

Clopidogrel Administration Impairs Post-Stroke Learning and Memory Recovery in Mice
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Short-Term Clopidogrel Administration Impaired Memory and Learning Post-Stroke
2.2. Short-Term Clopidogrel Administration Did Not Alter Motor Impairment
2.3. Short-Term Clopidogrel Administration Decreased Mouse Survival Rates and Body Weight
2.4. Short-Term Clopidogrel Administration Did Not Alter Vasculature Post-Stroke
2.5. Short-Term Clopidogrel Administration Increased Vascular Leakage Post-Stroke
2.6. Short-Term Clopidogrel Administration Changed the Microglia Number and Morphology Post-Stroke
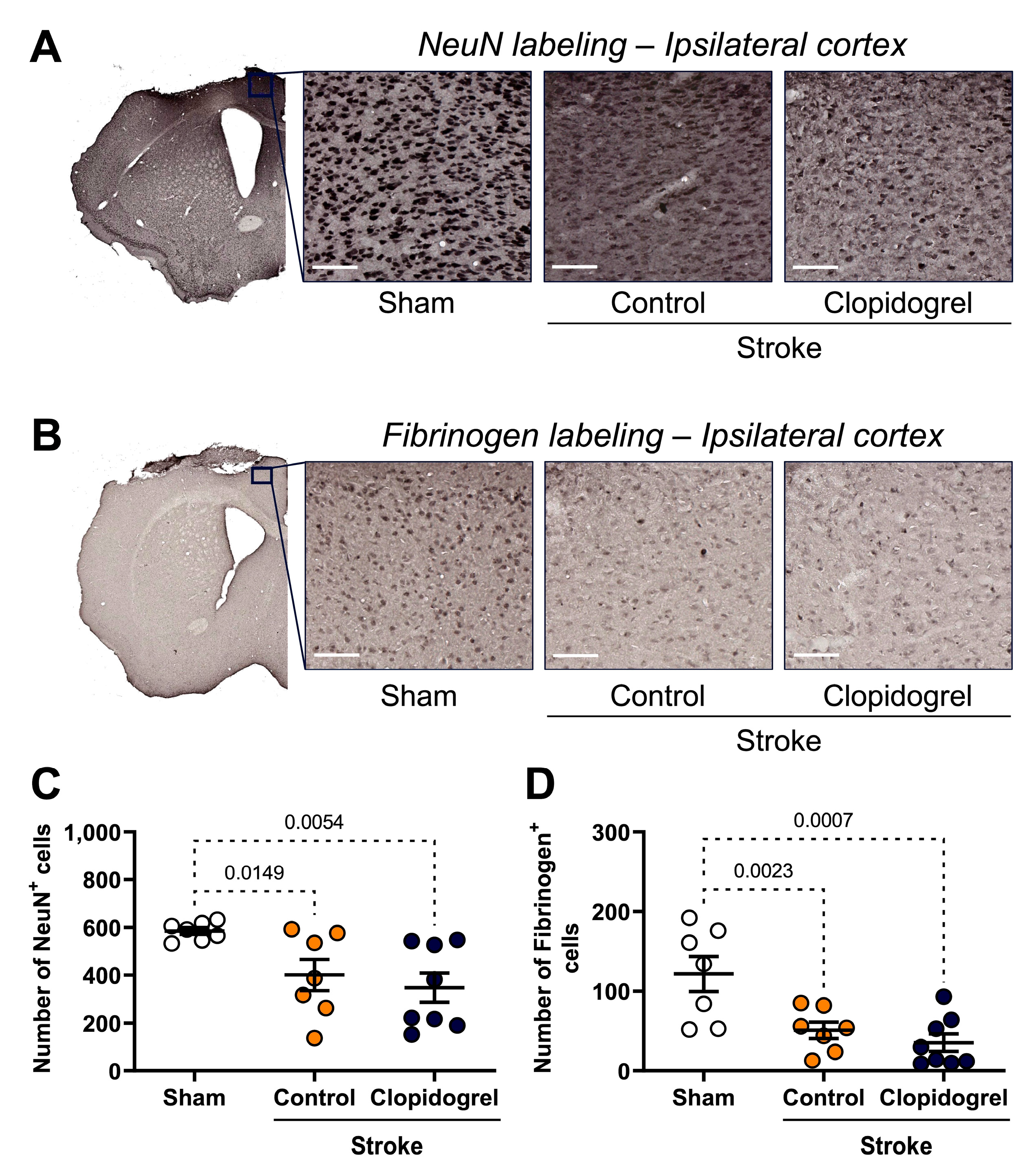
2.7. Short-Term Clopidogrel Administration Did Not Alter Neuronal Loss Post-Stroke
2.8. Short-Term Clopidogrel Administration Did Not Alter Fibrinogen Post-Stroke
2.9. Short-Term Clopidogrel Administration Decreased the Number of T Cells Post-Stroke
3. Discussion
4. Materials and Methods
4.1. Animals
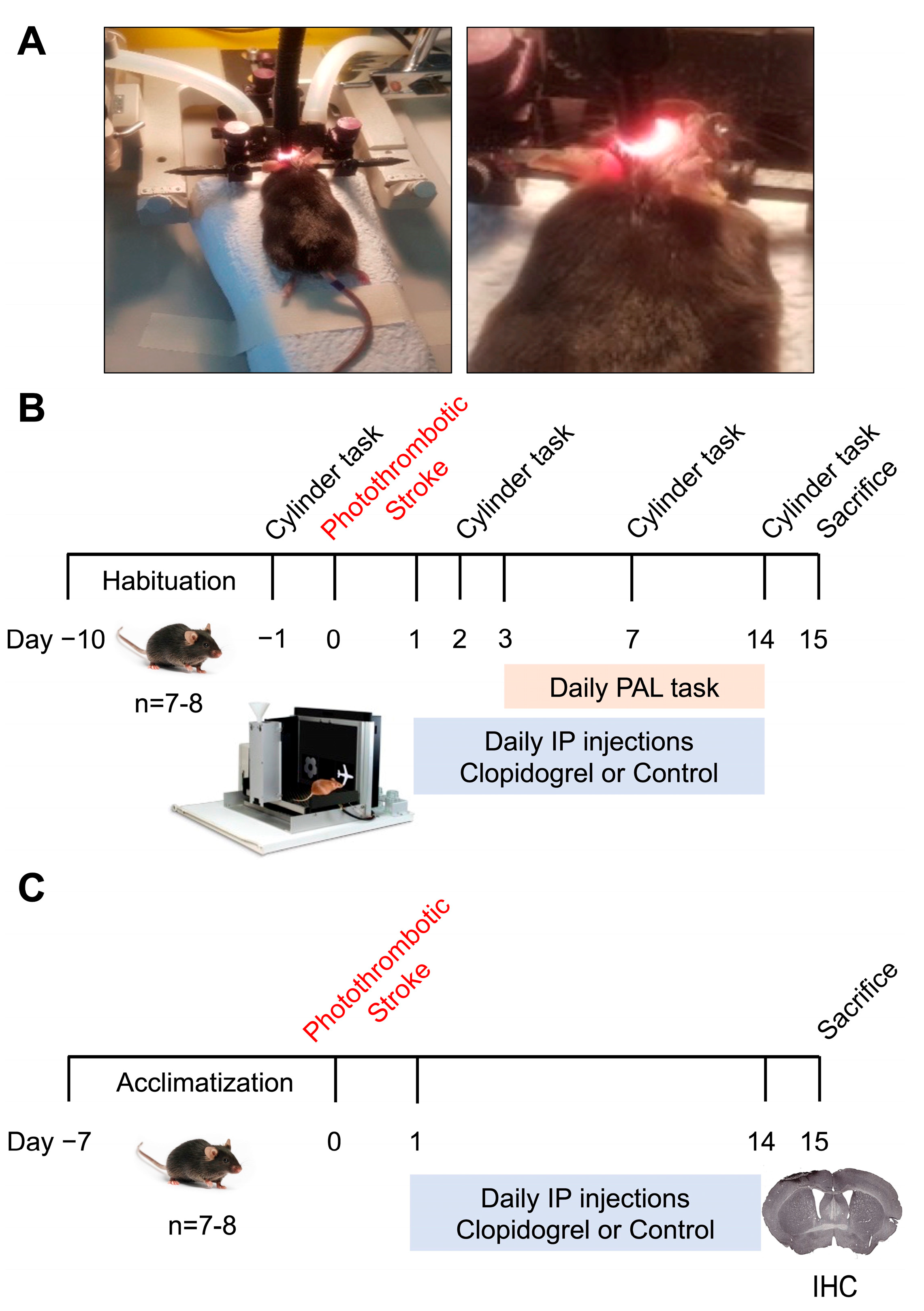
4.2. Experimental Design
4.3. Photothrombotic Occlusion
4.4. Clopidogrel Administration
4.5. Motor Testing
4.6. Cognitive Testing
4.7. Perfusion, Tissue Collection, and Tissue Processing
4.8. Immunohistochemistry
4.9. Image Acquisition and Analyses
4.10. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campbell, K.L.; Cohn, J.R.; Savage, M.P. Clopidogrel hypersensitivity: Clinical challenges and options for management. Expert Rev. Clin. Pharmacol. 2010, 3, 553–561. [Google Scholar] [CrossRef] [Green Version]
- Savi, P.; Labouret, C.; Delesque, N.; Guette, F.; Lupker, J.; Herbert, J.M. P2y(12), a new platelet ADP receptor, target of clopidogrel. Biochem. Biophys. Res. Commun. 2001, 283, 379–383. [Google Scholar] [CrossRef]
- Baqi, Y.; Muller, C.E. Antithrombotic P2Y12 receptor antagonists: Recent developments in drug discovery. Drug Discov. Today 2019, 24, 325–333. [Google Scholar] [CrossRef]
- Haynes, S.E.; Hollopeter, G.; Yang, G.; Kurpius, D.; Dailey, M.E.; Gan, W.B.; Julius, D. The P2Y12 receptor regulates microglial activation by extracellular nucleotides. Nat. Neurosci. 2006, 9, 1512–1519. [Google Scholar] [CrossRef]
- Hollopeter, G.; Jantzen, H.M.; Vincent, D.; Li, G.; England, L.; Ramakrishnan, V.; Yang, R.B.; Nurden, P.; Nurden, A.; Julius, D.; et al. Identification of the platelet ADP receptor targeted by antithrombotic drugs. Nature 2001, 409, 202–207. [Google Scholar] [CrossRef]
- Sasaki, Y.; Hoshi, M.; Akazawa, C.; Nakamura, Y.; Tsuzuki, H.; Inoue, K.; Kohsaka, S. Selective expression of Gi/o-coupled ATP receptor P2Y12 in microglia in rat brain. Glia 2003, 44, 242–250. [Google Scholar] [CrossRef]
- Aguzzi, A.; Barres, B.A.; Bennett, M.L. Microglia: Scapegoat, saboteur, or something else? Science 2013, 339, 156–161. [Google Scholar] [CrossRef] [Green Version]
- Lalancette-Hebert, M.; Gowing, G.; Simard, A.; Weng, Y.C.; Kriz, J. Selective ablation of proliferating microglial cells exacerbates ischemic injury in the brain. J. Neurosci. 2007, 27, 2596–2605. [Google Scholar] [CrossRef] [Green Version]
- Szalay, G.; Martinecz, B.; Lenart, N.; Kornyei, Z.; Orsolits, B.; Judak, L.; Csaszar, E.; Fekete, R.; West, B.L.; Katona, G.; et al. Microglia protect against brain injury and their selective elimination dysregulates neuronal network activity after stroke. Nat. Commun. 2016, 7, 11499. [Google Scholar] [CrossRef] [Green Version]
- Bolmont, T.; Haiss, F.; Eicke, D.; Radde, R.; Mathis, C.A.; Klunk, W.E.; Kohsaka, S.; Jucker, M.; Calhoun, M.E. Dynamics of the microglial/amyloid interaction indicate a role in plaque maintenance. J. Neurosci. 2008, 28, 4283–4292. [Google Scholar] [CrossRef] [Green Version]
- Davalos, D.; Grutzendler, J.; Yang, G.; Kim, J.V.; Zuo, Y.; Jung, S.; Littman, D.R.; Dustin, M.L.; Gan, W.B. ATP mediates rapid microglial response to local brain injury in vivo. Nat. Neurosci. 2005, 8, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Honda, S.; Sasaki, Y.; Ohsawa, K.; Imai, Y.; Nakamura, Y.; Inoue, K.; Kohsaka, S. Extracellular ATP or ADP induce chemotaxis of cultured microglia through Gi/o-coupled P2Y receptors. J. Neurosci. 2001, 21, 1975–1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohsawa, K.; Irino, Y.; Sanagi, T.; Nakamura, Y.; Suzuki, E.; Inoue, K.; Kohsaka, S. P2Y12 receptor-mediated integrin-beta1 activation regulates microglial process extension induced by ATP. Glia 2010, 58, 790–801. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, K.; Sloan, S.A.; Bennett, M.L.; Scholze, A.R.; O’Keeffe, S.; Phatnani, H.P.; Guarnieri, P.; Caneda, C.; Ruderisch, N.; et al. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J. Neurosci. 2014, 34, 11929–11947. [Google Scholar] [CrossRef] [Green Version]
- Eyo, U.B.; Gu, N.; De, S.; Dong, H.; Richardson, J.R.; Wu, L.J. Modulation of microglial process convergence toward neuronal dendrites by extracellular calcium. J. Neurosci. 2015, 35, 2417–2422. [Google Scholar] [CrossRef] [Green Version]
- Sipe, G.O.; Lowery, R.L.; Tremblay, M.E.; Kelly, E.A.; Lamantia, C.E.; Majewska, A.K. Microglial P2Y12 is necessary for synaptic plasticity in mouse visual cortex. Nat. Commun. 2016, 7, 10905. [Google Scholar] [CrossRef]
- Eyo, U.B.; Mo, M.; Yi, M.H.; Murugan, M.; Liu, J.; Yarlagadda, R.; Margolis, D.J.; Xu, P.; Wu, L.J. P2Y12R-Dependent Translocation Mechanisms Gate the Changing Microglial Landscape. Cell Rep. 2018, 23, 959–966. [Google Scholar] [CrossRef]
- Prakash, R.; Carmichael, S.T. Blood-brain barrier breakdown and neovascularization processes after stroke and traumatic brain injury. Curr. Opin. Neurol. 2015, 28, 556–564. [Google Scholar] [CrossRef] [Green Version]
- Sweeney, M.D.; Kisler, K.; Montagne, A.; Toga, A.W.; Zlokovic, B.V. The role of brain vasculature in neurodegenerative disorders. Nat. Neurosci. 2018, 21, 1318–1331. [Google Scholar] [CrossRef]
- Cserep, C.; Posfai, B.; Lenart, N.; Fekete, R.; Laszlo, Z.I.; Lele, Z.; Orsolits, B.; Molnar, G.; Heindl, S.; Schwarcz, A.D.; et al. Microglia monitor and protect neuronal function through specialized somatic purinergic junctions. Science 2020, 367, 528–537. [Google Scholar] [CrossRef]
- Lou, N.; Takano, T.; Pei, Y.; Xavier, A.L.; Goldman, S.A.; Nedergaard, M. Purinergic receptor P2RY12-dependent microglial closure of the injured blood-brain barrier. Proc. Natl. Acad. Sci. USA 2016, 113, 1074–1079. [Google Scholar] [CrossRef] [PubMed]
- Diener, H.C.; Sacco, R.L.; Yusuf, S.; Cotton, D.; Ounpuu, S.; Lawton, W.A.; Palesch, Y.; Martin, R.H.; Albers, G.W.; Bath, P.; et al. Effects of aspirin plus extended-release dipyridamole versus clopidogrel and telmisartan on disability and cognitive function after recurrent stroke in patients with ischaemic stroke in the PRoFESS trial. Lancet Neurol. 2008, 7, 875–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douiri, A.; McKevitt, C.; Emmett, E.S.; Rudd, A.G.; Wolfe, C.D. Long-term effects of secondary prevention on cognitive function in stroke patients. Circulation 2013, 128, 1341–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, W.Z.; Ong, L.K.; Kluge, M.G.; Gyawali, P.; Walker, F.R.; Nilsson, M. Similar cognitive deficits in mice and humans in the chronic phase post-stroke identified using the touchscreen-based paired-associate learning task. Sci. Rep. 2020, 10, 19545. [Google Scholar] [CrossRef]
- Golanov, E.V.; Sharpe, M.A.; Regnier-Golanov, A.S.; Del Zoppo, G.J.; Baskin, D.S.; Britz, G.W. Fibrinogen Chains Intrinsic to the Brain. Front. Neurosci. 2019, 13, 541. [Google Scholar] [CrossRef] [Green Version]
- Morris, R. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Sanchez-Bezanilla, S.; TeBay, C.; Nilsson, M.; Walker, F.R.; Ong, L.K. Visual discrimination impairment after experimental stroke is associated with disturbances in the polarization of the astrocytic aquaporin-4 and increased accumulation of neurotoxic proteins. Exp. Neurol. 2019, 318, 232–243. [Google Scholar] [CrossRef]
- Sanchez-Bezanilla, S.; Hood, R.J.; Collins-Praino, L.E.; Turner, R.J.; Walker, F.R.; Nilsson, M.; Ong, L.K. More than motor impairment: A spatiotemporal analysis of cognitive impairment and associated neuropathological changes following cortical photothrombotic stroke. J. Cereb. Blood Flow Metab. 2021, 41, 2439–2455. [Google Scholar] [CrossRef]
- Zalewska, K.; Pietrogrande, G.; Ong, L.K.; Abdolhoseini, M.; Kluge, M.; Johnson, S.J.; Walker, F.R.; Nilsson, M. Sustained administration of corticosterone at stress-like levels after stroke suppressed glial reactivity at sites of thalamic secondary neurodegeneration. Brain Behav. Immun. 2018, 69, 210–222. [Google Scholar] [CrossRef]
- Pietrogrande, G.; Zalewska, K.; Zhao, Z.; Johnson, S.J.; Nilsson, M.; Walker, F.R. Low Oxygen Post Conditioning as an Efficient Non-pharmacological Strategy to Promote Motor Function After Stroke. Transl. Stroke Res. 2019, 10, 402–412. [Google Scholar] [CrossRef]
- Coull, A.J.; Lovett, J.K.; Rothwell, P.M.; Oxford Vascular, S. Population based study of early risk of stroke after transient ischaemic attack or minor stroke: Implications for public education and organisation of services. BMJ 2004, 328, 326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giles, M.F.; Rothwell, P.M. Risk of stroke early after transient ischaemic attack: A systematic review and meta-analysis. Lancet Neurol. 2007, 6, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Johnston, S.C.; Gress, D.R.; Browner, W.S.; Sidney, S. Short-term prognosis after emergency department diagnosis of TIA. JAMA 2000, 284, 2901–2906. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.M.; McLaughlin, K.; Lorenzetti, D.L.; Hill, M.D.; Manns, B.J.; Ghali, W.A. Early risk of stroke after transient ischemic attack: A systematic review and meta-analysis. Arch. Intern. Med. 2007, 167, 2417–2422. [Google Scholar] [CrossRef] [Green Version]
- Antithrombotic Trialists, C. Collaborative meta-analysis of randomised trials of antiplatelet therapy for prevention of death, myocardial infarction, and stroke in high risk patients. BMJ 2002, 324, 71–86. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z. CAST: Randomised placebo-controlled trial of early aspirin use in 20,000 patients with acute ischaemic stroke. CAST (Chinese Acute Stroke Trial) Collaborative Group. Lancet 1997, 349, 1641–1649. [Google Scholar]
- Bowry, A.D.; Brookhart, M.A.; Choudhry, N.K. Meta-analysis of the efficacy and safety of clopidogrel plus aspirin as compared to antiplatelet monotherapy for the prevention of vascular events. Am. J. Cardiol. 2008, 101, 960–966. [Google Scholar] [CrossRef]
- Investigators, A.; Connolly, S.J.; Pogue, J.; Hart, R.G.; Hohnloser, S.H.; Pfeffer, M.; Chrolavicius, S.; Yusuf, S. Effect of clopidogrel added to aspirin in patients with atrial fibrillation. N. Engl. J. Med. 2009, 360, 2066–2078. [Google Scholar] [CrossRef]
- Johnston, S.C.; Easton, J.D.; Farrant, M.; Barsan, W.; Conwit, R.A.; Elm, J.J.; Kim, A.S.; Lindblad, A.S.; Palesch, Y.Y.; Clinical Research Collaboration, N.E.T.T.N.; et al. Clopidogrel and Aspirin in Acute Ischemic Stroke and High-Risk TIA. N. Engl. J. Med. 2018, 379, 215–225. [Google Scholar] [CrossRef]
- Tillman, H.; Johnston, S.C.; Farrant, M.; Barsan, W.; Elm, J.J.; Kim, A.S.; Lindblad, A.S.; Palesch, Y.Y.; Easton, J.D. Risk for Major Hemorrhages in Patients Receiving Clopidogrel and Aspirin Compared With Aspirin Alone After Transient Ischemic Attack or Minor Ischemic Stroke: A Secondary Analysis of the POINT Randomized Clinical Trial. JAMA Neurol. 2019, 76, 774–782. [Google Scholar] [CrossRef]
- Bhatt, D.L.; Flather, M.D.; Hacke, W.; Berger, P.B.; Black, H.R.; Boden, W.E.; Cacoub, P.; Cohen, E.A.; Creager, M.A.; Easton, J.D.; et al. Patients with prior myocardial infarction, stroke, or symptomatic peripheral arterial disease in the CHARISMA trial. J. Am. Coll. Cardiol. 2007, 49, 1982–1988. [Google Scholar] [CrossRef] [PubMed]
- Pöschl, E.; Schlötzer-Schrehardt, U.; Brachvogel, B.; Saito, K.; Ninomiya, Y.; Mayer, U. Collagen IV is essential for basement membrane stability but dispensable for initiation of its assembly during early development. Development 2004, 131, 1619–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalewska, K.; Ong, L.K.; Johnson, S.J.; Nilsson, M.; Walker, F.R. Oral administration of corticosterone at stress-like levels drives microglial but not vascular disturbances post-stroke. Neuroscience 2017, 352, 30–38. [Google Scholar] [CrossRef]
- Hamann, G.F.; Liebetrau, M.; Martens, H.; Burggraf, D.; Kloss, C.U.; Bültemeier, G.; Wunderlich, N.; Jäger, G.; Pfefferkorn, T. Microvascular basal lamina injury after experimental focal cerebral ischemia and reperfusion in the rat. J. Cereb. Blood Flow Metab. 2002, 22, 526–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trinkl, A.; Vosko, M.R.; Wunderlich, N.; Dichgans, M.; Hamann, G.F. Pravastatin reduces microvascular basal lamina damage following focal cerebral ischemia and reperfusion. Eur. J. Neurosci. 2006, 24, 520–526. [Google Scholar] [CrossRef] [PubMed]
- Vosko, M.R.; Busch, E.; Burggraf, D.; Bültemeier, G.; Hamann, G.F. Microvascular basal lamina damage in thromboembolic stroke in a rat model. Neurosci. Lett. 2003, 353, 217–220. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Hood, R.J.; Ong, L.K.; Pietrogrande, G.; Sanchez Bezanilla, S.; Warren, K.E.; Ilicic, M.; Kluge, M.G.; TeBay, C.; Ottersen, O.P.; et al. Exploring How Low Oxygen Post Conditioning Improves Stroke-Induced Cognitive Impairment: A Consideration of Amyloid-Beta Loading and Other Mechanisms. Front. Neurol. 2021, 12, 585189. [Google Scholar] [CrossRef] [PubMed]
- Pereillo, J.M.; Maftouh, M.; Andrieu, A.; Uzabiaga, M.F.; Fedeli, O.; Savi, P.; Pascal, M.; Herbert, J.M.; Maffrand, J.P.; Picard, C. Structure and stereochemistry of the active metabolite of clopidogrel. Drug Metab. Dispos. Biol. Fate Chem. 2002, 30, 1288–1295. [Google Scholar] [CrossRef]
- Ito, D.; Tanaka, K.; Suzuki, S.; Dembo, T.; Fukuuchi, Y. Enhanced expression of Iba1, ionized calcium-binding adapter molecule 1, after transient focal cerebral ischemia in rat brain. Stroke 2001, 32, 1208–1215. [Google Scholar] [CrossRef] [Green Version]
- Taubert, D.; Kastrati, A.; Harlfinger, S.; Gorchakova, O.; Lazar, A.; von Beckerath, N.; Schomig, A.; Schomig, E. Pharmacokinetics of clopidogrel after administration of a high loading dose. Thromb. Haemost. 2004, 92, 311–316. [Google Scholar] [CrossRef] [Green Version]
- Webster, C.M.; Hokari, M.; McManus, A.; Tang, X.N.; Ma, H.; Kacimi, R.; Yenari, M.A. Microglial P2Y12 deficiency/inhibition protects against brain ischemia. PLoS ONE 2013, 8, e70927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelosa, P.; Lecca, D.; Fumagalli, M.; Wypych, D.; Pignieri, A.; Cimino, M.; Verderio, C.; Enerback, M.; Nikookhesal, E.; Tremoli, E.; et al. Microglia is a key player in the reduction of stroke damage promoted by the new antithrombotic agent ticagrelor. J. Cereb. Blood Flow Metab. 2014, 34, 979–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, W.; Liu, X.; Liu, W.; Wang, C.; Zhao, S.; Wen, S.; Gong, T.; Chen, W.; Chen, Q.; Ye, W.; et al. Dual antiplatelet therapy improves functional recovery and inhibits inflammation after cerebral ischemia/reperfusion injury. Ann. Transl. Med. 2022, 10, 283. [Google Scholar] [CrossRef] [PubMed]
- Neher, J.J.; Neniskyte, U.; Zhao, J.W.; Bal-Price, A.; Tolkovsky, A.M.; Brown, G.C. Inhibition of microglial phagocytosis is sufficient to prevent inflammatory neuronal death. J. Immunol. 2011, 186, 4973–4983. [Google Scholar] [CrossRef] [Green Version]
- Gill, D.; Veltkamp, R. Dynamics of T cell responses after stroke. Curr. Opin. Pharmacol. 2016, 26, 26–32. [Google Scholar] [CrossRef]
- Kluge, M.G.; Abdolhoseini, M.; Zalewska, K.; Ong, L.K.; Johnson, S.J.; Nilsson, M.; Walker, F.R. Spatiotemporal analysis of impaired microglia process movement at sites of secondary neurodegeneration post-stroke. J. Cereb. Blood Flow Metab. 2019, 39, 2456–2470. [Google Scholar] [CrossRef]
- Kluge, M.G.; Kracht, L.; Abdolhoseini, M.; Ong, L.K.; Johnson, S.J.; Nilsson, M.; Walker, F.R. Impaired microglia process dynamics post-stroke are specific to sites of secondary neurodegeneration. Glia 2017, 65, 1885–1899. [Google Scholar] [CrossRef]
- Ong, L.K.; Chow, W.Z.; TeBay, C.; Kluge, M.; Pietrogrande, G.; Zalewska, K.; Crock, P.; Åberg, N.D.; Bivard, A.; Johnson, S.J.; et al. Growth Hormone Improves Cognitive Function After Experimental Stroke. Stroke 2018, 49, 1257–1266. [Google Scholar] [CrossRef]
- Sanchez-Bezanilla, S.; Aberg, N.D.; Crock, P.; Walker, F.R.; Nilsson, M.; Isgaard, J.; Ong, L.K. Growth Hormone Promotes Motor Function after Experimental Stroke and Enhances Recovery-Promoting Mechanisms within the Peri-Infarct Area. Int. J. Mol. Sci. 2020, 21, 606. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Ong, L.K.; Johnson, S.; Nilsson, M.; Walker, F.R. Chronic stress induced disruption of the peri-infarct neurovascular unit following experimentally induced photothrombotic stroke. J. Cereb. Blood Flow Metab. 2017, 37, 3709–3724. [Google Scholar] [CrossRef] [Green Version]
- Franklin, K.B.J.; Paxinos, G. Paxinos and Franklin’s the Mouse Brain in Stereotaxic Coordinates, 4th ed.; Academic Press: Cambridge, MA, USA, 2013; an imprint of Elsevier: Amsterdam, The Netherlands; 2019; p. 360. [Google Scholar]
- Tynan, R.J.; Naicker, S.; Hinwood, M.; Nalivaiko, E.; Buller, K.M.; Pow, D.V.; Day, T.A.; Walker, F.R. Chronic stress alters the density and morphology of microglia in a subset of stress-responsive brain regions. Brain Behav. Immun. 2010, 24, 1058–1068. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.J.; Walker, F.R. Strategies to improve quantitative assessment of immunohistochemical and immunofluorescent labelling. Sci. Rep. 2015, 5, 10607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belaya, I.; Ivanova, M.; Sorvari, A.; Ilicic, M.; Loppi, S.; Koivisto, H.; Varricchio, A.; Tikkanen, H.; Walker, F.R.; Atalay, M.; et al. Astrocyte remodeling in the beneficial effects of long-term voluntary exercise in Alzheimer’s disease. J. Neuroinflamm. 2020, 17, 271. [Google Scholar] [CrossRef]
- Abdolhoseini, M.; Kluge, M.G.; Walker, F.R.; Johnson, S.J. Segmentation, Tracing, and Quantification of Microglial Cells from 3D Image Stacks. Sci. Rep. 2019, 9, 8557. [Google Scholar] [CrossRef] [Green Version]
- Abdolhoseini, M.; Kluge, M.G.; Walker, F.R.; Johnson, S.J. Segmentation of Heavily Clustered Nuclei from Histopathological Images. Sci. Rep. 2019, 9, 4551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdolhoseini, M.; Walker, F.; Johnson, S. Automated tracing of microglia using multilevel thresholding and minimum spanning trees. In Proceedings of the 2016 38th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Orlando, FL, USA, 16–20 August 2016; pp. 1208–1211. [Google Scholar]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paul, M.; Paul, J.W.; Hinwood, M.; Hood, R.J.; Martin, K.; Abdolhoseini, M.; Johnson, S.J.; Pollack, M.; Nilsson, M.; Walker, F.R. Clopidogrel Administration Impairs Post-Stroke Learning and Memory Recovery in Mice. Int. J. Mol. Sci. 2023, 24, 11706. https://doi.org/10.3390/ijms241411706
Paul M, Paul JW, Hinwood M, Hood RJ, Martin K, Abdolhoseini M, Johnson SJ, Pollack M, Nilsson M, Walker FR. Clopidogrel Administration Impairs Post-Stroke Learning and Memory Recovery in Mice. International Journal of Molecular Sciences. 2023; 24(14):11706. https://doi.org/10.3390/ijms241411706
Chicago/Turabian StylePaul, Marina, Jonathan W. Paul, Madeleine Hinwood, Rebecca J. Hood, Kristy Martin, Mahmoud Abdolhoseini, Sarah J. Johnson, Michael Pollack, Michael Nilsson, and Frederick R. Walker. 2023. "Clopidogrel Administration Impairs Post-Stroke Learning and Memory Recovery in Mice" International Journal of Molecular Sciences 24, no. 14: 11706. https://doi.org/10.3390/ijms241411706
APA StylePaul, M., Paul, J. W., Hinwood, M., Hood, R. J., Martin, K., Abdolhoseini, M., Johnson, S. J., Pollack, M., Nilsson, M., & Walker, F. R. (2023). Clopidogrel Administration Impairs Post-Stroke Learning and Memory Recovery in Mice. International Journal of Molecular Sciences, 24(14), 11706. https://doi.org/10.3390/ijms241411706