicon 1965, 2, 225–249. [Google Scholar] [CrossRef] [PubMed]
- Kohn, A.J. Food Specialization in Conus in Hawaii and California. Ecology 1966, 47, 1041–1043. [Google Scholar] [CrossRef]
- Teichert, R.W.; Jimenez, E.C.; Twede, V.; Watkins, M.; Hollmann, M.; Bulaj, G.; Olivera, B.M. Novel conantokins from Conus parius venom are specific antagonists of N-methyl-D-aspartate receptors. J. Biol. Chem. 2007, 282, 36905–36913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, R.; Dutertre, S.; Vetter, I.; Christie, M. Conus venom peptide pharmacology. Pharmacol. Rev. 2012, 6, 259–298. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, I.A.; Gehrmann, J.; Loughnan, M.L.; Thomas, L.; Adams, D.A.; Atkins, A.; Palant, E.; Craik, D.J.; Adams, D.; Alewood, P.F.; et al. Two new classes of conopeptides inhibit the α1-adrenoceptor and noradrenaline transporter. Nat. Neurosci. 2001, 4, 902–907. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, I.; Palant, E.; Schroeder, C.; Kaye, D.; Adams, D.; Alewood, P.; Lewis, R. Inhibition of the norepinephrine transporter by the venom peptide χ-MrIA. J. Biol. Chem. 2003, 278, 40317–40323. [Google Scholar] [CrossRef] [Green Version]
- Petrel, C.; Hocking, H.G.; Reynaud, M.; Upert, G.; Favreau, P.; Biass, D.; Paolini-Bertrand, M.; Peigneur, S.; Tytgat, J.; Gilles, N.; et al. Identification, structural and pharmacological characterization of τ-CnVA, a conopeptide that selectively interacts with somatostatin sst3 receptor. Biochem. Pharmacol. 2013, 85, 1663–1671. [Google Scholar] [CrossRef]
- Finol-Urdaneta, R.K.; Belovanovic, A.; Micic-Vicovac, M.; Kinsella, G.K.; McArthur, J.R.; Al-Sabi, A. Marine toxins targeting Kv1 channels: Pharmacological tools and therapeutic scaffolds. Mar. Drugs 2020, 18, 173. [Google Scholar] [CrossRef] [Green Version]
- Tosti, E.; Boni, R.; Gallo, A. Pathophysiological responses to conotoxin modulation of voltage-gated ion currents. Mar. Drugs 2022, 20, 282. [Google Scholar] [CrossRef]
- Tian, C.; Zhu, R.; Zhu, L.; Qiu, T.; Cao, Z.; Kang, T. Potassium channels: Structures, diseases, and modulators. Chem. Biol. Drug Des. 2014, 83, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, N.; Grunnet, M.; Olesen, S.-P. Cardiac potassium channel subtypes: New roles in repolarization and arrhythmia. Physiol. Rev. 2014, 94, 609–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortensen, L.S.; Schmidt, H.; Farsi, Z.; Barrantes-Freer, A.; Rubio, M.E.; Ufartes, R.; Eilers, J.; Sakaba, T.; Stühmer, W.; Pardo, L.A. Kv10.1 opposes activity-dependent increase in Ca2+ influx into the presynaptic terminal of the parallel fibre–Purkinje cell synapse. J. Physiol. 2015, 593, 181–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peck, L.J.; Patel, R.; Diaz, P.; Wintle, Y.M.; Dickenson, A.H.; Todd, A.J.; Calvo, M.; Bennett, D.L.H. Studying independent Kcna6 knock-out mice reveals toxicity of exogenous LacZ to central nociceptor terminals and differential effects of Kv1.6 on acute and neuropathic pain sensation. J. Neurosci. 2021, 41, 9141–9162. [Google Scholar] [CrossRef] [PubMed]
- Sanguinetti, M.C.; Tristani-Firouzi, M. hERG potassium channels and cardiac arrhythmia. Nature 2006, 440, 463–469. [Google Scholar] [CrossRef]
- Hemmerlein, B.; Weseloh, R.; de Queiroz, F.M.; Knötgen, H.; Sánchez, A.; Rubio, M.; Martin, S.; Schliephacke, T.; Jenke, M.; Heinz-Joachim-Radzun; et al. Overexpression of Eag1 potassium channels in clinical tumours. Mol. Cancer 2006, 5, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamora-Bustillos, R.; Martínez-Núñez, M.A.; Aguilar, M.B.; Collí-Dula, R.C.; Brito-Domínguez, D.A. Identification of novel conotoxin precursors from the cone snail Conus spurius by high-throughput RNA sequencing. Mar. Drugs 2021, 19, 547. [Google Scholar] [CrossRef]
- Kaas, Q.; Yu, R.; Jin, A.-H.; Dutertre, S.; Craik, D.J. ConoServer: Updated content, knowledge, and discovery tools in the conopeptide database. Nucleic Acids Res. 2012, 40, D325–D330. [Google Scholar] [CrossRef]
- García-Ferreiro, R.E.; Kerschensteiner, D.; Major, F.; Monje, F.; Stühmer, W.; Pardo, L.A. Mechanism of block of hEag1 K+ channels by imipramine and astemizole. J. Gen. Physiol. 2004, 124, 301–317. [Google Scholar] [CrossRef] [Green Version]
- Neves, J.L.B.; Imperial, J.S.; Morgenstern, D.; Ueberheide, B.; Gajewiak, J.; Antunes, A.; Robinson, S.D.; Espino, S.; Watkins, M.; Vascoselos, V.; et al. Characterization of the first conotoxin from Conus ateralbus, a vermivorous cone snail from the Cabo Verde Archipielago. Mar. Drugs 2019, 17, 432. [Google Scholar] [CrossRef] [Green Version]
- Imperial, J.S.; Bansal, P.S.; Alewood, P.F.; Daly, N.L.; Craik, D.J.; Sporning, A.; Olivera, B.M. A novel conotoxin inhibitor of Kv1.6 channel and nAChR subtypes defines a new superfamily of conotoxins. Biochemistry 2006, 45, 8331–8340. [Google Scholar] [CrossRef] [PubMed]
- Imperial, J.; Chen, P.; Sporning, A.; Terlau, H.; Daly, N.; Craik, D.; Olivera, B.M. Tyrosine-rich conopeptides affect voltage-gated K+ channels. J. Biol. Chem. 2008, 283, 23026–23032. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Dendorfer, A.; Finol-Urdaneta, R.K.; Terlau, H.; Olivera, B.M. Biochemical Characterization of κM-RIIIJ, a Kv1.2 channel blocker: Evaluation of cardioprotective effects of kM-conotoxins. J. Biol. Chem. 2010, 285, 14882–14889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar, M.B.; Pérez-Reyes, L.; López, Z.; Heimer de la Cotera, E.; Falcón, A.; Ayala, C.; Galván, M.; Salvador, C.; Escobar, L. Peptide sr11a from Conus spurius is a novel peptide blocker for Kv1 potassium channels. Peptides 2010, 31, 1287–1291. [Google Scholar] [CrossRef] [PubMed]
- López-Vera, E.; Martínez-Hernández, L.; Aguilar, M.B.; Carrillo, E.; Gajewiak, J. Studies of conorfamide-Sr3 on human voltage-gated Kv1 potassium channel subtypes. Mar. Drugs 2020, 18, 425. [Google Scholar] [CrossRef] [PubMed]
- Terlau, H.; Shon, K.-J.; Grilley, M.; Stocker, M.; Stuhmer, W.; Olivera, B. Strategy for rapid immobilization of prey by a fish-hunting marine snail. Nature 1996, 381, 148–151. [Google Scholar] [CrossRef]
- Simons, C.; Rash, L.D.; Crawford, J.; Ma, L.; Cristofori-Armstrong, B.; Miller, D.; Ru, K.; Baillie, G.J.; Alanay, Y.; Jacquinet, A.; et al. Mutations in the voltage-gated potassium channel gene KCNH1 cause Temple-Baraitser syndrome and epilepsy. Nat. Genet. 2015, 47, 73–79. [Google Scholar] [CrossRef]
- Kortüm, F.; Caputo, V.; Bauer, C.K.; Stella, L.; Ciolfi, A.; Alawi, M.; Bocchinfuso, G.; Flex, E.; Paolacci, S.; Dentici, M.L.; et al. Mutations in KCNH1 and ATP6V1B2 cause Zimmermann-Laband syndrome. Nat. Genet. 2015, 47, 661–669. [Google Scholar] [CrossRef] [Green Version]
- Pardo, L.; Camino, D.; Sánchez, A.; Alves, F.; Brüggemann, A.; Beckh, S.; Stühmer, W. Oncogenic potential of EAG K+ channels. EMBO J. 1999, 18, 5540–5547. [Google Scholar] [CrossRef]
- Moreels, L.; Peigneur, S.; Yamaguchi, Y.; Vriens, K.; Waelkens, E.; Zhue, S.; Thevissen, K.; Cammue, B.; Sato, K.; Tytgat, J. Expanding the pharmacological profile of κ-hefutoxin 1 and analogues: A focus on the inhibitory effect on the oncogenic channel Kv10.1. Peptides 2017, 98, 43–50. [Google Scholar] [CrossRef]
- Peigneur, S.; Yamaguchi, Y.; Goto, H.; Srinivasan, K.N.; Gopalakrishnakone, P.; Tytgat, J.; Sato, K. Synthesis and characterization of amino acid deletion analogs of κ-hefutoxin 1, a scorpion toxin on potassium channels. Toxicon 2013, 71, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Moreels, L.; Peigneur, S.; Galan, T.D.; De Pauw, E.; Béress, L.; Waelkens, E.; Pardo, A.L.; Quinton, L.; Tytgat, J. APETx4, a novel sea anemone toxin and a modulator of the cancer-relevant potassium channel Kv10.1. Mar. Drugs 2017, 15, 287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Chin, Y.; Dekan, Z.; Herzig, V.; Chow, C.Y.; Heighway, J.; Lam, S.W.; Guillemin, G.; Alewood, P.; King, G. Novel venom-derived inhibitors of the human EAG channel, a putative antiepileptic drug target. Biochem. Pharmacol. 2018, 158, 60–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.D.; Higgins, T.J.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, positions-specific gap penalties and wight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Combet, C.; Blanchet, C.; Geourjon, G.; Deléage, G. NPS@: Network protein sequence analysis. Trends Biochem. Sci. 2000, 25, 147–150. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Hernández, L.; López-Vera, E.; Aguilar, M.B.; Rodriguez-Ruiz, X.C.; Ortíz-Arellano, M.A. κO-SrVIA Conopeptide, a Novel Inhibitor Peptide for Two Members of the Human EAG Potassium Channel Family. Int. J. Mol. Sci. 2023, 24, 11513. https://doi.org/10.3390/ijms241411513
Martínez-Hernández L, López-Vera E, Aguilar MB, Rodriguez-Ruiz XC, Ortíz-Arellano MA. κO-SrVIA Conopeptide, a Novel Inhibitor Peptide for Two Members of the Human EAG Potassium Channel Family. International Journal of Molecular Sciences. 2023; 24(14):11513. https://doi.org/10.3390/ijms241411513
Chicago/Turabian StyleMartínez-Hernández, Luis, Estuardo López-Vera, Manuel B. Aguilar, Ximena C. Rodriguez-Ruiz, and Mónica A. Ortíz-Arellano. 2023. "κO-SrVIA Conopeptide, a Novel Inhibitor Peptide for Two Members of the Human EAG Potassium Channel Family" International Journal of Molecular Sciences 24, no. 14: 11513. https://doi.org/10.3390/ijms241411513