

Small Molecules with Thiourea Skeleton Induce Ethylene Response in Arabidopsis

 and
and
Abstract
:1. Introduction
2. Results
2.1. Screening of Chemicals That Stimulate the Triple Response in Arabidopsis Thaliana from a Chemical Library
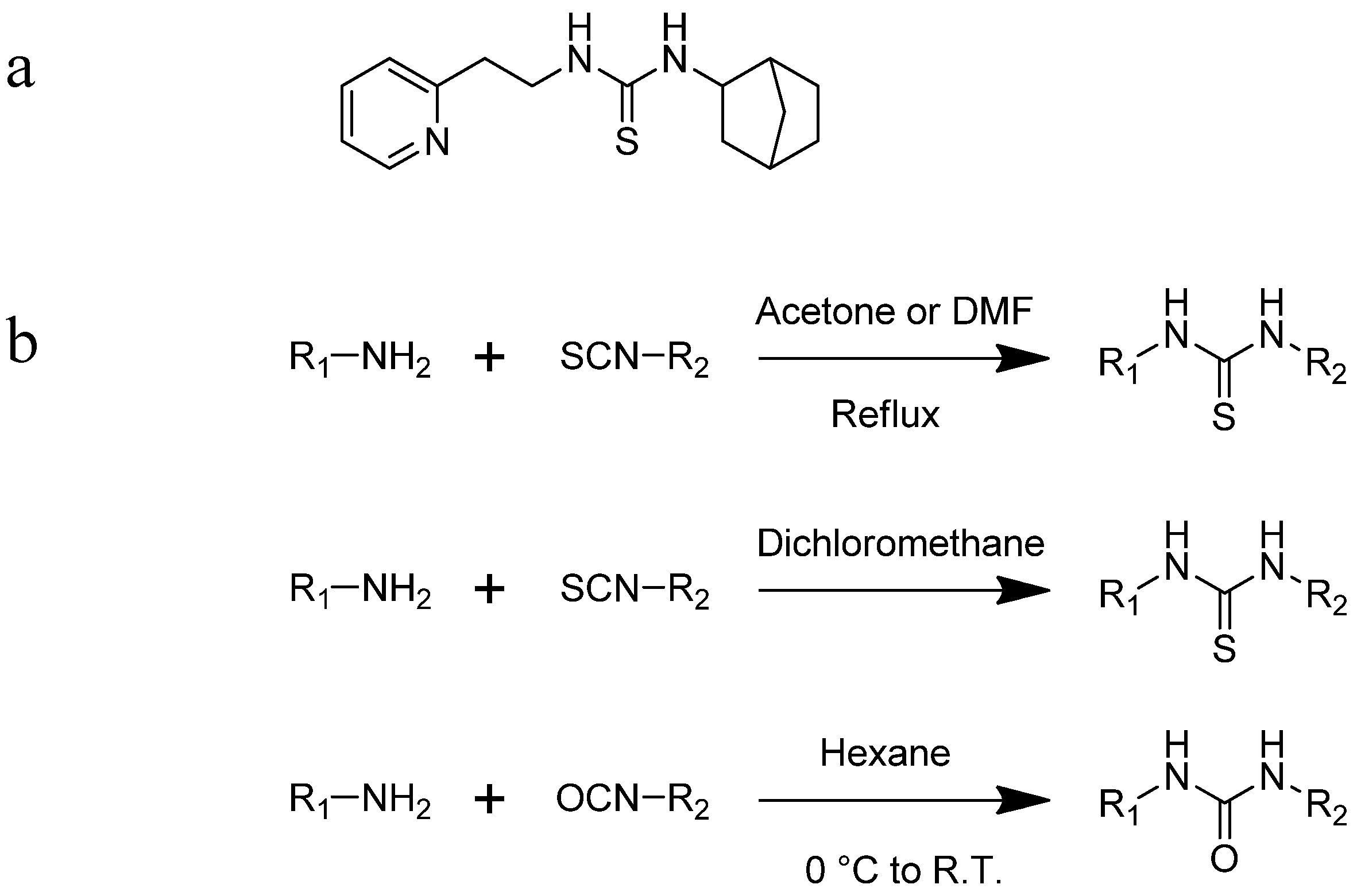
2.2. Synthesis of Derivatives of ZKT1
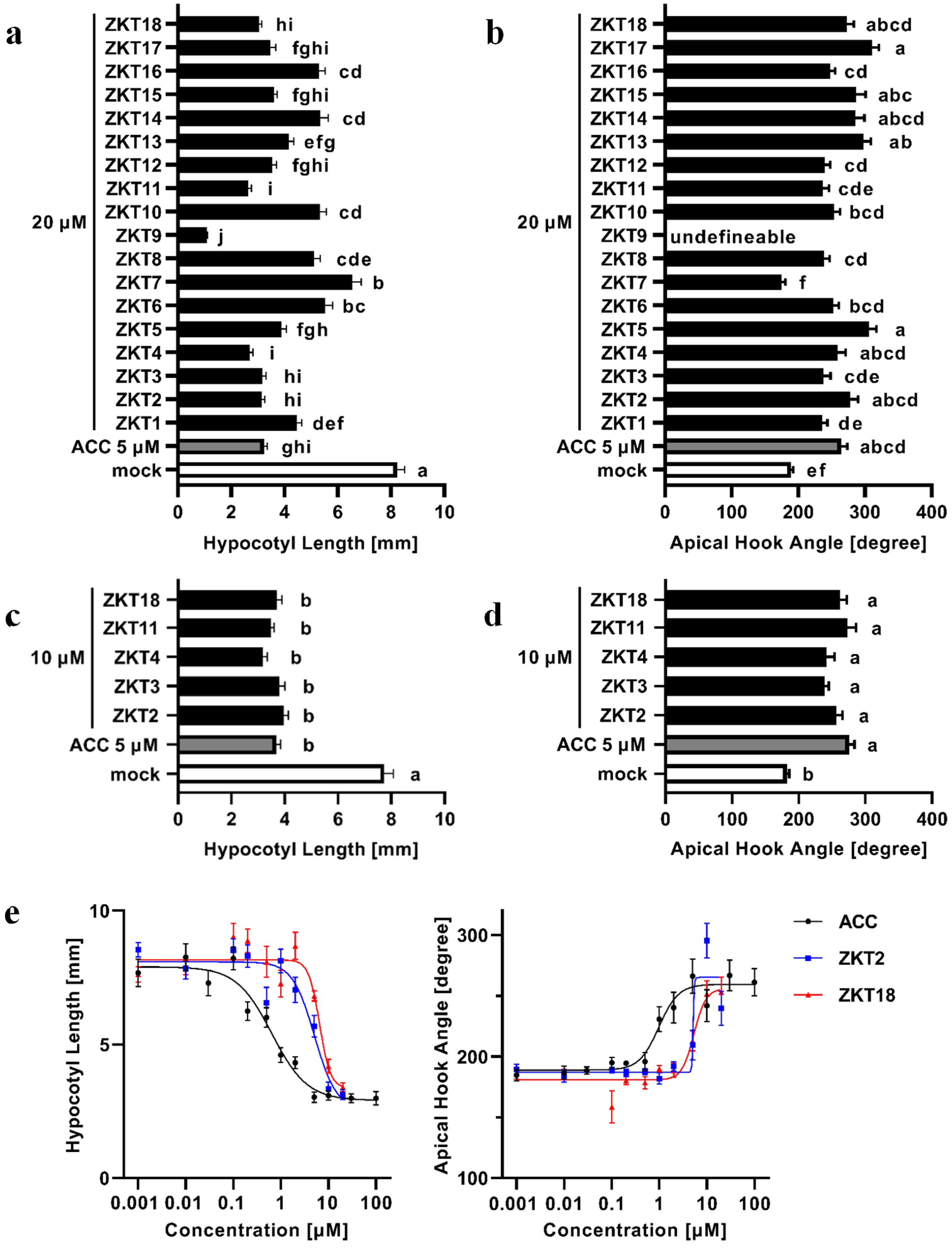
2.3. Evaluation of Ethylene-Like Activity of ZKT1 Derivatives
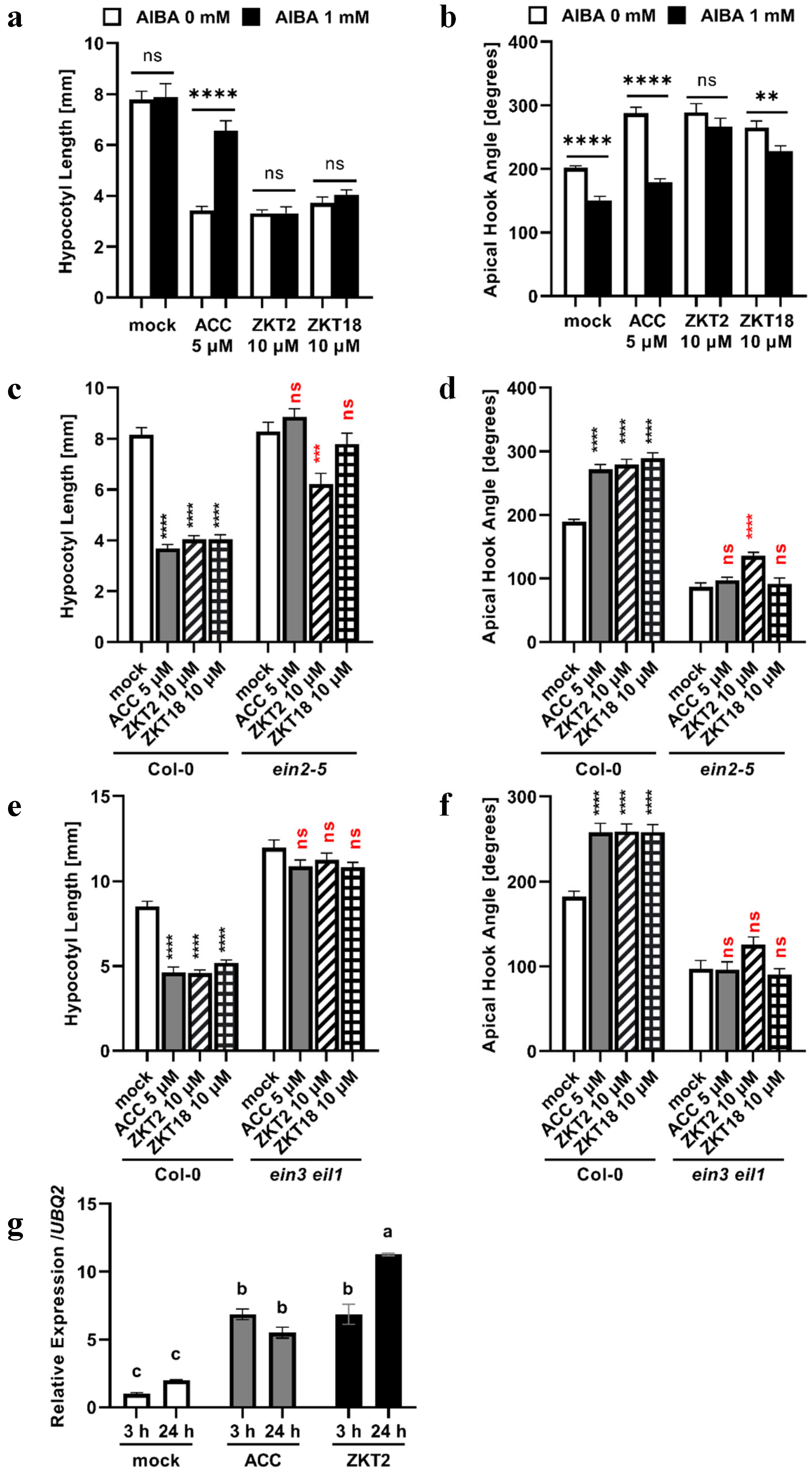
2.4. Analysis of the Mechanism of Action of ZKT Compounds
3. Discussion
3.1. Structure-Activity Relationship of ZKT1 Derivatives
3.2. Possible Mechanisms for ZKT1 Derivatives to Express Ethylene-Like Activity
3.3. Comparison of ZKT1 Derivatives with Existing Ethylene Mimics
4. Materials and Methods
4.1. General
4.2. Chemical Synthesis
- 1-(4-tert-butylphenyl)-3-(2-(pyridin-2-yl)ethyl)thiourea (ZKT2)
- 1-(4-bromophenyl)-3-(2-(pyridin-2-yl)ethyl)thiourea (ZKT3)
- 1-(2-(pyridin-2-yl)ethyl)-3-(4-(trifluoromethyl)phenyl)thiourea (ZKT4)
- 1-phenyl-3-(pyridin-2-yl)thiourea (ZKT5)
- 1-phenyl-3-(pyridin-2-ylmethyl)thiourea (ZKT6)
- 1-phenyl-3-(2-(pyridin-2-yl)ethyl)urea (ZKT7)
- 1-(2-chlorophenyl)-3-(pyridin-2-ylmethyl)thiourea (ZKT8)
- 1-(4-chlorophenyl)-3-(pyridin-2-ylmethyl)thiourea (ZKT9)
- 1-(4-fluorophenyl)-3-(pyridin-2-ylmethyl)thiourea (ZKT10)
- 1-(3,4-dichlorophenyl)-3-(pyridin-2-ylmethyl)thiourea (ZKT11)
- 1-(2,4-dichlorophenyl)-3-(pyridin-2-ylmethyl)thiourea (ZKT12)
- 1-(2,3-dichlorophenyl)-3-(pyridin-2-ylmethyl)thiourea (ZKT13)
- 1-(4-ethoxyphenyl)-3-(pyridin-2-ylmethyl)thiourea (ZKT14)
- 1-(4-tert-butylphenyl)-3-(pyridin-2-ylmethyl)thiourea (ZKT15)
- 1,3-diphenylthiourea (ZKT16)
- 1-benzyl-3-phenylthiourea (ZKT17)
- 1-phenetyl-3-phenylthiourea (ZKT18)
4.3. Plant Materials and Growth Conditions
4.4. Isolation of Total RNA
4.5. qRT-PCR Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bleecker, A.B.; Kende, H. ETHYLENE: A gaseous signal molecule in plants. Annu. Rev. Cell Dev. Biol. 2000, 16, 1–18. [Google Scholar] [PubMed] [Green Version]
- Larsen, P.B. Mechanisms of ethylene biosynthesis and response in plants. Essays Biochem. 2015, 58, 61–70. [Google Scholar]
- Lieberman, M.; Kunishi, A.T.; Mapson, L.W.; Wardale, D.A. Ethylene production from methionine. Biochem. J. 1965, 97, 449–459. [Google Scholar] [PubMed]
- Adams, D.O.; Yang, S.F. Methionine metabolism in apple tissue: Implication of S-adenosylmethionine as an intermediate in the conversion of methionine to ethylene. Plant Physiol. 1977, 60, 892–896. [Google Scholar] [PubMed] [Green Version]
- Adams, D.O.; Yang, S.F. Ethylene biosynthesis: Identification of 1-aminocyclopropane-1-carboxylic acid as an intermediate in the conversion of methionine to ethylene. Proc. Natl. Acad. Sci. USA 1979, 76, 170–174. [Google Scholar] [CrossRef]
- Lürssen, K.; Naumann, K.; Schröder, R. 1-Aminocyclopropane-1-carboxylic acid—A new intermediate of ethylene biosynthesis. Naturwissenschaften 1979, 66, 264–265. [Google Scholar] [CrossRef]
- Lürssen, K.; Naumann, K.; Schröder, R. 1-Aminocyclopropane-1-carboxylic acid—An intermediate of the ethylene biosynthesis in higher plants. Z. Für Pflanzenphysiol. 1979, 92, 285–294. [Google Scholar] [CrossRef]
- Chang, C.; Kwok, S.F.; Bleecker, A.B.; Meyerowitz, E.M. Arabidopsis ethylene-response gene ETR1: Similarity of product to two-component regulators. Science 1993, 262, 539–544. [Google Scholar]
- Chen, Y.F.; Randlett, M.D.; Findell, J.L.; Schaller, G.E. Localization of ethylene receptor ETR1 to the endoplasmic reticulum of Arabidopsis. J. Biol. Chem. 2002, 277, 19861–19866. [Google Scholar] [CrossRef] [Green Version]
- Grefen, C.; Stadele, K.; Rzicka, K.; Obrdlik, P.; Hater, K.; Horak, J. Subcellular localization and in vivo interactions of the Arabidopsis thaliana ethylene receptor family members. Mol. Plant 2008, 1, 308–320. [Google Scholar] [CrossRef]
- Kieber, J.J.; Rothenberg, M.; Roman, G.; Feldmann, K.; Ecker, J.R. CTR1, a negative regulator of the ethylene response pathway in Arabidopsis, encodes a member of the Raf family of protein kinase. Cell 1993, 72, 427–441. [Google Scholar] [PubMed]
- Clark, K.L.; Larsen, P.B.; Wang, X.; Chang, C. Association of the Arabidopsis CTR1 Raf-like kinase with the ETR1 and ERS ethylene receptors. Proc. Natl. Acad. Sci. USA 1998, 95, 5401–5406. [Google Scholar] [PubMed]
- Qu, X.; Schaller, G.E. Requirement of the histidine kinase domain for signal transduction by the ethylene receptor ETR1. Plant Physiol. 2004, 136, 2961–2970. [Google Scholar] [PubMed] [Green Version]
- Zhong, S.; Lin, Z.; Grierson, D. Tomato ethylene receptor-CTR intractions; Visualization of NEVER-RIPE interactions with multiple CTRs at the endoplasmic reticulum. J. Exp. Bot. 2008, 59, 965–972. [Google Scholar]
- Ju, C.; Yoon, G.M.; Shemansky, J.M.; Lin, D.Y.; Ying, Z.I.; Chang, J.; Garrett, W.M.; Kessenbrock, M.; Groth, G.; Tucker, M.L.; et al. CTR1 phosphorylates the central regulator EIN2 to control ethylene hormone signaling from the ER membrane to the nucleus in Arabidopsis. Proc. Natl. Acad. Sci. USA 2012, 109, 19486–19491. [Google Scholar]
- Guo, H.; Ecker, J.R. Plant responses to ethylene gas are mediated by SCF(EBF1/EBF2)-dependent proteolysis of EIN3 transcription factor. Cell 2003, 115, 667–677. [Google Scholar]
- Qiao, H.; Shen, Z.; Huang, S.S.; Shmitz, R.J.; Urich, M.A.; Briggs, S.P.; Ecker, J.R. Processing and subcellular trafficking of ER-tethered EIN2 control response to ethylene gas. Science 2012, 338, 390–393. [Google Scholar]
- Wen, X.; Zhang, C.; Ji, Y.; Zhao, Q.; He, W.; An, F.; Jiang, L.; Guo, H. Activation of ethylene signaling is mediated by nuclear translocation of the cleaved EIN2 carboxyl terminus. Cell Res. 2012, 22, 1613–1616. [Google Scholar]
- Solano, R.; Stepanova, A.; Chao, Q.; Ecker, J.R. Nuclear events in ethylene signaling: A transcriptional cascade mediated by ETHYLENE-INSENSITIVE3 and ETHYLENE-RESPONSE-FACTOR1. Genes Dev. 1998, 12, 3703–3714. [Google Scholar] [PubMed] [Green Version]
- Binder, B.M. Ethylene signaling in plants. J. Biol. Chem. 2020, 295, 7710–7725. [Google Scholar]
- Yang, S.F. Ethylene evolution from 2-chloroethylphosphonic acid. Plant Physiol. 1969, 44, 1203–1204. [Google Scholar]
- Oh, K.; Hoshi, T.; Tomio, S.; Ueda, K.; Hara, K. A chemical genetics strategy that identifies small molecules which induce the triple response in Arabidopsis. Molecules 2017, 22, 2270. [Google Scholar] [PubMed] [Green Version]
- Pennisi, E. Armed and dangerous. Science 2010, 327, 804. [Google Scholar] [PubMed] [Green Version]
- Kountche, B.A.; Jamil, M.; Yonli, D.; Nikiema, M.P.; Blanco-Ania, D.; Asami, T.; Zwanenburg, B.; Al-Babili, S. Suicidal germination as a control strategy for Striga hermonthica (Benth.) in smallholder farms of sub-Saharan Africa. Plants People Planet 2019, 1, 107–118. [Google Scholar]
- Iverson, R.D.; Westbrooks, R.G.; Eplee, R.E.; Tasker, A.V. Overview and status of the witchweed (Striga asiatica) eradication program in the Carolinas. In Invasive Plant Management Issues and Challenges in the United States: 2011 Overview; Lesilie, A.R., Westbrooks, R.G., Eds.; ACS Publications: Washington, DC, USA, 2011; pp. 51–68. [Google Scholar]
- Guzmán, P.; Ecker, J.R. Exploiting the triple response of Arabidopsis to identify ethylene-related mutants. Plant Cell 1990, 2, 513–523. [Google Scholar]
- Vandenbussche, F.; Petrášek, J.; Žádníková, P.; Hoyerová, K.; Pešek, B.; Raz, V.; Swarup, R.; Bennett, M.; Zažímalová, E.; Benková, E.; et al. The auxin influx carriers AUX1 and LAX3 are involved in auxin-ethylene interactions during apical hook development in Arabidopsis thaliana seedlings. Development 2010, 137, 597–606. [Google Scholar] [CrossRef] [Green Version]
- Satoh, S.; Esashi, Y. α-Aminoisobutyric acid: A probable competitive inhibitor of conversion of 1-aminocylopropane-1-carboxylic acid to ethylene. Plant Cell Physiol. 1980, 21, 939–949. [Google Scholar]
- Scharein, B.; Vormizeele, J.V.; Harter, K.; Groth, G. Ethylene signaling: Identification of a putative ETR1-AHP1 phosphorelay complex by fluorescence spectroscopy. Anal. Biochem. 2008, 377, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Urao, T.; Miyata, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Possible His to Asp phosphorelay signaling in an Arabidopsis two-component system. FEBS Lett. 2000, 478, 227–232. [Google Scholar] [PubMed] [Green Version]
- Zdarska, M.; Cuyacot, A.R.; Tarr, P.T.; Yamoune, A.; Szmitkowska, A.; Hrdinová, B.; Gelová, Z.; Meyerowitz, E.M.; Hejátko, J. ETR1 integrates response to ethylene and cytokinins into a single multistep phosphorelay pathway to control root growth. Mol. Plant 2019, 12, 1338–1352. [Google Scholar]
- Rodriguez, F.I.; Esch, J.J.; Hall, A.E.; Binder, B.M.; Schaller, G.E.; Bleecker, A.B. A copper cofactor for the ethylene receptor ETR1 from Arabidopsis. Science 1999, 283, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Suenaga, Y.; Wu, L.P.; Kuroda-sowa, T.; Munakata, M.; Maekawa, M. Structure and 1H NMR study of copper (I) complex with ethylene and tetramethylethylenediamine. Polyhedron 1997, 16, 67–70. [Google Scholar]
- Hirsch, J.; George, S.D.; Solomon, E.I.; Hedman, B.; Hodgson, K.O.; Burstyn, J.N. Raman and extended X-ray absorption fine structure characterization of a sulfur-ligated Cu(I) ethylene-complex: Modeling the proposed ethylene binding site of Arabidopsis thaliana ETR1. Inorg. Chem. 2001, 40, 2439–2441. [Google Scholar] [CrossRef]
- Hoppen, C.; Müller, L.; Hänsch, S.; Uzun, B.; Milić, D.; Meyer, A.J.; Weidtkamp-Peters, S.; Groth, G. Soluble and membrane-bound protein carrier mediate direct copper transport to the ethylene receptor family. Sci. Rep. 2019, 9, 10715. [Google Scholar] [PubMed] [Green Version]
- Woeste, K.E.; Kieber, J.J. A strong loss-of-function mutation in RAN1 results in constitutive activation of the ethylene response pathway as well as a rosette-lethal phenotype. Plant Cell 2000, 12, 443–455. [Google Scholar] [CrossRef] [Green Version]
- Knobler, C.B.; Okaya, Y.; Pepinsky, R. The crystal structure of tris(thiourea)copper(I) chloride, Cu(SCN2H4)3Cl. Z. Für Krist. Cryst. Mater. 1959, 111, 385–397. [Google Scholar] [CrossRef]
- Hanic, F.; Ďurčanská, E. The crystal structure of tris(thiourea)copper(I) perchlorate Cu(SCN2H4)3ClO4. Inorg. Chim. Acta 1969, 3, 293–298. [Google Scholar] [CrossRef]
- Hollmann, K.; Oppermann, A.; Witte, M.; Li, S.; Amen, M.; Flörke, U.; Egold, H.; Henkel, G.; Herres-Pawlis, S. Copper(I) complexes with thiourea derivatives as ligands: Revealing secrets of their bonding scheme. Eur. J. Inorg. Chem. 2017, 2017, 1266–1279. [Google Scholar] [CrossRef]
- Arslan, H.; Kulcu, N.; Florke, U. Synthesis and characterization of copper(II), nickel(II) and cobalt(II) complexes with novel thiourea derivatives. Transit. Metal Chem. 2003, 28, 816–819. [Google Scholar] [CrossRef]
- Binder, B.M.; Rodríguez, F.I.; Bleecker, A.B. The copper transporter RAN1 is essential for biogenesis of ethylene receptors in Arabidopsis. J. Biol. Chem. 2010, 285, 37263–37270. [Google Scholar] [PubMed] [Green Version]
- Chen, S.H.; Lin, J.K.; Liu, S.H.; Liang, Y.C.; Lin-Shiau, S.Y. Apoptosis of cultured astrocytes induced by the copper and neocuproine complex through oxidative stress and JNK activation. Toxicol. Sci. 2008, 102, 138–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Lacey, R.F.; Ye, Y.; Lu, J.; Yeh, K.C.; Xiao, Y.; Li, L.; Wen, C.K.; Binder, B.M.; Zhao, Y. Triplin, a small molecule, reveals copper ion transport in ethylene signaling from ATX1 to RAN1. PLoS Genet. 2017, 13, e1006703. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
| Entry | Structure | Entry | Structure |
|---|---|---|---|
| ZKT2 |  | ZKT11 | 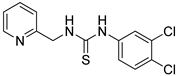 |
| ZKT3 | 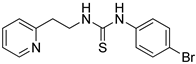 | ZKT12 | 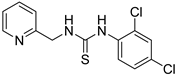 |
| ZKT4 | 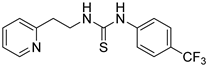 | ZKT13 | 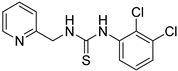 |
| ZKT5 |  | ZKT14 | 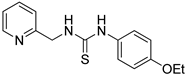 |
| ZKT6 | 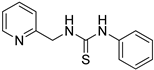 | ZKT15 | 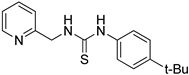 |
| ZKT7 | 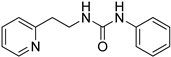 | ZKT16 |  |
| ZKT8 | 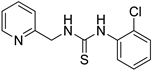 | ZKT17 | 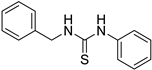 |
| ZKT9 | 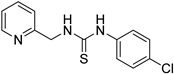 | ZKT18 | 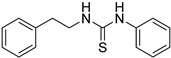 |
| ZKT10 | 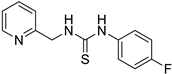 |
| Entry | EC50 Values Calculated from Hypocotyl Length | EC50 Values Calculated from Apical Hook Angle |
|---|---|---|
| ACC | 0.648 (μM) | 0.976 (μM) |
| ZKT2 | 5.07 (μM) | 5.14 (μM) |
| ZKT18 | 6.49 (μM) | 5.31 (μM) |
| Primer Name | Forward/Reverse | Primer Sequence |
|---|---|---|
| AtERF1F_Fwd | F | AGACGACGGCCATGGGGGAA |
| AtETR1R_Rev | R | TCCGCGCTTTCGAACGTCCC |
| AtUBQ2_Fwd | F | CCAAGATCCAGGACAAAGAAGGA |
| AtUBQ2_Rev | R | TGGAGACGAGAGCATAACACTTGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koyama, T.; Zaizen, H.; Takahashi, I.; Nakamura, H.; Nakajima, M.; Asami, T. Small Molecules with Thiourea Skeleton Induce Ethylene Response in Arabidopsis. Int. J. Mol. Sci. 2023, 24, 12420. https://doi.org/10.3390/ijms241512420
Koyama T, Zaizen H, Takahashi I, Nakamura H, Nakajima M, Asami T. Small Molecules with Thiourea Skeleton Induce Ethylene Response in Arabidopsis. International Journal of Molecular Sciences. 2023; 24(15):12420. https://doi.org/10.3390/ijms241512420
Chicago/Turabian StyleKoyama, Tomoyuki, Honami Zaizen, Ikuo Takahashi, Hidemitsu Nakamura, Masatoshi Nakajima, and Tadao Asami. 2023. "Small Molecules with Thiourea Skeleton Induce Ethylene Response in Arabidopsis" International Journal of Molecular Sciences 24, no. 15: 12420. https://doi.org/10.3390/ijms241512420
APA StyleKoyama, T., Zaizen, H., Takahashi, I., Nakamura, H., Nakajima, M., & Asami, T. (2023). Small Molecules with Thiourea Skeleton Induce Ethylene Response in Arabidopsis. International Journal of Molecular Sciences, 24(15), 12420. https://doi.org/10.3390/ijms241512420