

Nutritional Provision of Iron Complexes by the Major Allergen Alt a 1 to Human Immune Cells Decreases Its Presentation
 , , , , , ,
, , , , , ,  ,
,  , ,
, , (This article belongs to the Section Molecular Biology)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
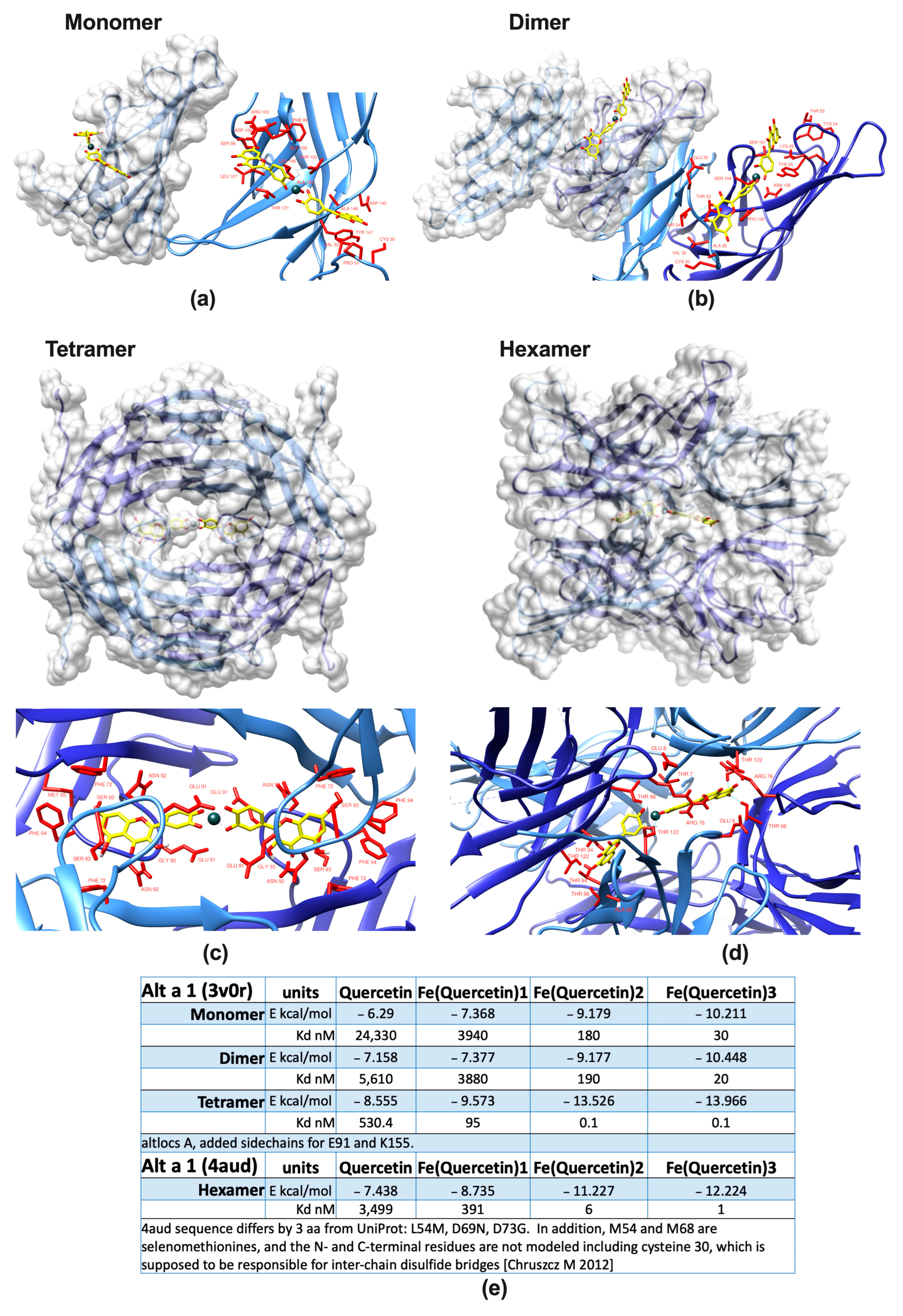
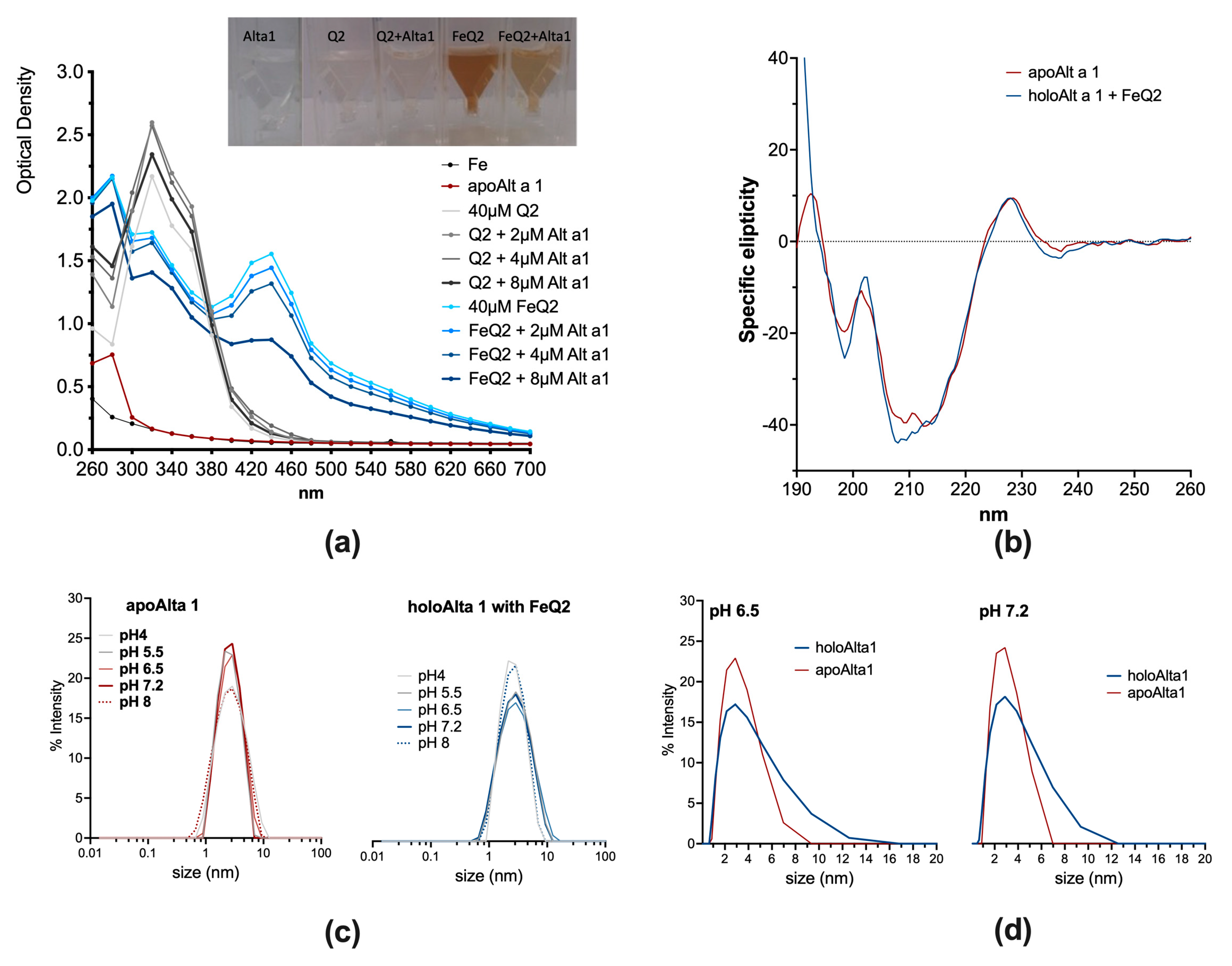
2.1. Alt a 1 Binds with Very High Affinity to Iron–Quercetin Complexes In Silico and In Vitro
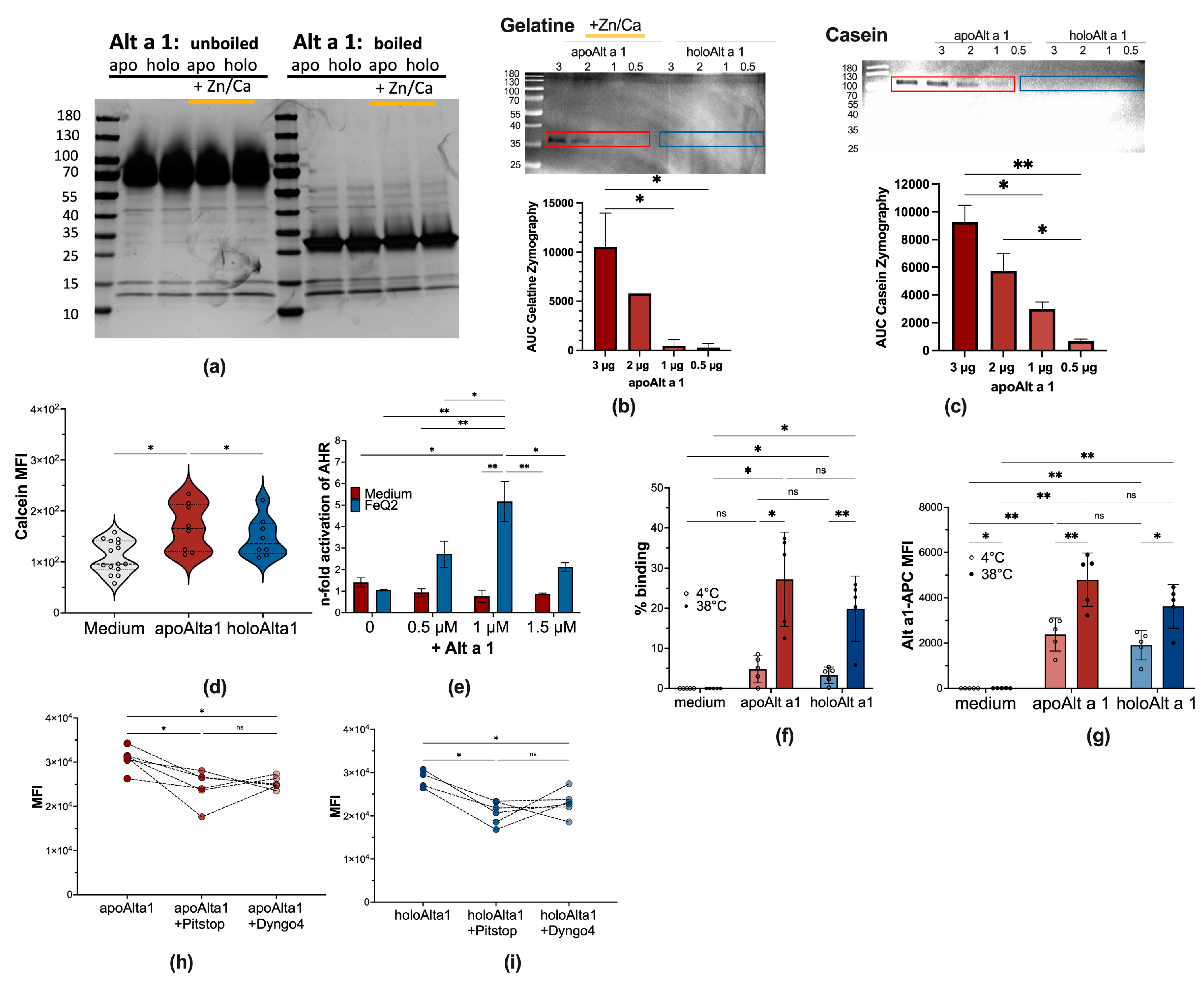
2.2. Alt a 1: Enzymatic Activity, Transport Function, and Clathrin-Dependent Uptake
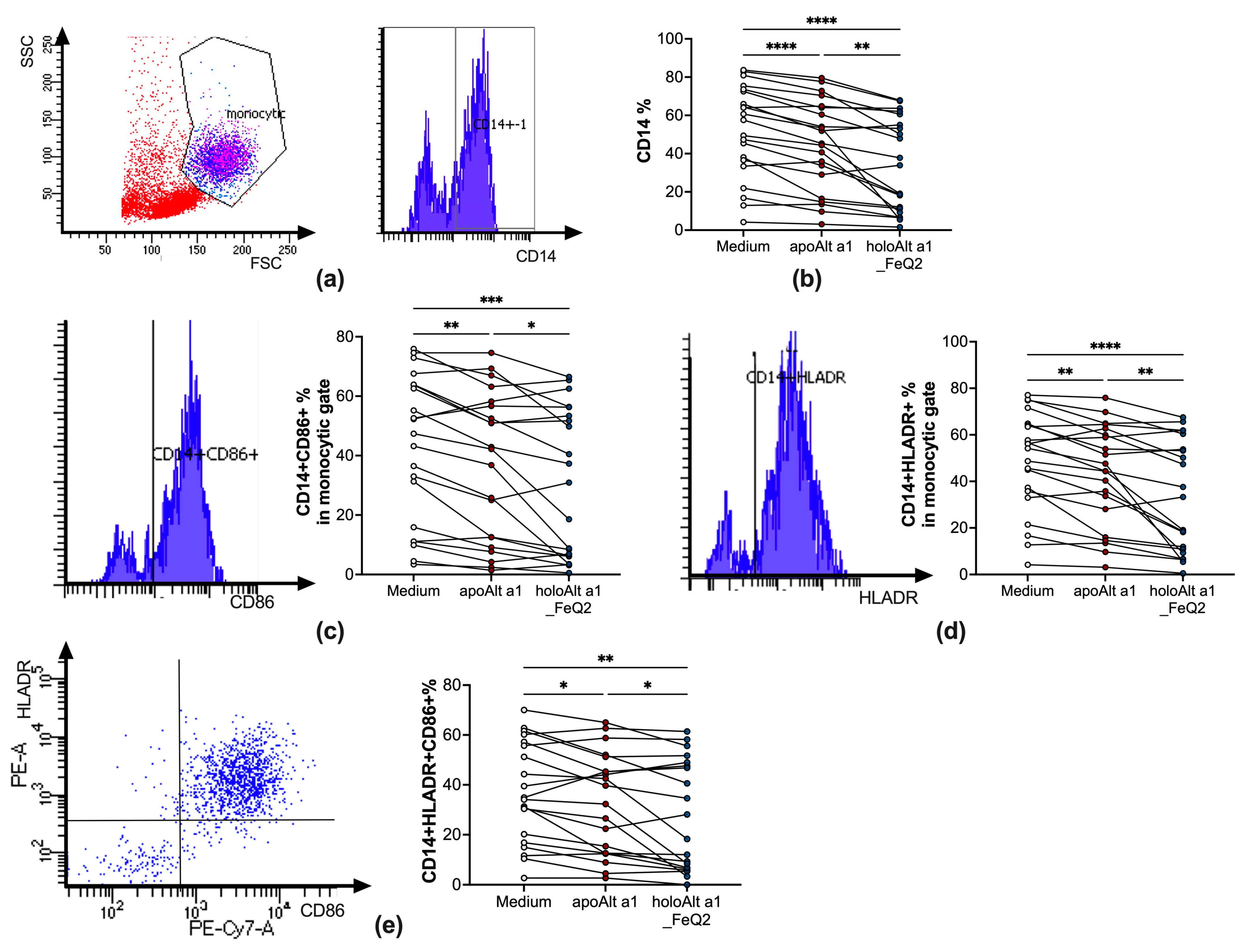
2.3. Decreased Antigen Presentation upon Incubation with Alt a 1 Binding to Iron–Quercetin Complexes
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. In Silico Structural and Docking Analysis
4.3. Alt a 1 Expression in Pichia Pastoris
4.4. Generation of apo– and holoAlt a 1
4.5. Spectral Analysis of Alt a 1 with Iron–Quercetin Complexes
4.6. Silverstain, Casein, and Gelatine-Zymography with apo–/holoAlt a 1
4.7. Endocytosis Experiments with Thp-1 Cells and Labile Iron Measurements
4.8. AhR Reporter Assay
4.9. Flow Cytometric Analysis of Human PBMCs
4.10. Cytokine Analysis of PBMCs Supernatant
4.11. Statistical Analysis
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lindblom, S.D.; Wangeline, A.L.; Barillas, J.R.V.; Devilbiss, B.; Fakra, S.C.; Pilon-Smits, E.A.H. Fungal Endophyte Alternaria tenuissima Can Affect Growth and Selenium Accumulation in Its Hyperaccumulator Host Astragalus bisulcatus. Front. Plant Sci. 2018, 9, 1213. [Google Scholar] [CrossRef] [PubMed]
- DeMers, M. Alternaria alternata as endophyte and pathogen. Microbiology 2022, 168. [Google Scholar] [CrossRef]
- Kumari, V.V.; Banerjee, P.; Verma, V.C.; Sukumaran, S.; Chandran, M.A.S.; Gopinath, K.A.; Venkatesh, G.; Yadav, S.K.; Singh, V.K.; Awasthi, N.K. Plant Nutrition: An Effective Way to Alleviate Abiotic Stress in Agricultural Crops. Int. J. Mol. Sci. 2022, 23, 8519. [Google Scholar] [CrossRef] [PubMed]
- Vicki, A.M.; Daniel, J.W. Alternaria: A Sinonasal Pathogen of Immunocompromised Hosts. Clin. Infect. Dis. 1993, 16, 265–270. [Google Scholar]
- Yu, H. Studies on fungi of the normal skin. Hifuka kiyo. Acta Dermatol. 1965, 60, 126–174. [Google Scholar]
- Pastor, F.J.; Guarro, J. Alternaria infections: Laboratory diagnosis and relevant clinical features. Clin. Microbiol. Infect. 2008, 14, 734–746. [Google Scholar] [CrossRef] [PubMed]
- Nema, H.V.; Ahuja, O.P.; Bal, A.; Mohapatra, L.N. Mycotic Flora of the Conjunctiva. Am. J. Ophthalmol. 1966, 62, 968–970. [Google Scholar] [CrossRef] [PubMed]
- Lo Porto, D.; Cona, A.; Todaro, F.; De Carolis, E.; Cardinale, F.; Hafeez, N.; Di Martino, G.; Conaldi, P.G.; Sanguinetti, M.; Grossi, P.A.; et al. Phaeohyphomycosis in Solid Organ Transplant Recipients: A Case Series and Narrative Review of the Literature. J. Fungi 2023, 9, 283. [Google Scholar] [CrossRef]
- Lee, J.Y.; Hyun, M.; Kim, H.A.; Ryu, S.Y. Unusual Presentation of Subcutaneous Phaeohyphomycosis by Alternaria alternate. Ann. Dermatol. 2019, 31, 563–566. [Google Scholar] [CrossRef]
- Hattab, Z.; Ben Lasfar, N.; Abid, M.; Bellazreg, F.; Fathallah, A.; Hachfi, W.; Letaief, A. Alternaria alternata infection causing rhinosinusitis and orbital involvement in an immunocompetent patient. New Microbes New Infect. 2019, 32, 100561. [Google Scholar] [CrossRef]
- Sanchez, P.; Velez-Del-Burgo, A.; Sunen, E.; Martinez, J.; Postigo, I. Fungal Allergen and Mold Allergy Diagnosis: Role and Relevance of Alternaria alternata Alt a 1 Protein Family. J. Fungi 2022, 8, 277. [Google Scholar] [CrossRef] [PubMed]
- Kespohl, S.; Raulf, M. Mold Sensitization in Asthmatic and Non-asthmatic Subjects Diagnosed with Extract-Based Versus Component-Based Allergens. Adv. Exp. Med. Biol. 2019, 1153, 79–89. [Google Scholar] [CrossRef]
- Chruszcz, M.; Chapman, M.D.; Osinski, T.; Solberg, R.; Demas, M.; Porebski, P.J.; Majorek, K.A.; Pomes, A.; Minor, W. Alternaria alternata allergen Alt a 1: A unique beta-barrel protein dimer found exclusively in fungi. J. Allergy Clin. Immunol. 2012, 130, 241–247.e249. [Google Scholar] [CrossRef]
- Garrido-Arandia, M.; Silva-Navas, J.; Ramirez-Castillejo, C.; Cubells-Baeza, N.; Gomez-Casado, C.; Barber, D.; Pozo, J.C.; Melendi, P.G.; Pacios, L.F.; Diaz-Perales, A. Characterisation of a flavonoid ligand of the fungal protein Alt a 1. Sci. Rep. 2016, 6, 33468. [Google Scholar] [CrossRef] [PubMed]
- Mechaly, A.E.; Ibanez de Opakua, A.; Asturia, J.; Viguera, A.R. Crystal Structure of Alternaria alternata Major Allergen Alt a 1. Available online: https://www.wwpdb.org/pdb?id=pdb_00004aud (accessed on 21 November 2021).
- Garrido-Arandia, M.; Bretones, J.; Gomez-Casado, C.; Cubells, N.; Diaz-Perales, A.; Pacios, L.F. Computational study of pH-dependent oligomerization and ligand binding in Alt a 1, a highly allergenic protein with a unique fold. J. Comput. Aided Mol. Des. 2016, 30, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Regner, A.; Szepannek, N.; Wiederstein, M.; Fakhimahmadi, A.; Paciosis, L.F.; Blokhuis, B.R.; Redegeld, F.A.; Hofstetter, G.; Dvorak, Z.; Jensen-Jarolim, E.; et al. Binding to Iron Quercetin Complexes Increases the Antioxidant Capacity of the Major Birch Pollen Allergen Bet v 1 and Reduces Its Allergenicity. Antioxidants 2022, 12, 42. [Google Scholar] [CrossRef] [PubMed]
- Afify, S.M.; Regner, A.; Pacios, L.F.; Blokhuis, B.R.; Jensen, S.A.; Redegeld, F.A.; Pali-Scholl, I.; Hufnagl, K.; Bianchini, R.; Guethoff, S.; et al. Micronutritional supplementation with a holoBLG-based FSMP (food for special medical purposes)-lozenge alleviates allergic symptoms in BALB/c mice: Imitating the protective farm effect. Clin. Exp. Allergy 2021, 52, 426–441. [Google Scholar] [CrossRef]
- Afify, S.M.; Pali-Scholl, I.; Hufnagl, K.; Hofstetter, G.; El-Bassuoni, M.A.-R.; Roth-Walter, F.; Jensen-Jarolim, E. Bovine Holo-Beta-Lactoglobulin Cross-Protects Against Pollen Allergies in an Innate Manner in BALB/c Mice: Potential Model for the Farm Effect. Front. Immunol. 2021, 12, 176. [Google Scholar] [CrossRef]
- Roth-Walter, F. Iron-Deficiency in Atopic Diseases: Innate Immune Priming by Allergens and Siderophores. Front. Allergy 2022, 3, 859922. [Google Scholar] [CrossRef]
- Roth-Walter, F.; Afify, S.M.; Pacios, L.F.; Blokhuis, B.R.; Redegeld, F.; Regner, A.; Petje, L.M.; Fiocchi, A.; Untersmayr, E.; Dvorak, Z.; et al. Cow’s milk protein beta-lactoglobulin confers resilience against allergy by targeting complexed iron into immune cells. J. Allergy Clin. Immunol. 2021, 147, 321–334.e324. [Google Scholar] [CrossRef]
- Roth-Walter, F.; Pacios, L.F.; Gomez-Casado, C.; Hofstetter, G.; Roth, G.A.; Singer, J.; Diaz-Perales, A.; Jensen-Jarolim, E. The major cow milk allergen Bos d 5 manipulates T-helper cells depending on its load with siderophore-bound iron. PLoS ONE 2014, 9, e104803. [Google Scholar] [CrossRef] [PubMed]
- Luscher, A.; Gasser, V.; Bumann, D.; Mislin, G.L.A.; Schalk, I.J.; Kohler, T. Plant-Derived Catechols Are Substrates of TonB-Dependent Transporters and Sensitize Pseudomonas aeruginosa to Siderophore-Drug Conjugates. mBio 2022, 13, e0149822. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Xu, X.; Xia, L.; Xia, C.; Tang, J.; Ouyang, Z. Quercetin-Iron Complex: Synthesis, Characterization, Antioxidant, DNA Binding, DNA Cleavage, and Antibacterial Activity Studies. J. Fluoresc. 2016, 26, 2023–2031. [Google Scholar] [CrossRef]
- Symonowicz, M.; Sykula-Zajac, A.; Lodyga-Chruscinska, E.; Rumora, I.; Straukas, M. Evaluation of polyphenols and anthocyanins contents in black chockeberry—Photinia melanocarpa (Michx.) fruits extract. Acta Pol. Pharm. 2012, 69, 381–387. [Google Scholar] [PubMed]
- El Hajji, H.; Nkhili, E.; Tomao, V.; Dangles, O. Interactions of quercetin with iron and copper ions: Complexation and autoxidation. Free Radic. Res. 2006, 40, 303–320. [Google Scholar] [CrossRef]
- Perron, N.R.; Brumaghim, J.L. A review of the antioxidant mechanisms of polyphenol compounds related to iron binding. Cell Biochem. Biophys. 2009, 53, 75–100. [Google Scholar] [CrossRef]
- Raymond, K.N.; Dertz, E.A.; Kim, S.S. Enterobactin: An archetype for microbial iron transport. Proc. Natl. Acad. Sci. USA 2003, 100, 3584–3588. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Luo, G.; Tang, Y.; Yao, P. Quercetin and iron metabolism: What we know and what we need to know. Food Chem. Toxicol. 2018, 114, 190–203. [Google Scholar] [CrossRef]
- Wang, X.; Li, Y.; Han, L.; Li, J.; Liu, C.; Sun, C. Role of Flavonoids in the Treatment of Iron Overload. Front. Cell Dev. Biol. 2021, 9, 685364. [Google Scholar] [CrossRef]
- Yin, M.; Liu, Y.; Chen, Y. Iron metabolism: An emerging therapeutic target underlying the anti-cancer effect of quercetin. Free Radic. Res. 2021, 55, 296–303. [Google Scholar] [CrossRef]
- Lesjak, M.; Hoque, R.; Balesaria, S.; Skinner, V.; Debnam, E.S.; Srai, S.K.; Sharp, P.A. Quercetin inhibits intestinal iron absorption and ferroportin transporter expression in vivo and in vitro. PLoS ONE 2014, 9, e102900. [Google Scholar] [CrossRef] [PubMed]
- Bardestani, A.; Ebrahimpour, S.; Esmaeili, A.; Esmaeili, A. Quercetin attenuates neurotoxicity induced by iron oxide nanoparticles. J. Nanobiotechnol. 2021, 19, 327. [Google Scholar] [CrossRef] [PubMed]
- Sajadi Hezaveh, Z.; Azarkeivan, A.; Janani, L.; Hosseini, S.; Shidfar, F. The effect of quercetin on iron overload and inflammation in beta-thalassemia major patients: A double-blind randomized clinical trial. Complement. Ther. Med. 2019, 46, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Roth-Walter, F.; Gomez-Casado, C.; Jensen-Jarolim, E.; Diaz Perales, A.; Pacios, L.F.; Singer, J. Method and Means for Diagnosing and Treating Allergy Using Lipocalin Levels. U.S. Patent No. 10,914,744, 7 January 2015. [Google Scholar]
- Roth-Walter, F.; Gomez-Casado, C.; Pacios, L.F.; Mothes-Luksch, N.; Roth, G.A.; Singer, J.; Diaz-Perales, A.; Jensen-Jarolim, E. Bet v 1 from birch pollen is a lipocalin-like protein acting as allergen only when devoid of iron by promoting Th2 lymphocytes. J. Biol. Chem. 2014, 289, 17416–17421. [Google Scholar] [CrossRef]
- Roth-Walter, F.; Schmutz, R.; Mothes-Luksch, N.; Lemell, P.; Zieglmayer, P.; Zieglmayer, R.; Jensen-Jarolim, E. Clinical efficacy of sublingual immunotherapy is associated with restoration of steady-state serum lipocalin 2 after SLIT: A pilot study. World Allergy Organ. J. 2018, 11, 21. [Google Scholar] [CrossRef]
- Stetefeld, J.; McKenna, S.A.; Patel, T.R. Dynamic light scattering: A practical guide and applications in biomedical sciences. Biophys. Rev. 2016, 8, 409–427. [Google Scholar] [CrossRef]
- Tenopoulou, M.; Kurz, T.; Doulias, P.T.; Galaris, D.; Brunk, U.T. Does the calcein-AM method assay the total cellular ‘labile iron pool’ or only a fraction of it? Biochem. J. 2007, 403, 261–266. [Google Scholar] [CrossRef]
- Novotna, A.; Pavek, P.; Dvorak, Z. Novel Stably Transfected Gene Reporter Human Hepatoma Cell Line for Assessment of Aryl Hydrocarbon Receptor Transcriptional Activity: Construction and Characterization. Environ. Sci. Technol. 2011, 45, 10133–10139. [Google Scholar] [CrossRef] [PubMed]
- Elkhatib, N.; Porshneva, K.; Montagnac, G. Migration cues interpretation by clathrin-coated structures. Curr. Opin. Cell Biol. 2021, 72, 100–105. [Google Scholar] [CrossRef]
- Wardlaw, A.J.; Rick, E.M.; Pur Ozyigit, L.; Scadding, A.; Gaillard, E.A.; Pashley, C.H. New Perspectives in the Diagnosis and Management of Allergic Fungal Airway Disease. J. Asthma Allergy 2021, 14, 557–573. [Google Scholar] [CrossRef]
- Diaz, P.I.; Hong, B.-Y.; Dupuy, A.K.; Strausbaugh, L.D. Mining the oral mycobiome: Methods, components, and meaning. Virulence 2017, 8, 313–323. [Google Scholar] [CrossRef]
- Santus, W.; Devlin, J.R.; Behnsen, J. Crossing Kingdoms: How the Mycobiota and Fungal-Bacterial Interactions Impact Host Health and Disease. Infect. Immun. 2021, 89, 1110–1128. [Google Scholar] [CrossRef]
- Gallelli, B.; Viviani, M.; Nebuloni, M.; Marzano, A.V.; Pozzi, C.; Messa, P.; Fogazzi, G.B. Skin infection due to Alternaria species in kidney allograft recipients: Report of a new case and review of the literature. J. Nephrol. 2006, 19, 668–672. [Google Scholar]
- Vermeire, S.E.; de Jonge, H.; Lagrou, K.; Kuypers, D.R. Cutaneous phaeohyphomycosis in renal allograft recipients: Report of 2 cases and review of the literature. Diagn. Microbiol. Infect. Dis. 2010, 68, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Jiang, Y.; Wang, D.; Zheng, M.; Liu, X.; Yuan, C. Enhancement in serum (1-3)-beta-D-glucan level by cutaneous alternariosis: A case report and literature review. Microb. Pathog. 2021, 150, 104703. [Google Scholar] [CrossRef]
- Gabriel, M.F.; Postigo, I.; Tomaz, C.T.; Martinez, J. Alternaria alternata allergens: Markers of exposure, phylogeny and risk of fungi-induced respiratory allergy. Environ. Int. 2016, 89–90, 71–80. [Google Scholar] [CrossRef]
- Vailes, L.D.; Perzanowski, M.S.; Wheatley, L.M.; Platts-Mills, T.A.; Chapman, M.D. IgE and IgG antibody responses to recombinant Alt a 1 as a marker of sensitization to Alternaria in asthma and atopic dermatitis. Clin. Exp. Allergy 2001, 31, 1891–1895. [Google Scholar] [CrossRef] [PubMed]
- Lopes, L.; Borges-Costa, J.; Soares-Almeida, L.; Filipe, P.; Neves, F.; Santana, A.; Guerra, J.; Kutzner, H. Cutaneous Alternariosis Caused by Alternaria infectoria: Three Cases in Kidney Transplant Patients. Healthcare 2013, 1, 100–106. [Google Scholar] [CrossRef]
- Kejik, Z.; Kaplanek, R.; Masarik, M.; Babula, P.; Matkowski, A.; Filipensky, P.; Vesela, K.; Gburek, J.; Sykora, D.; Martasek, P.; et al. Iron Complexes of Flavonoids-Antioxidant Capacity and Beyond. Int. J. Mol. Sci. 2021, 22, 646. [Google Scholar] [CrossRef] [PubMed]
- Chobot, V.; Hadacek, F. Iron and its complexation by phenolic cellular metabolites: From oxidative stress to chemical weapons. Plant Signal. Behav. 2010, 5, 4–8. [Google Scholar] [CrossRef]
- von Loetzen, C.S.; Hoffmann, T.; Hartl, M.J.; Schweimer, K.; Schwab, W.; Rosch, P.; Hartl-Spiegelhauer, O. Secret of the major birch pollen allergen Bet v 1: Identification of the physiological ligand. Biochem. J. 2014, 457, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Jacob, T.; von Loetzen, C.S.; Reuter, A.; Lacher, U.; Schiller, D.; Schobert, R.; Mahler, V.; Vieths, S.; Rosch, P.; Schweimer, K.; et al. Identification of a natural ligand of the hazel allergen Cor a 1. Sci. Rep. 2019, 9, 8714. [Google Scholar] [CrossRef] [PubMed]
- Casanal, A.; Zander, U.; Dupeux, F.; Valpuesta, V.; Marquez, J.A. Purification, crystallization and preliminary X-ray analysis of the strawberry allergens Fra a 1E and Fra a 3 in the presence of catechin. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2013, 69, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Vesic, J.; Stambolic, I.; Apostolovic, D.; Milcic, M.; Stanic-Vucinic, D.; Cirkovic Velickovic, T. Complexes of green tea polyphenol, epigalocatechin-3-gallate, and 2S albumins of peanut. Food Chem. 2015, 185, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Structure of PR 10 Allergen Ara h 8.01 with Quercetin. RCSB PDB. 2022. Available online: https://www.rcsb.org/structure/6B1D (accessed on 19 January 2022).
- Hurlburt, B.K.; Offermann, L.R.; McBride, J.K.; Majorek, K.A.; Maleki, S.J.; Chruszcz, M. Structure and function of the peanut panallergen Ara h 8. J. Biol. Chem. 2013, 288, 36890–36901. [Google Scholar] [CrossRef] [PubMed]
- van Boxtel, E.L.; van den Broek, L.A.; Koppelman, S.J.; Vincken, J.P.; Gruppen, H. Peanut allergen Ara h 1 interacts with proanthocyanidins into higher molecular weight complexes. J. Agric. Food Chem. 2007, 55, 8772–8778. [Google Scholar] [CrossRef] [PubMed]
- Ohradanova-Repic, A.; Prazenicova, R.; Gebetsberger, L.; Moskalets, T.; Skrabana, R.; Cehlar, O.; Tajti, G.; Stockinger, H.; Leksa, V. Time to Kill and Time to Heal: The Multifaceted Role of Lactoferrin and Lactoferricin in Host Defense. Pharmaceutics 2023, 15, 1056. [Google Scholar] [CrossRef]
- Li, G.; Wei, Y.; Ma, L.; Mao, Y.; Xun, R.; Deng, Y. A novel highly sensitive soy aptasensor for antigen beta-conglycinin determination. Anal. Methods 2021, 13, 3059–3067. [Google Scholar] [CrossRef]
- Chinnappan, R.; Rahamn, A.A.; AlZabn, R.; Kamath, S.; Lopata, A.L.; Abu-Salah, K.M.; Zourob, M. Aptameric biosensor for the sensitive detection of major shrimp allergen, tropomyosin. Food Chem. 2020, 314, 126133. [Google Scholar] [CrossRef]
- Tipu, H.N.; Ahmed, D.; Gardezi, S.A.H. In silico identification of epitopes from house cat and dog proteins as peptide immunotherapy candidates based on human leukocyte antigen binding affinity. Iran J. Vet. Res. 2017, 18, 56–59. [Google Scholar]
- Wang, X.; Qiao, O.; Han, L.; Li, N.; Gong, Y. A Novel Rabbit Anti-Myoglobin Monoclonal Antibody’s Potential Application in Rhabdomyolysis Associated Acute Kidney Injury. Int. J. Mol. Sci. 2023, 24, 7822. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Fayomi, A.P.; Bai, S.; Gupta, N.; Cascio, S.; Yang, D.; Buckanovich, R.J. Generation and characterization of humanized affinity-matured EGFL6 antibodies for ovarian cancer therapy. Gynecol. Oncol. 2023, 171, 49–58. [Google Scholar] [CrossRef]
- Joyce, A.; Shea, C.; You, Z.; Gorovits, B.; Lepsy, C. Determination of Anti-drug Antibody Affinity in Clinical Study Samples Provides a Tool for Evaluation of Immune Response Maturation. AAPS J. 2022, 24, 114. [Google Scholar] [CrossRef] [PubMed]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Python Bindings. J. Chem. Inf. Model 2021, 61, 3891–3898. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Shahinozzaman, M.; Ishii, T.; Ahmed, S.; Halim, M.A.; Tawata, S. A computational approach to explore and identify potential herbal inhibitors for the p21-activated kinase 1 (PAK1). J. Biomol. Struct. Dyn. 2020, 38, 3514–3526. [Google Scholar] [CrossRef]
- Wang, W.; Liu, R.; Zhu, Y.; Wang, L.; Tang, Y.; Dou, B.; Tian, S.; Wang, F. YuNu-Jian attenuates diabetes-induced cardiomyopathy: Integrating network pharmacology and experimental validation. Front. Endocrinol. 2023, 14, 1195149. [Google Scholar] [CrossRef]
- Kizhakedathil, M.P.J.; Madasu, P.K.; Chandran, T.; Vijaykumar, S.D. In-silico structural studies on anti-inflammatory activity of phytocompounds from the genus Andrographis. J. Biomol. Struct. Dyn 2023, 1–13. [Google Scholar] [CrossRef]
- Almansour, N.M.; Allemailem, K.S.; Abd El Aty, A.A.; Ismail, E.I.F.; Ibrahim, M.A.A. In Silico Mining of Natural Products Atlas (NPAtlas) Database for Identifying Effective Bcl-2 Inhibitors: Molecular Docking, Molecular Dynamics, and Pharmacokinetics Characteristics. Molecules 2023, 28, 783. [Google Scholar] [CrossRef]
- Xia, J.; Hu, H.; Xue, W.; Wang, X.S.; Wu, S. The discovery of novel HDAC3 inhibitors via virtual screening and in vitro bioassay. J. Enzyme Inhib. Med. Chem. 2018, 33, 525–535. [Google Scholar] [CrossRef]
- Zeng, L.; Shin, W.H.; Zhu, X.; Park, S.H.; Park, C.; Tao, W.A.; Kihara, D. Discovery of Nicotinamide Adenine Dinucleotide Binding Proteins in the Escherichia coli Proteome Using a Combined Energetic- and Structural-Bioinformatics-Based Approach. J. Proteome Res. 2017, 16, 470–480. [Google Scholar] [CrossRef]
- Tanchuk, V.Y.; Tanin, V.O.; Vovk, A.I.; Poda, G. A New, Improved Hybrid Scoring Function for Molecular Docking and Scoring Based on AutoDock and AutoDock Vina. Chem. Biol. Drug Des. 2016, 87, 618–625. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, Y.; Liang, Y.; Dong, Y.; Yang, X.; Qiu, D. Verticillium dahliae PevD1, an Alt a 1-like protein, targets cotton PR5-like protein and promotes fungal infection. J. Exp. Bot. 2019, 70, 613–626. [Google Scholar] [CrossRef]
- Saenz-de-Santamaria, M.; Guisantes, J.A.; Martinez, J. Enzymatic activities of Alternaria alternata allergenic extracts and its major allergen (Alt a 1). Mycoses 2006, 49, 288–292. [Google Scholar] [CrossRef]
- Garrido-Arandia, M.; Tome-Amat, J.; Pazos-Castro, D.; Esteban, V.; Escribese, M.M.; Hernandez-Ramirez, G.; Yuste-Montalvo, A.; Barber, D.; Pacios, L.F.; Diaz-Perales, A. Interaction of Alt a 1 with SLC22A17 in the airway mucosa. Allergy 2019, 74, 2167–2180. [Google Scholar] [CrossRef]
- Hayes, T.; Rumore, A.; Howard, B.; He, X.; Luo, M.; Wuenschmann, S.; Chapman, M.; Kale, S.; Li, L.; Kita, H.; et al. Innate Immunity Induced by the Major Allergen Alt a 1 From the Fungus Alternaria Is Dependent Upon Toll-like Receptors 2/4 in Human Lung Epithelial Cells. Front. Immunol. 2018, 9, 1507. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Dai, Y.; Xia, Y. An overview of aryl hydrocarbon receptor ligands in the Last two decades (2002–2022): A medicinal chemistry perspective. Eur. J. Med. Chem. 2022, 244, 114845. [Google Scholar] [CrossRef]
- Koch, S.; Stroisch, T.J.; Vorac, J.; Herrmann, N.; Leib, N.; Schnautz, S.; Kirins, H.; Forster, I.; Weighardt, H.; Bieber, T. AhR mediates an anti-inflammatory feedback mechanism in human Langerhans cells involving FcepsilonRI and IDO. Allergy 2017, 72, 1686–1693. [Google Scholar] [CrossRef]
- Aguilera-Montilla, N.; Chamorro, S.; Nieto, C.; Sanchez-Cabo, F.; Dopazo, A.; Fernandez-Salguero, P.M.; Rodriguez-Fernandez, J.L.; Pello, O.M.; Andres, V.; Cuenda, A.; et al. Aryl hydrocarbon receptor contributes to the MEK/ERK-dependent maintenance of the immature state of human dendritic cells. Blood 2013, 121, e108–e117. [Google Scholar] [CrossRef] [PubMed]
- Maazi, H.; Shirinbak, S.; Bloksma, N.; Nawijn, M.C.; van Oosterhout, A.J. Iron administration reduces airway hyperreactivity and eosinophilia in a mouse model of allergic asthma. Clin. Exp. Immunol. 2011, 166, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Leung, S.; Holbrook, A.; King, B.; Lu, H.T.; Evans, V.; Miyamoto, N.; Mallari, C.; Harvey, S.; Davey, D.; Ho, E.; et al. Differential inhibition of inducible T cell cytokine secretion by potent iron chelators. J. Biomol. Screen. 2005, 10, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Jason, J.; Archibald, L.K.; Nwanyanwu, O.C.; Bell, M.; Jensen, R.J.; Gunter, E.; Buchanan, I.; Larned, J.; Kazembe, P.N.; Dobbie, H.; et al. The effects of iron deficiency on lymphocyte cytokine production and activation: Preservation of hepatic iron but not at all cost. Clin. Exp. Immunol. 2001, 126, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Thorson, J.A.; Smith, K.M.; Gomez, F.; Naumann, P.W.; Kemp, J.D. Role of iron in T cell activation: TH1 clones differ from TH2 clones in their sensitivity to inhibition of DNA synthesis caused by IgG Mabs against the transferrin receptor and the iron chelator deferoxamine. Cell Immunol. 1991, 134, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Drury, K.E.; Schaeffer, M.; Silverberg, J.I. Association between Atopic Disease and Anemia in US Children. JAMA Pediatr. 2016, 170, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Rhew, K.; Brown, J.D.; Oh, J.M. Atopic Disease and Anemia in Korean Patients: Cross-Sectional Study with Propensity Score Analysis. Int. J. Environ. Res. Public Health 2020, 17, 1978. [Google Scholar] [CrossRef]
- Rhew, K.; Oh, J.M. Association between atopic disease and anemia in pediatrics: A cross-sectional study. BMC Pediatr. 2019, 19, 455. [Google Scholar] [CrossRef]
- Shaheen, S.O.; Gissler, M.; Devereux, G.; Erkkola, M.; Kinnunen, T.I.; McArdle, H.; Sheikh, A.; Hemminki, E.; Nwaru, B.I. Maternal iron supplementation in pregnancy and asthma in the offspring: Follow-up of a randomised trial in Finland. Eur. Respir. J. 2020, 55, 1902335. [Google Scholar] [CrossRef]
- Shaheen, S.O.; Macdonald-Wallis, C.; Lawlor, D.A.; Henderson, A.J. Haemoglobin concentrations in pregnancy and respiratory and allergic outcomes in childhood: Birth cohort study. Clin. Exp. Allergy 2017, 47, 1615–1624. [Google Scholar] [CrossRef]
- Shaheen, S.O.; Newson, R.B.; Henderson, A.J.; Emmett, P.M.; Sherriff, A.; Cooke, M.; Team, A.S. Umbilical cord trace elements and minerals and risk of early childhood wheezing and eczema. Eur. Respir. J. 2004, 24, 292–297. [Google Scholar] [CrossRef]
- Nyakeriga, A.M.; Williams, T.N.; Marsh, K.; Wambua, S.; Perlmann, H.; Perlmann, P.; Grandien, A.; Troye-Blomberg, M. Cytokine mRNA expression and iron status in children living in a malaria endemic area. Scand J. Immunol. 2005, 61, 370–375. [Google Scholar] [CrossRef]
- Ghatak, S.K.; Majumdar, D.; Singha, A.; Sen, S.; Das, D.; Chakrabarti, A.; Mukhopadhyay, C.; Sen, K. Peanut protein sensitivity towards trace iron: A novel mode to ebb allergic response. Food Chem. 2015, 176, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Tong, P.; Gao, L.; Gao, J.; Li, X.; Wu, Z.; Yang, A.; Chen, H. Iron-induced chelation alleviates the potential allergenicity of ovotransferrin in a BALB/c mouse model. Nutr. Res. 2017, 47, 81–89. [Google Scholar] [CrossRef]
- Word, J.M.; Lovell, S.C.; Richardson, J.S.; Richardson, D.C. Asparagine and glutamine: Using hydrogen atom contacts in the choice of side-chain amide orientation. J. Mol. Biol. 1999, 285, 1735–1747. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Borrego, J.; Naseem, M.U.; Sehgal, A.N.A.; Panda, L.R.; Shakeel, K.; Gaspar, A.; Nagy, C.; Varga, Z.; Panyi, G. Recombinant Expression in Pichia pastoris System of Three Potent Kv1.3 Channel Blockers: Vm24, Anuroctoxin, and Ts6. J. Fungi 2022, 8, 1215. [Google Scholar] [CrossRef]
- Hufnagl, K.; Afify, S.M.; Braun, N.; Wagner, S.; Wallner, M.; Hauser, M.; Wiederstein, M.; Gadermaier, G.; Wildner, S.; Redegeld, F.A.; et al. Retinoic acid-loading of the major birch pollen allergen Bet v 1 may improve specific allergen immunotherapy: In silico, in vitro and in vivo data in BALB/c mice. Allergy 2020, 75, 2073–2077. [Google Scholar] [CrossRef]
- Roth-Walter, F.; Starkl, P.; Zuberbier, T.; Hummel, K.; Nobauer, K.; Razzazi-Fazeli, E.; Brunner, R.; Pali-Scholl, I.; Kinkel, J.; Felix, F.; et al. Glutathione exposes sequential IgE-epitopes in ovomucoid relevant in persistent egg allergy. Mol. Nutr. Food Res. 2013, 57, 536–544. [Google Scholar] [CrossRef]
- Oida, K.; Einhorn, L.; Herrmann, I.; Panakova, L.; Resch, Y.; Vrtala, S.; Hofstetter, G.; Tanaka, A.; Matsuda, H.; Jensen-Jarolim, E. Innate function of house dust mite allergens: Robust enzymatic degradation of extracellular matrix at elevated pH. World Allergy Organ. J. 2017, 10, 23. [Google Scholar] [CrossRef][Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fakhimahmadi, A.; Hasanaj, I.; Hofstetter, G.; Pogner, C.; Gorfer, M.; Wiederstein, M.; Szepannek, N.; Bianchini, R.; Dvorak, Z.; Jensen, S.A.; et al. Nutritional Provision of Iron Complexes by the Major Allergen Alt a 1 to Human Immune Cells Decreases Its Presentation. Int. J. Mol. Sci. 2023, 24, 11934. https://doi.org/10.3390/ijms241511934
Fakhimahmadi A, Hasanaj I, Hofstetter G, Pogner C, Gorfer M, Wiederstein M, Szepannek N, Bianchini R, Dvorak Z, Jensen SA, et al. Nutritional Provision of Iron Complexes by the Major Allergen Alt a 1 to Human Immune Cells Decreases Its Presentation. International Journal of Molecular Sciences. 2023; 24(15):11934. https://doi.org/10.3390/ijms241511934
Chicago/Turabian StyleFakhimahmadi, Aila, Ilir Hasanaj, Gerlinde Hofstetter, Clara Pogner, Markus Gorfer, Markus Wiederstein, Nathalie Szepannek, Rodolfo Bianchini, Zdenek Dvorak, Sebastian A. Jensen, and et al. 2023. "Nutritional Provision of Iron Complexes by the Major Allergen Alt a 1 to Human Immune Cells Decreases Its Presentation" International Journal of Molecular Sciences 24, no. 15: 11934. https://doi.org/10.3390/ijms241511934
APA StyleFakhimahmadi, A., Hasanaj, I., Hofstetter, G., Pogner, C., Gorfer, M., Wiederstein, M., Szepannek, N., Bianchini, R., Dvorak, Z., Jensen, S. A., Berger, M., Jensen-Jarolim, E., Hufnagl, K., & Roth-Walter, F. (2023). Nutritional Provision of Iron Complexes by the Major Allergen Alt a 1 to Human Immune Cells Decreases Its Presentation. International Journal of Molecular Sciences, 24(15), 11934. https://doi.org/10.3390/ijms241511934