
The Molecular Mechanisms Employed by the Parasite Myxobolus bejeranoi (Cnidaria: Myxozoa) from Invasion through Sporulation for Successful Proliferation in Its Fish Host

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
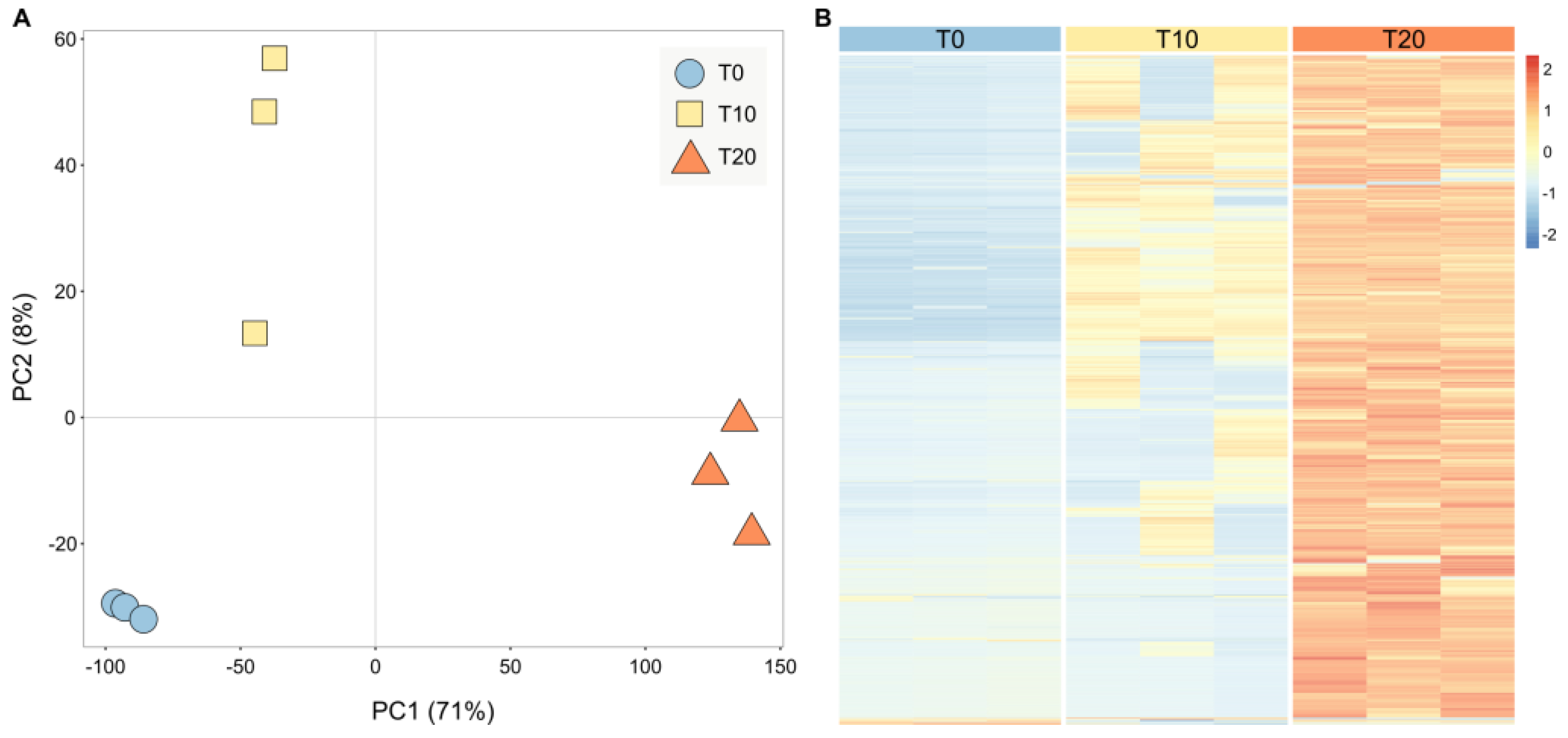
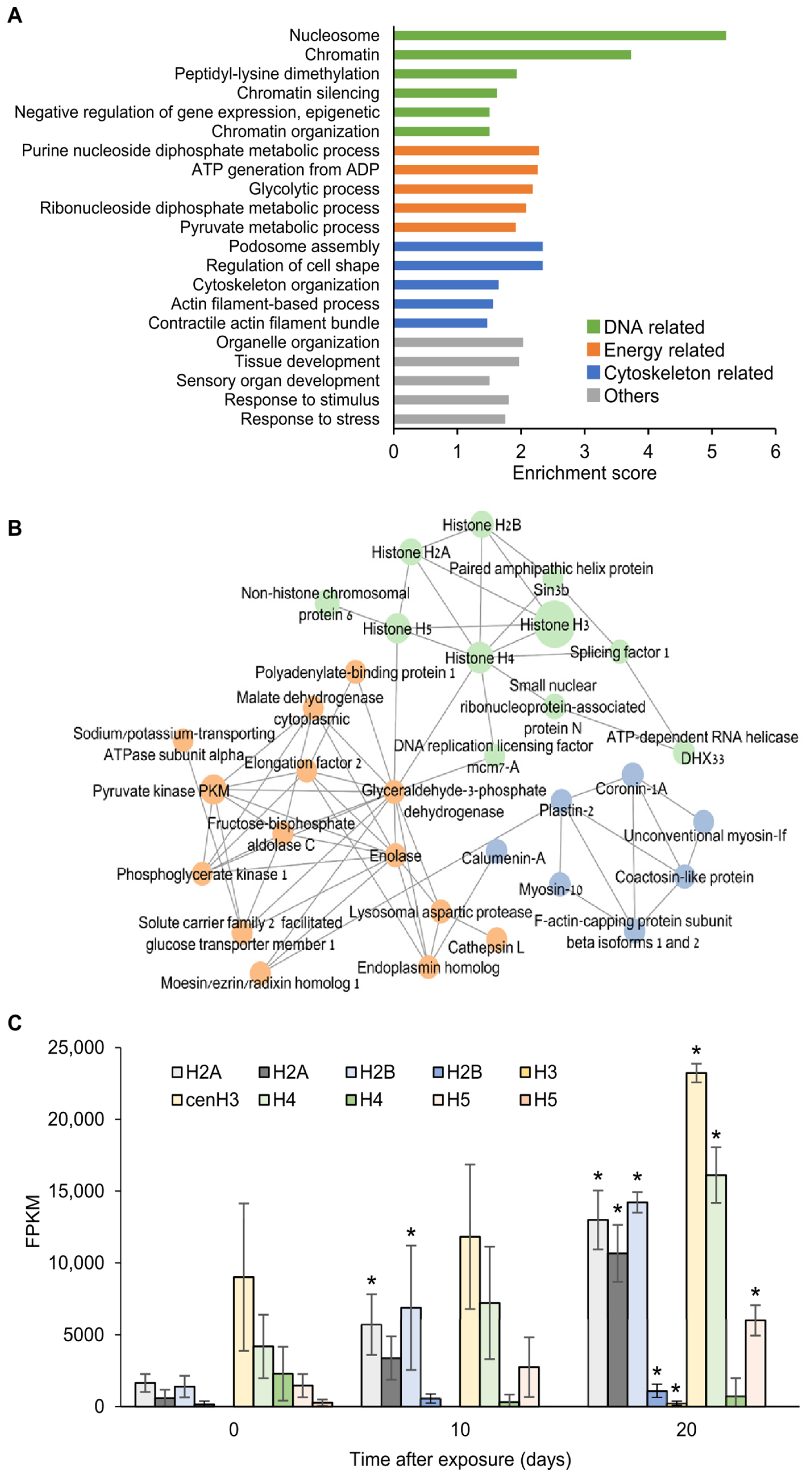
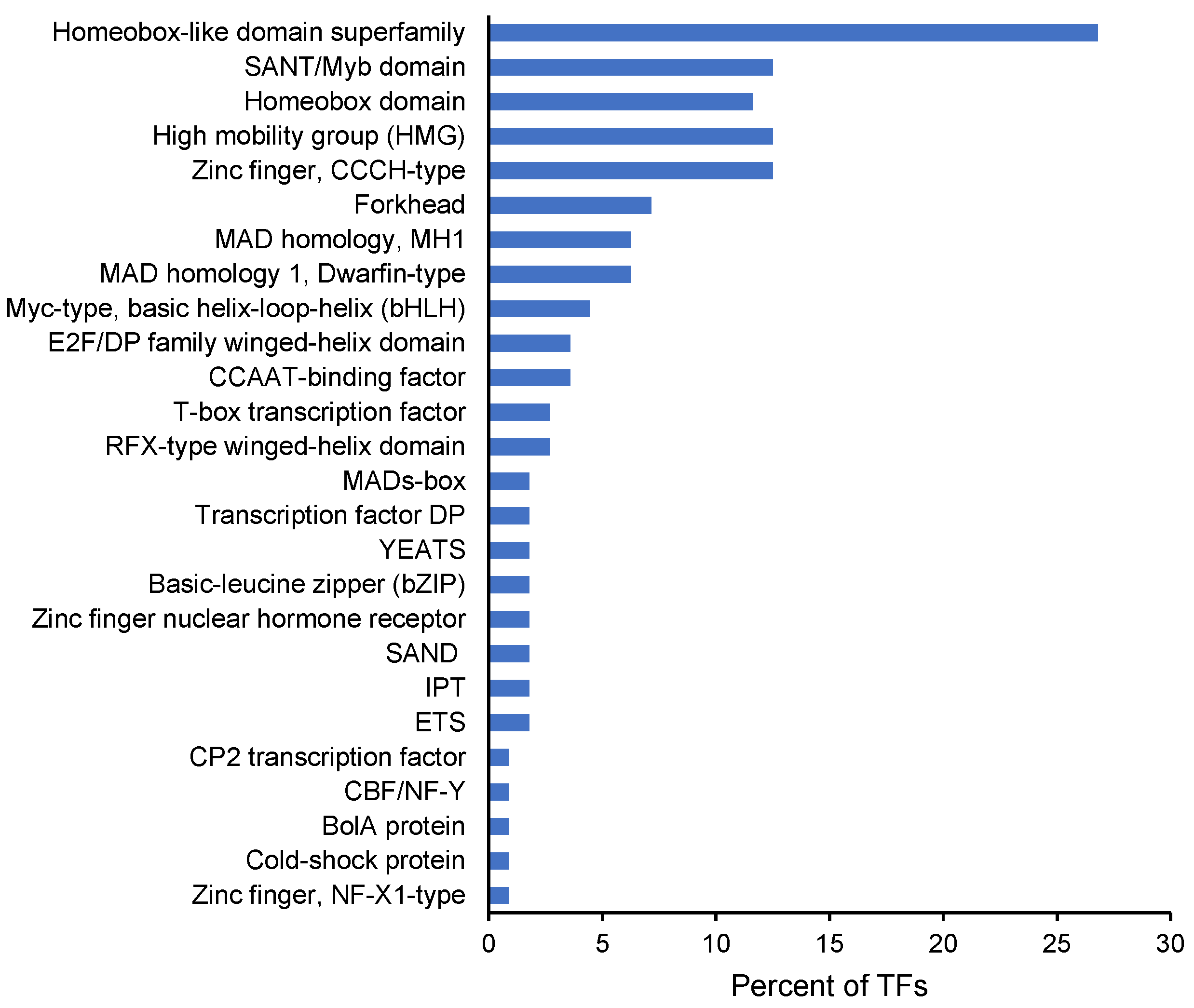
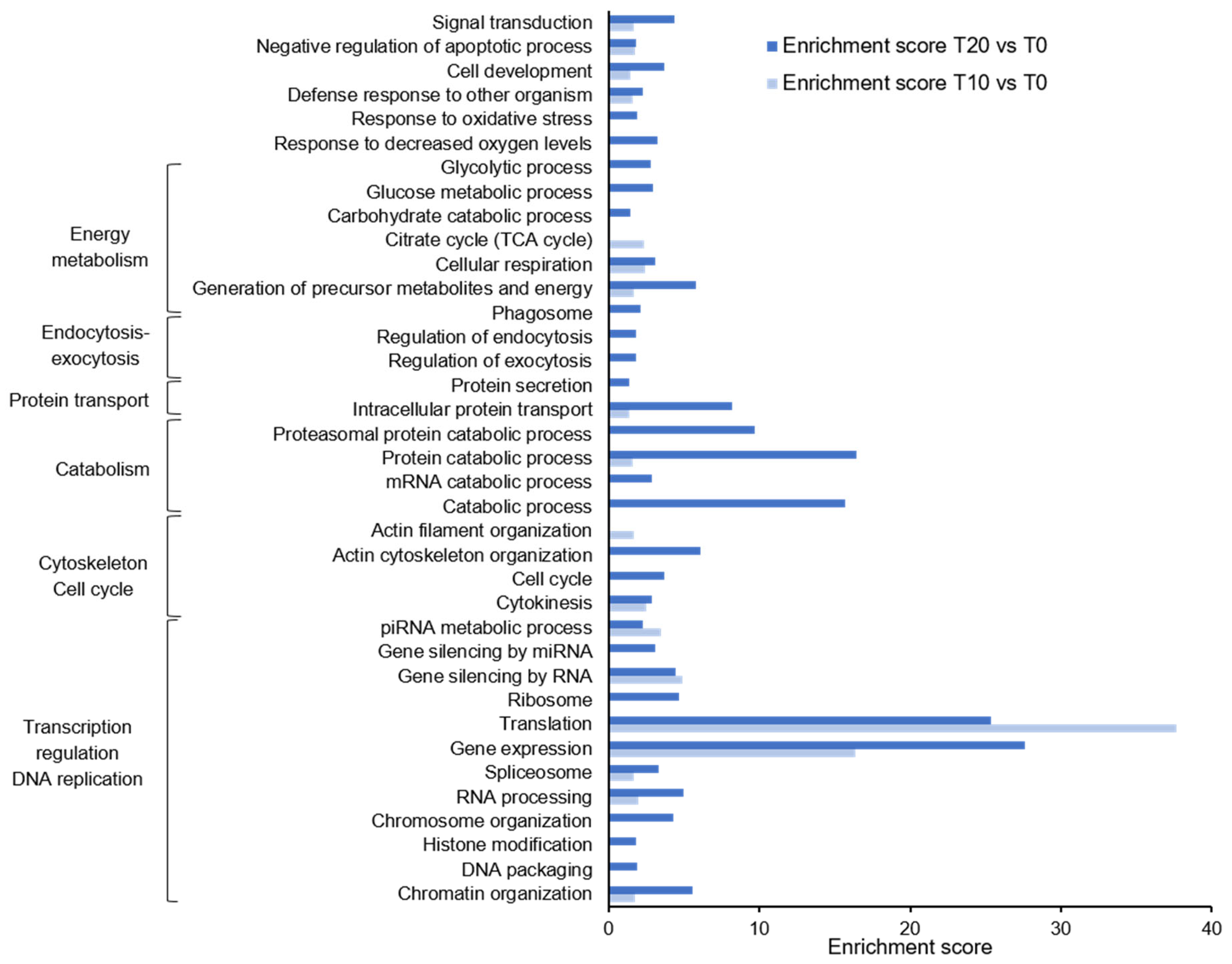
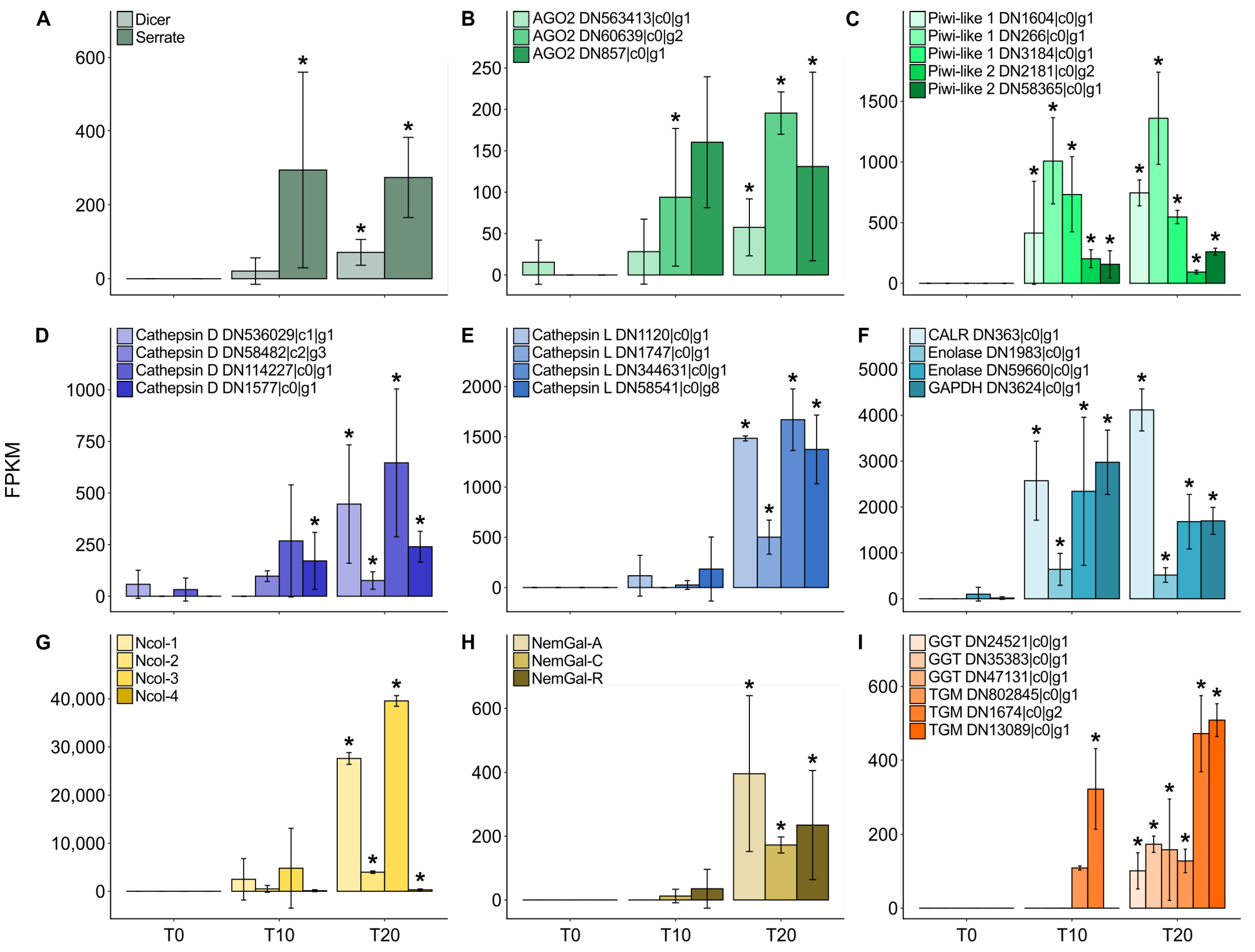
2. Results
3. Discussion
4. Materials and Methods
4.1. Fish Infection Experimental Design
4.2. RNA and DNA Extraction and Evaluation of Infection Severity
4.3. Sequencing and Transcriptome Assembly
4.4. Differential Gene Expression Analysis and Annotation
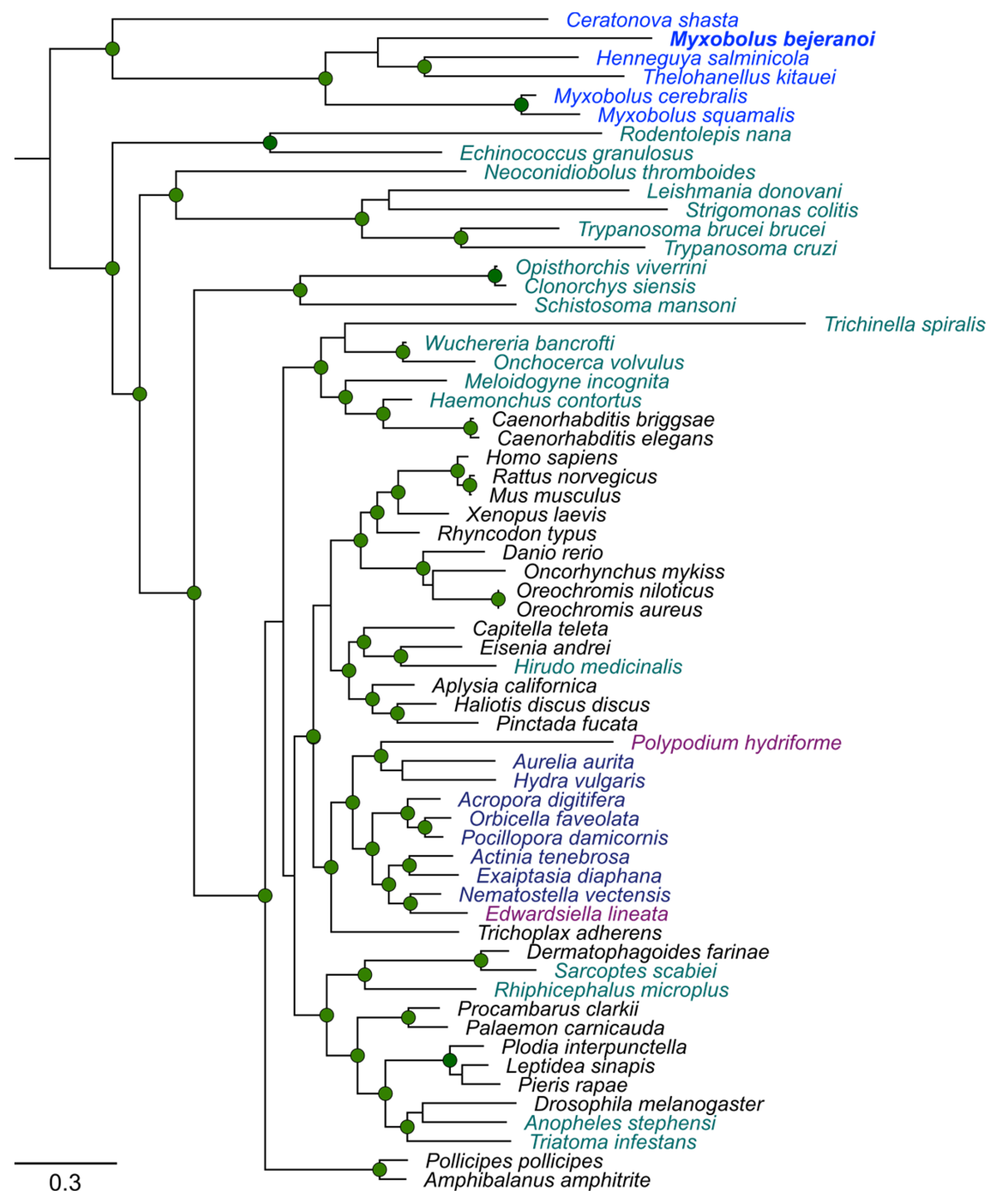
4.5. Phylogenetic Trees
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frank, S.A. Models of parasite virulence. Q. Rev. Biol. 1996, 71, 37–78. [Google Scholar] [CrossRef]
- Poulin, R. Are there general laws in parasite ecology? Parasitology 2007, 134, 763–776. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, S.D.; Bartholomew, J.L.; Lotan, T. Myxozoans: Ancient metazoan parasites find a home in phylum Cnidaria. Zoology 2018, 129, 66–68. [Google Scholar] [CrossRef]
- Chang, E.S.; Neuhof, M.; Rubinstein, N.D.; Diamant, A.; Philippe, H.; Huchon, D.; Cartwright, P. Genomic insights into the evolutionary origin of Myxozoa within Cnidaria. Proc. Natl. Acad. Sci. USA 2015, 112, 14912–14917. [Google Scholar] [CrossRef]
- Yang, Y.; Xiong, J.; Zhou, Z.; Huo, F.; Miao, W.; Ran, C.; Liu, Y.; Zhang, J.; Feng, J.; Wang, M.; et al. The genome of the myxosporean Thelohanellus kitauei shows adaptation to nutrient acquisition within its fish host. Genome Biol. Evol. 2014, 6, 3182–3198. [Google Scholar] [CrossRef] [Green Version]
- Eszterbauer, E.; Atkinson, S.; Diamant, A.; Morris, D.; El-Matbouli, M.; Hartikainen, H. Myxozoan Life Cycles: Practical approaches and insights. In Myxozoan Evolution, Ecology and Development; Okamura, B., Gruhl, A., Bartholomew, J.L., Eds.; Springer: Cham, Switzerland, 2015; pp. 175–198. [Google Scholar]
- Holzer, A.S.; Barto, P.; Born-torrijos, A.; Alena, L.; Hartigan, A.; Fiala, I. The joint evolution of the Myxozoa and their alternate hosts: A cnidarian recipe for success and vast biodiversity. Mol. Ecol. 2018, 27, 1651–1666. [Google Scholar] [CrossRef] [PubMed]
- Wolf, K.; Markiw, M.E. Biology contravenes taxonomy in the Myxozoa: New discoveries show alternation of invertebrate and vertebrate hosts. Science 1984, 225, 1449–1452. [Google Scholar] [CrossRef]
- Kent, M.; Margolis, L.; Corliss, J. The demise of a class of protists: Taxonomic and nomenclatural revisions proposed for the protist phylum Myxozoa Grasse, 1970. Can. J. Zool. 1994, 72, 932–937. [Google Scholar] [CrossRef]
- Ben-David, J.; Atkinson, S.D.; Pollak, Y.; Yossifon, G.; Shavit, U.; Bartholomew, J.L.; Lotan, T. Myxozoan polar tubules display structural and functional variation. Parasit. Vectors 2016, 9, 549. [Google Scholar] [CrossRef] [Green Version]
- Kallert, D.M.; Ponader, S.; Eszterbauer, E.; El-Matbouli, M.; Haas, W. Myxozoan transmission via actinospores: New insights into mechanisms and adaptations for host invasion. Parasitology 2007, 134, 1741–1750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallert, D.M.; El-Matbouli, M.; Haas, W. Polar filament discharge of Myxobolus cerebralis actinospores is triggered by combined non-specific mechanical and chemical cues. Parasitology 2005, 131, 609–616. [Google Scholar] [CrossRef] [Green Version]
- Americus, B.; Austin, B.M.; Lotan, T.; Bartholomew, J.L.; Atkinson, S.D. In vitro and in vivo assays reveal that cations affect nematocyst discharge in Myxobolus cerebralis (Cnidaria: Myxozoa). Parasitology 2020, 147, 1352–1358. [Google Scholar] [CrossRef]
- Americus, B.; Lotan, T.; Bartholomew, J.L.; Stephen, D. A comparison of the structure and function of nematocysts in free-living and parasitic cnidarians (Myxozoa). Int. J. Parasitol. 2020, 50, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Kallert, D.M.; Grabner, D.; Yokoyama, H.; El-Matbouli, M.; Eszterbauer, E. Transmission of Myxozoans to vertebrate hosts. In Myxozoan Evolution, Ecology and Development; Okamura, B., Gruhl, A., Bartholomew, J.L., Eds.; Springer: Cham, Switzerland, 2015; pp. 235–251. [Google Scholar]
- Lom, J.; Dyková, I. Myxozoan genera definition and notes on taxonomy, life-cycle terminology and pathogenic species. Folia Parasitol. 2006, 53, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, M.A.; Granath, W.O. Persistent infection of Myxobolus cerebralis, the causative agent of salmonid whirling disease, in Tubifex tubifex. J. Parasitol. 2001, 87, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Lövy, A.; Smirnov, M.; Brekhman, V.; Ofek, T.; Lotan, T. Morphological and molecular characterization of a novel myxosporean parasite Myxobolus bejeranoi n. sp. (Cnidaria: Myxosporea) from hybrid tilapia in Israel. Parasitol. Res. 2018, 117, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Maor-Landaw, K.; Smirnov, M.; Brekhman, V.; Ofek-lalzar, M.; Yahav, T.; Lotan, T. Infection by the Parasite Myxobolus bejeranoi (Cnidaria: Myxozoa) suppresses the immune system of hybrid tilapia. Microorganisms 2022, 10, 1893. [Google Scholar] [CrossRef] [PubMed]
- Maor-Landaw, K.; Avidor, I.; Salti, B.; Smirnov, M.; Brekhman, V.; Lotan, T. The myxozoan parasite Myxobolus bejeranoi (Cnidaria: Myxozoa) infection dynamics and host specificity in hybrid tilapia aquaculture. Parasitology 2023, 150, 524–530. [Google Scholar] [CrossRef]
- El-Sayed, A.-F.M. Tilapia Culture, 2nd ed.; El-Sayed, A.-F.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; ISBN 9780128165096. [Google Scholar]
- Hulata, G. Tilapias-Biology and Culture; Ma’arechet Publishing House: Dalia, Israel, 2014. [Google Scholar]
- Alama-Bermejo, G.; Holzer, A.S. Advances and discoveries in myxozoan genomics. Trends Parasitol. 2021, 37, 552–568. [Google Scholar] [CrossRef]
- Draizen, E.J.; Shaytan, A.K.; Marino-Ramirez, L.; Talbert, P.B.; Landsman, D.; Panchenko, A.R. HistoneDB 2.0: A histone database with variants—An integrated resource to explore histones and their variants. Database 2016, 2016, baw014. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, M. The Role of Synaptotagmin and slp in regulated exocytosis. In Molecular Mechanisms of Exocytosis; Regazzi, R., Ed.; Springer: New York, NY, USA, 2007; pp. 42–61. [Google Scholar]
- Mishima, T.; Fujiwara, T.; Sanada, M.; Kofuji, T.; Kanai-azuma, M. Syntaxin 1B, but not syntaxin 1A, is necessary for the regulation of synaptic vesicle exocytosis and of the readily releasable pool at central synapses. PLoS ONE 2014, 9, e90004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Hui, J.H.L. Small RNAs in Cnidaria: A review. Evol. Appl. 2022, 16, 354–364. [Google Scholar] [CrossRef]
- Ramírez-Toloza, G.; Sosoniuk-Roche, E.; Valck, C.; Aguilar-Guzmán, L.; Ferreira, V.P.; Ferreira, A. Trypanosoma cruzi calreticulin: Immune evasion, infectivity, and tumorigenesis. Trends Parasitol. 2020, 36, 368–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shpirer, E.; Chang, E.S.; Diamant, A.; Rubinstein, N.; Cartwright, P.; Huchon, D. Diversity and evolution of myxozoan minicollagens and nematogalectins. BMC Evol. Biol. 2014, 14, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szczepanek, S.; Cikala, M.; David, C.N. Poly-γ-glutamate synthesis during formation of nematocyst capsules in Hydra. J. Cell Sci. 2002, 115, 745–751. [Google Scholar] [CrossRef] [PubMed]
- Weber, J. The development of cnidarian stinging cells: Maturation and migration of stenoteles of Hydra vulgaris. Roux’s Arch. Dev. Biol. 1995, 205, 171–181. [Google Scholar] [CrossRef]
- Okamura, B.; Gruhl, A.; Bartholomew, J.L. Myxozoan Evolution, Ecology and Development; Springer: Cham, Switzerland, 2015. [Google Scholar]
- Schmid-Hempel, P. Immune defence, parasite evasion strategies and their relevance for ‘macroscopic phenomena’ such as virulence. Phil. Trans. R. Soc. B 2009, 364, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Yazdanbakhsh, M.; Sacks, D.L. Why does immunity to parasites take so long to develop? Nat. Rev. Immunol. 2010, 10, 80–81. [Google Scholar] [CrossRef] [Green Version]
- Maizels, R.M. Parasite immunomodulation and polymorphisms of the immune system. J. Biol. 2009, 8, 62. [Google Scholar] [CrossRef]
- Gibieža, P.; Petrikaitė, V. The regulation of actin dynamics during cell division and malignancy. Am. J. Cancer Res. 2021, 11, 4050–4069. [Google Scholar]
- Brekhman, V.; Ofek-lalzar, M.; Atkinson, S.D.; Alama-bermejo, G.; Maor-landaw, K.; Malik, A.; Bartholomew, J.L.; Lotan, T. Proteomic analysis of the parasitic cnidarian Ceratonova shasta (Cnidaria:Myxozoa) reveals diverse roles of actin in motility and spore formation. Front. Mar. Sci. 2021, 8, 632700. [Google Scholar] [CrossRef]
- Alama-Bermejo, G.; Holzer, A.S.; Bartholomew, J.L. Myxozoan adhesion and virulence: Ceratonova shasta on the move. Microorganisms 2019, 7, 397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hechinger, R.F.; Lafferty, K.D.; Kuris, A.M. Parasites. In Metabolic Ecology: A Scaling Approach; Sibly, R.M., Brown, J.H., Kodric-Brown, A., Eds.; Wiley-Blackwell: Oxford, UK, 2012; pp. 234–247. [Google Scholar]
- Van Niekerk, D.D.; Penkler, G.P.; Du Toit, F.; Snoep, J.L. Targeting glycolysis in the malaria parasite Plasmodium falciparum. FEBS J. 2016, 283, 634–646. [Google Scholar] [CrossRef] [Green Version]
- Rattray, A.M.J.; Müller, B. The control of histone gene expression. Biochem. Soc. Trans. 2012, 40, 880–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, G.S.; Stein, J.L. Is human histone gene expression autogenously regulated? Mol. Cell. Biochem. 1984, 64, 105–110. [Google Scholar] [CrossRef]
- Stein, G.S.; Baumbach, L.L.; Stein, J.L. Regulation of human histone gene expression: Transcriptional and posttranscriptional control in the coupling of histone messenger RNA stability with DNA replication. Biochemistry 1987, 26, 6178–6187. [Google Scholar]
- Miller, D.J.; Harrison, P.L.; Mahony, T.J.; Mcmillan, J.P.; Miles, A.; Odorico, D.M.; Lohuis, M.R. Nucleotide sequence of the histone gene cluster in the coral Acropora formosa (Cnidaria; Scleractinia): Features of histone gene structure and organization are common to diploblastic and triploblastic metazoans. J. Mol. Evol. 1993, 37, 245–253. [Google Scholar] [CrossRef]
- Török, A.; Schiffer, P.H.; Schnitzler, C.E.; Ford, K.; Mullikin, J.C.; Baxevanis, A.D.; Bacic, A.; Frank, U.; Gornik, S.G. The cnidarian Hydractinia echinata employs canonical and highly adapted histones to pack its DNA. Epigenetics Chromatin 2016, 9, 36. [Google Scholar] [CrossRef] [Green Version]
- Reddy, P.C.; Ubhe, S.; Sirwani, N.; Lohokare, R.; Galande, S. Rapid divergence of histones in Hydrozoa (Cnidaria) and evolution of a novel histone involved in DNA damage response in hydra. Zoology 2017, 123, 53–63. [Google Scholar] [CrossRef]
- Piriatinskiy, G.; Atkinson, S.D.; Park, S.; Morgenstern, D.; Yossifon, G.; Bartholomew, J.L.; Lotan, T. Functional and proteomic analysis of Ceratonova shasta (Cnidaria: Myxozoa) polar capsules reveals adaptations to parasitism. Sci. Rep. 2017, 7, 9010. [Google Scholar] [CrossRef] [Green Version]
- Kumar, G.; Gotesman, M.; El-Matbouli, M. Interaction of Tetracapsuloides bryosalmonae, the causative agent of proliferative kidney disease, with host proteins in the kidney of Salmo trutta. Parasitol. Res. 2015, 114, 1721–1727. [Google Scholar] [CrossRef] [Green Version]
- Fall, M.; Cosette, P.; Hardouin, J.; Toguebaye, B.; Cornillot, E. A proteome study of Myxobolus episquamalis parasite of flathead mullet (Mugil cephalus Linné 1758) of Senegalese coasts. Rev. Cames 2017, 5, 6–12. [Google Scholar]
- Miao, J.; Fan, Q.; Cui, L.; Li, J.; Li, J.; Cui, L. The malaria parasite Plasmodium falciparum histones: Organization, expression, and acetylation. Gene 2006, 369, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Soto, M.; Requena, J.M.; Quijada, L.; Alonso, C. Organization, transcription and regulation of the Leishmania infantum histone H3 genes. Biochem. J. 1996, 318, 813–819. [Google Scholar] [CrossRef] [Green Version]
- García-Salcedo, J.A.; Oliver, J.L.; Stock, R.P.; González, A. Molecular characterization and transcription of the histone H2B gene from the protozoan parasite Trypanosoma cruzi. Mol. Microbiol. 1994, 13, 1033–1043. [Google Scholar] [CrossRef]
- Espinoza, I.; Gomez, C.R.; Galindo, M.; Galanti, N. Developmental expression pattern of histone H4 gene associated to DNA synthesis in the endoparasitic platyhelminth Mesocestoides corti. Gene 2007, 386, 35–41. [Google Scholar] [CrossRef]
- Ahmad, F.; Debes, P.V.; Pukk, L.; Kahar, S.; Hartikainen, H.; Gross, R.; Vasemägi, A. Know your enemy—Transcriptome of myxozoan Tetracapsuloides bryosalmonae reveals potential drug targets against proliferative kidney disease in salmonids. Parasitology 2021, 148, 726–739. [Google Scholar] [CrossRef]
- Coakley, G.; Maizels, R.M.; Buck, A.H. Exosomes and other extracellular vesicles: The new communicators in parasite infections. Trends Parasitol. 2015, 31, 477–489. [Google Scholar] [CrossRef] [Green Version]
- Marshall, S.; Kelly, P.H.; Singh, B.K.; Pope, R.M.; Kim, P.; Zhanbolat, B.; Wilson, M.E.; Yao, C. Extracellular release of virulence factor major surface protease via exosomes in Leishmania infantum promastigotes. Parasites Vectors 2018, 11, 355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartigan, A.; Kosakyan, A.; Pecková, H.; Eszterbauer, E.; Holzer, A.S. Transcriptome of Sphaerospora molnari (Cnidaria, Myxosporea) blood stages provides proteolytic arsenal as potential therapeutic targets against sphaerosporosis in common carp. BMC Genomics 2020, 21, 404. [Google Scholar] [CrossRef]
- Alama-Bermejo, G.; Bartošová-Sojková, P.; Atkinson, S.D.; Holzer, A.S.; Bartholomew, J.L. Proteases as therapeutic targets against the parasitic cnidarian Ceratonova shasta: Characterization of molecules key to parasite virulence in Salmonid hosts. Front. Cell. Infect. Microbiol. 2022, 11, 804864. [Google Scholar] [CrossRef] [PubMed]
- Faber, M.; Shaw, S.; Yoon, S.; de Paiva Alves, E.; Wang, B.; Qi, Z.; Okamura, B.; Hartikainen, H.; Secombes, C.J.; Holland, J.W. Comparative transcriptomics and host-specific parasite gene expression profiles inform on drivers of proliferative kidney disease. Sci. Rep. 2021, 11, 2149. [Google Scholar] [CrossRef] [PubMed]
- Kelley, G.O.; Adkison, M.A.; Leutenegger, C.M.; Hedrick, R.P. Myxobolus cerebralis: Identification of a cathepsin Z-like protease gene (Myx CP-1) expressed during parasite development in rainbow trout, Oncorhynchus mykiss. Exp. Parasitol. 2003, 105, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Almagro Armenteros, J.J.; Sønderby, C.K.; Sønderby, S.K.; Nielsen, H.; Winther, O. DeepLoc: Prediction of protein subcellular localization using deep learning. Bioinformatics 2017, 33, 3387–3395. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, V.; Valck, C.; Rojas, Á.; Aguilar, L.; Ramirez, G.; Schwaeble, W.; Ferreira, A. Role of calreticulin from parasites in its interaction with vertebrate hosts. Mol. Immunol. 2004, 40, 1279–1291. [Google Scholar] [CrossRef]
- Moreau, C.; Cioci, G.; Iannello, M.; Laffly, E.; Chouquet, A.; Ferreira, A.; Thielens, N.M.; Gaboriaud, C. Structures of parasite calreticulins provide insights into their flexibility and dual carbohydrate/peptide-binding properties. IUCrJ 2016, 3, 408–419. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Toloza, G.; Aguilar-Guzmán, L.; Valck, C.; Ferreira, V.P.; Ferreira, A. The Interactions of parasite calreticulin with initial complement components: Consequences in immunity and virulence. Front. Immunol. 2020, 11, 1561. [Google Scholar] [CrossRef]
- Wang, W.A.; Groenendyk, J.; Michalak, M. Calreticulin signaling in health and disease. Int. J. Biochem. Cell Biol. 2012, 44, 842–846. [Google Scholar] [CrossRef]
- Michalak, M.; Corbett, E.F.; Mesaeli, N.; Nakamura, K.; Opas, M. Calreticulin: One protein, one gene, many functions. Biochem. J. 1999, 344 Pt 2, 281–292. [Google Scholar] [CrossRef]
- Esperante, D.; Flisser, A.; Mendlovic, F. The many faces of parasite calreticulin. Front. Immunol. 2023, 14, 1101390. [Google Scholar] [CrossRef]
- Holland, J.W.; Holzer, A.S. Myxozoan Research Forum 2021—The ‘MyxoMixer’: Advances, methods, and problems yet to be solved in myxozoan research. Bull. Eur. Assoc. Fish Pathol. 2021, 41, 216–242. [Google Scholar] [CrossRef]
- Avilán, L.; Gualdrón-López, M.; Quiñones, W.; González-González, L.; Hannaert, V.; Michels, P.A.M.; Concepción, J.L. Enolase: A key player in the metabolism and a probable virulence factor of trypanosomatid parasites—Perspectives for its use as a therapeutic target. Enzyme Res. 2011, 2011, 932549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Arreaza, A.; Acosta, H.; Quiñones, W.; Concepción, J.L.; Michels, P.A.M.; Avilán, L. Extracellular functions of glycolytic enzymes of parasites: Unpredicted use of ancient proteins. Mol. Biochem. Parasitol. 2014, 193, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Wang, J.; Guo, J.; Li, Y.; Lian, S.; Guo, W.; Yang, H.; Kong, F.; Zhen, L.; Guo, L.; et al. Progress in the biological function of alpha-enolase. Anim. Nutr. 2016, 2, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Moran, Y.; Agron, M.; Praher, D.; Technau, U. The evolutionary origin of plant and animal microRNAs. Nat. Ecol. Evol. 2017, 1, 0027. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function genomics: The miRNA genes. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Hakimi, M.-a.; Cannella, D. Apicomplexan parasites and subversion of the host cell microRNA pathway. Trends Parasitol. 2011, 27, 481–486. [Google Scholar] [CrossRef]
- Nicolas, F.E.; Torres-martinez, S.; Ruiz-vazquez, R.M. Loss and retention of RNA interference in fungi and parasites. PLoS Pathog. 2013, 9, e1003089. [Google Scholar] [CrossRef] [Green Version]
- Britton, C.; Laing, R.; Devaney, E. Small RNAs in parasitic nematodes—Forms and functions. Parasitology 2020, 147, 855–864. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Cai, X.; Bradley, J.E. MicroRNAs in parasites and parasite infection. RNA Biol. 2013, 10, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Moss, E.G.; Tang, L. Conservation of the heterochronic regulator Lin-28, its developmental expression and microRNA complementary sites. Dev. Biol. 2003, 258, 432–442. [Google Scholar] [CrossRef]
- Nunes, C.C.; Gowda, M.; Sailsbery, J.; Xue, M.; Chen, F.; Brown, D.E.; Oh, Y.Y.; Mitchell, T.K.; Dean, R.A. Diverse and tissue-enriched small RNAs in the plant pathogenic fungus, Magnaporthe oryzae. BMC Genomics 2011, 12, 288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarrant, A. Ecology and physiology of dormancy in a changing world: Introduction to a virtual symposium in The Biological Bulletin. Biol. Bull. 2019, 237, 73–75. [Google Scholar] [CrossRef] [PubMed]
- Weiberg, A.; Wang, M.; Lin, F.M.; Zhao, H.; Zhang, Z.; Kaloshian, I.; Huang, H.D.; Jin, H. Fungal small RNAs suppress plant immunity by hijacking host RNA interference pathways. Science 2013, 342, 118–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Weiberg, A.; Lin, F.M.; Thomma, B.P.H.J.; Huang, H.D.; Jin, H. Bidirectional cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection. Nat. Plants 2016, 2, 16151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buck, A.H.; Coakley, G.; Simbari, F.; McSorley, H.J.; Quintana, J.F.; Le Bihan, T.; Kumar, S.; Abreu-Goodger, C.; Lear, M.; Harcus, Y.; et al. Exosomes secreted by nematode parasites transfer small RNAs to mammalian cells and modulate innate immunity. Nat. Commun. 2014, 5, 5488. [Google Scholar] [CrossRef] [Green Version]
- Quintana, J.F.; Makepeace, B.L.; Babayan, S.A.; Ivens, A.; Pfarr, K.M.; Blaxter, M.; Debrah, A.; Wanji, S.; Ngangyung, H.F.; Bah, G.S.; et al. Extracellular Onchocerca-derived small RNAs in host nodules and blood. Parasites Vectors 2015, 8, 58. [Google Scholar] [CrossRef] [Green Version]
- Zamanian, M.; Fraser, L.M.; Agbedanu, P.N.; Harischandra, H.; Moorhead, A.R.; Day, T.A.; Bartholomay, L.C.; Kimber, M.J. Release of small RNA-containing exosome-like vesicles from the human filarial parasite Brugia malayi. PLoS Negl. Trop. Dis. 2015, 9, e0004069. [Google Scholar] [CrossRef] [Green Version]
- Cheng, G.; Luo, R.; Hu, C.; Cao, J.; Jin, Y. Deep sequencing-based identification of pathogen-specific microRNAs in the plasma of rabbits infected with Schistosoma japonicum. Parasitology 2013, 140, 1751–1761. [Google Scholar] [CrossRef]
- Kim, V.N.; Han, J.; Siomi, M.C. Biogenesis of small RNAs in animals. Nat. Rev. Mol. Cell Biol. 2009, 10, 126–139. [Google Scholar] [CrossRef]
- Siomi, M.C.; Kuramochi-Miyagawa, S. RNA silencing in germlines-exquisite collaboration of Argonaute proteins with small RNAs for germline survival. Curr. Opin. Cell Biol. 2009, 21, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Silva, M.R.; Cura Das Neves, R.F.; Cabrera-Cabrera, F.; Sanguinetti, J.; Medeiros, L.C.; Robello, C.; Naya, H.; Fernandez-Calero, T.; Souto-Padron, T.; De Souza, W.; et al. Extracellular vesicles shed by Trypanosoma cruzi are linked to small RNA pathways, life cycle regulation, and susceptibility to infection of mammalian cells. Parasitol. Res. 2014, 113, 285–304. [Google Scholar] [CrossRef] [PubMed]
- Degnan, B.M.; Vervoort, M.; Larroux, C.; Richards, G.S. Early evolution of metazoan transcription factors. Curr. Opin. Genet. Dev. 2009, 19, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Putnam, N.H.; Srivastava, M.; Hellsten, U.; Dirks, B.; Chapman, J.; Salamov, A.; Terry, A.; Shapiro, H.; Lindquist, E.; Kapitonov, V.V.; et al. Sea anemone genome reveals ancestral eumetazoan gene repertoire and genomic organization. Science 2007, 317, 86–94. [Google Scholar] [CrossRef] [Green Version]
- De Mendoza, A.; Sebé-Pedrós, A.; Šestak, M.S.; Matejčić, M.; Torruella, G.; Domazet-Lošo, T.; Ruiz-Trillo, I. Transcription factor evolution in eukaryotes and the assembly of the regulatory toolkit in multicellular lineages. Proc. Natl. Acad. Sci. USA 2013, 110, E4858–E4866. [Google Scholar] [CrossRef]
- Brekhman, V.; Malik, A.; Haas, B.; Sher, N.; Lotan, T. Transcriptome profiling of the dynamic life cycle of the scypohozoan jellyfish Aurelia aurita. BMC Genom. 2015, 16, 74. [Google Scholar] [CrossRef] [Green Version]
- Gehring, W.J. The homeobox in perspective. Trends Biochem. Sci. 1992, 17, 277–280. [Google Scholar] [CrossRef]
- Malarkey, C.S.; Churchill, M.E.A. The high mobility group box: The ultimate utility player of a cell. Trends Biochem. Sci. 2015, 37, 553–562. [Google Scholar] [CrossRef] [Green Version]
- Vaquerizas, J.M.; Kummerfeld, S.K.; Teichmann, S.A.; Luscombe, N.M. A census of human transcription factors: Function, expression and evolution. Nat. Rev. Genet. 2009, 10, 252–263. [Google Scholar] [CrossRef]
- Neverov, A.M.; Panchin, A.Y.; Mikhailov, K.V.; Batueva, M.D.; Aleoshin, V.V.; Panchin, Y.V. Apoptotic gene loss in Cnidaria is associated with transition to parasitism. Sci. Rep. 2023, 13, 8015. [Google Scholar] [CrossRef]
- Leclère, L.; Horin, C.; Chevalier, S.; Lapébie, P.; Dru, P.; Peron, S.; Jager, M.; Condamine, T.; Pottin, K.; Romano, S.; et al. The genome of the jellyfish Clytia hemisphaerica and the evolution of the cnidarian life-cycle. Nat. Ecol. Evol. 2019, 3, 801–810. [Google Scholar] [CrossRef] [Green Version]
- Jager, M.; Quéinnec, E.; Guyader, H.L.; Manuel, M. Multiple Sox genes are expressed in stem cells or in differentiating neuro-sensory cells in the hydrozoan Clytia hemisphaerica. Evodevo 2011, 2, 12. [Google Scholar] [CrossRef] [Green Version]
- Magie, C.R.; Pang, K.; Martindale, M.Q. Genomic inventory and expression of Sox and Fox genes in the cnidarian Nematostella vectensis. Dev. Genes Evol. 2005, 215, 618–630. [Google Scholar] [CrossRef]
- Shinzato, C.; Iguchi, A.; Hayward, D.C.; Technau, U.; Ball, E.E.; Miller, D.J. Sox genes in the coral Acropora millepora: Divergent expression patterns reflect differences in developmental mechanisms within the Anthozoa. BMC Evol. Biol. 2008, 8, 311. [Google Scholar] [CrossRef] [Green Version]
- Kyslík, J.; Vancová, M.; Bartošová-sojková, P.; Lövy, A.; Holzer, A.S.; Fiala, I. Expression profiling and cellular localization of myxozoan minicollagens during nematocyst formation and sporogenesis. Int. J. Parasitol. 2022, 52, 667–675. [Google Scholar] [CrossRef]
- Poulin, R.; Randhawa, H.S. Evolution of parasitism along convergent lines: From ecology to genomics. Parasitology 2015, 142, S6–S15. [Google Scholar] [CrossRef] [PubMed]
- Sklarz, M.; Levin, L.; Gordon, M.; Chalifa-caspi, V. NeatSeq-Flow: A lightweight high-throughput sequencing workflow platform for non-programmers and programmers alike. bioRxiv 2018. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopylova, E.; Noé, L.; Touzet, H. SortMeRNA: Fast and accurate filtering of ribosomal RNAs in metatranscriptomic data. Bioinformatics 2012, 28, 3211–3217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menzel, P.; Ng, K.L.; Krogh, A. Fast and sensitive taxonomic classification for metagenomics with Kaiju. Nat. Commun. 2016, 7, 11257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Simão, F.A.; Zdobnov, E.M. BUSCO update: Novel and streamlined workflows along with broader and deeper phylogenetic coverage for scoring of eukaryotic, prokaryotic, and viral genomes. Mol. Biol. Evol. 2021, 38, 4647–4654. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinformatics 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryant, D.M.; Johnson, K.; DiTommaso, T.; Tickle, T.; Couger, M.B.; Payzin-Dogru, D.; Lee, T.J.; Leigh, N.D.; Kuo, T.H.; Davis, F.G.; et al. A tissue-mapped axolotl de novo transcriptome enables identification of limb regeneration factors. Cell Rep. 2017, 18, 762–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [Green Version]
- Afgan, E.; Sloggett, C.; Goonasekera, N.; Makunin, I.; Benson, D.; Crowe, M.; Gladman, S.; Kowsar, Y.; Pheasant, M.; Horst, R.; et al. Genomics Virtual Laboratory: A practical bioinformatics workbench for the cloud. PLoS ONE 2015, 10, e0140829. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. ClusterProfiler: An R package for comparing biological themes among gene clusters. Omics J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein—Protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y.; Kawashima, M. KEGG mapping tools for uncovering hidden features in biological data. Protein Sci. 2022, 31, 47–53. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y. KEGG Mapper for inferring cellular functions from protein sequences. Protein Sci. 2020, 29, 28–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voolstra, C.R.; Miller, D.J.; Ragan, M.A.; Hoffmann, A.A.; Hoegh-Guldberg, O.; Bourne, D.G.; Ball, E.E.; Ying, H.; Forêt, S.; Takahashi, S.; et al. The ReFuGe 2020 Consortium-using “omics” approaches to explore the adaptability and resilience of coral holobionts to environmental change. Front. Mar. Sci. 2015, 2, 68. [Google Scholar]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maor-Landaw, K.; Avidor, I.; Rostowsky, N.; Salti, B.; Smirnov, M.; Ofek-Lalzar, M.; Levin, L.; Brekhman, V.; Lotan, T. The Molecular Mechanisms Employed by the Parasite Myxobolus bejeranoi (Cnidaria: Myxozoa) from Invasion through Sporulation for Successful Proliferation in Its Fish Host. Int. J. Mol. Sci. 2023, 24, 12824. https://doi.org/10.3390/ijms241612824
Maor-Landaw K, Avidor I, Rostowsky N, Salti B, Smirnov M, Ofek-Lalzar M, Levin L, Brekhman V, Lotan T. The Molecular Mechanisms Employed by the Parasite Myxobolus bejeranoi (Cnidaria: Myxozoa) from Invasion through Sporulation for Successful Proliferation in Its Fish Host. International Journal of Molecular Sciences. 2023; 24(16):12824. https://doi.org/10.3390/ijms241612824
Chicago/Turabian StyleMaor-Landaw, Keren, Itamar Avidor, Nadav Rostowsky, Barbara Salti, Margarita Smirnov, Maya Ofek-Lalzar, Liron Levin, Vera Brekhman, and Tamar Lotan. 2023. "The Molecular Mechanisms Employed by the Parasite Myxobolus bejeranoi (Cnidaria: Myxozoa) from Invasion through Sporulation for Successful Proliferation in Its Fish Host" International Journal of Molecular Sciences 24, no. 16: 12824. https://doi.org/10.3390/ijms241612824
APA StyleMaor-Landaw, K., Avidor, I., Rostowsky, N., Salti, B., Smirnov, M., Ofek-Lalzar, M., Levin, L., Brekhman, V., & Lotan, T. (2023). The Molecular Mechanisms Employed by the Parasite Myxobolus bejeranoi (Cnidaria: Myxozoa) from Invasion through Sporulation for Successful Proliferation in Its Fish Host. International Journal of Molecular Sciences, 24(16), 12824. https://doi.org/10.3390/ijms241612824