
Multigenerational Effects of Graphene Oxide Nanoparticles on Acheta domesticus DNA Stability

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
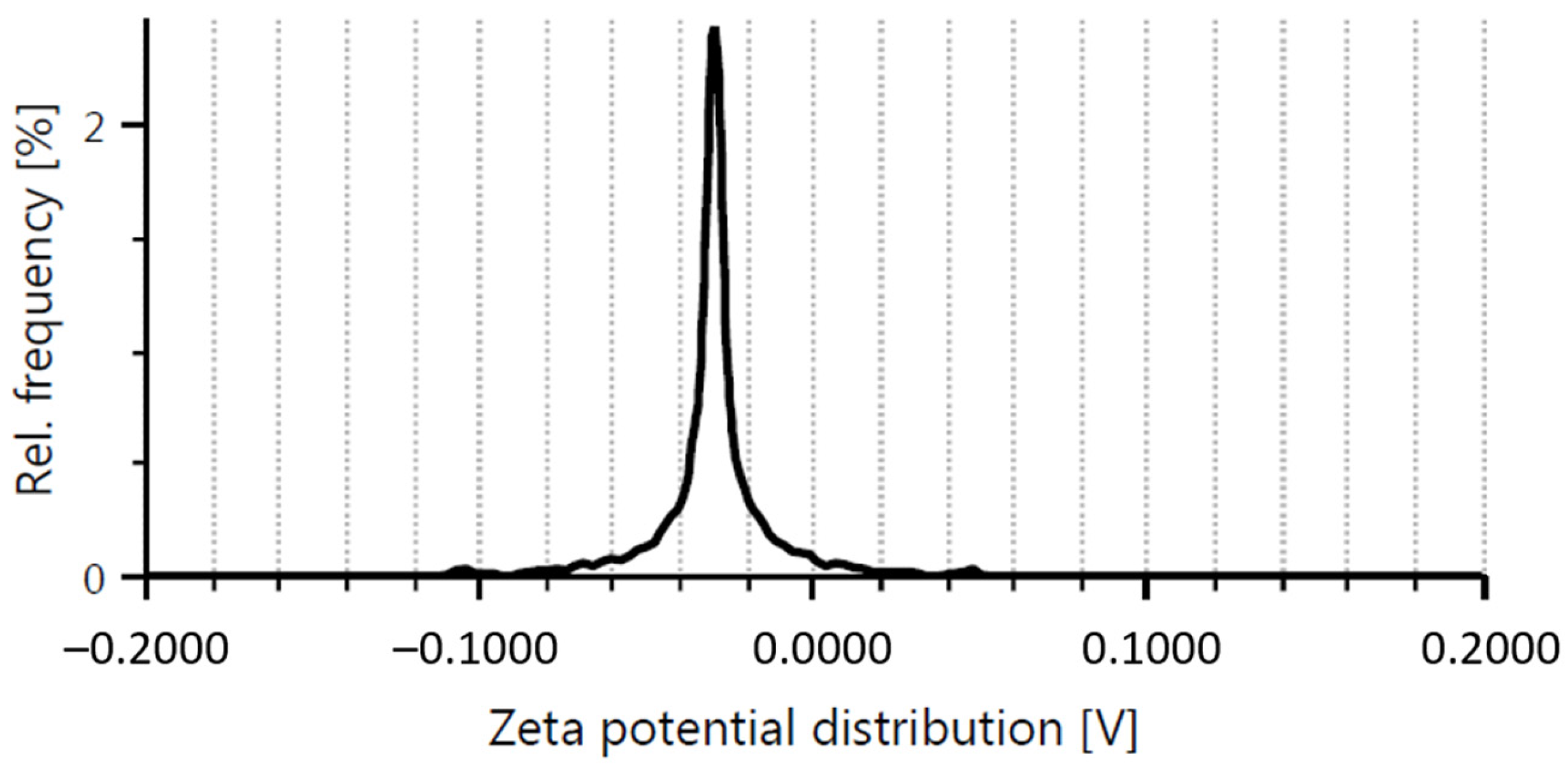
2.1. Graphene Oxide
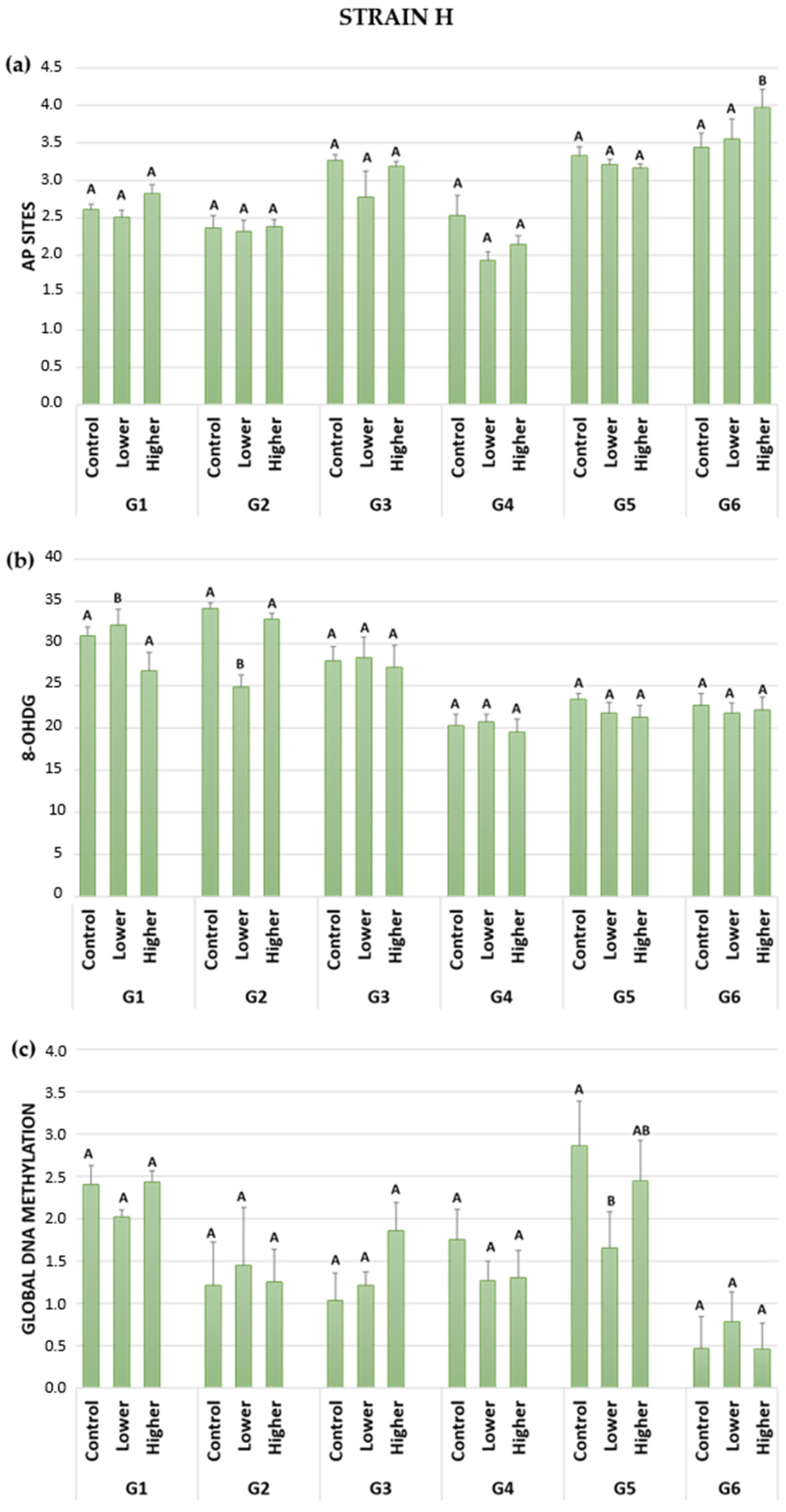
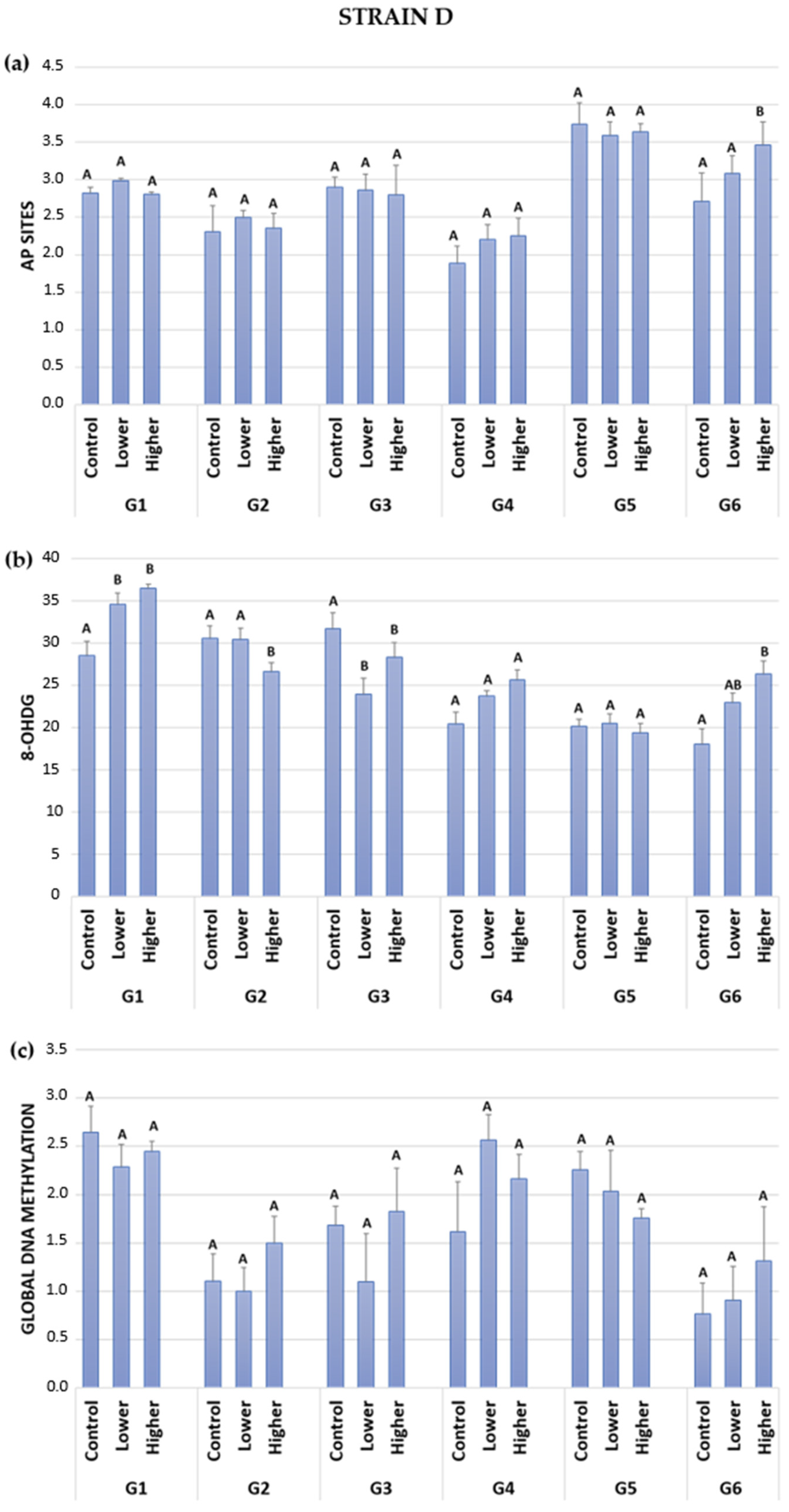
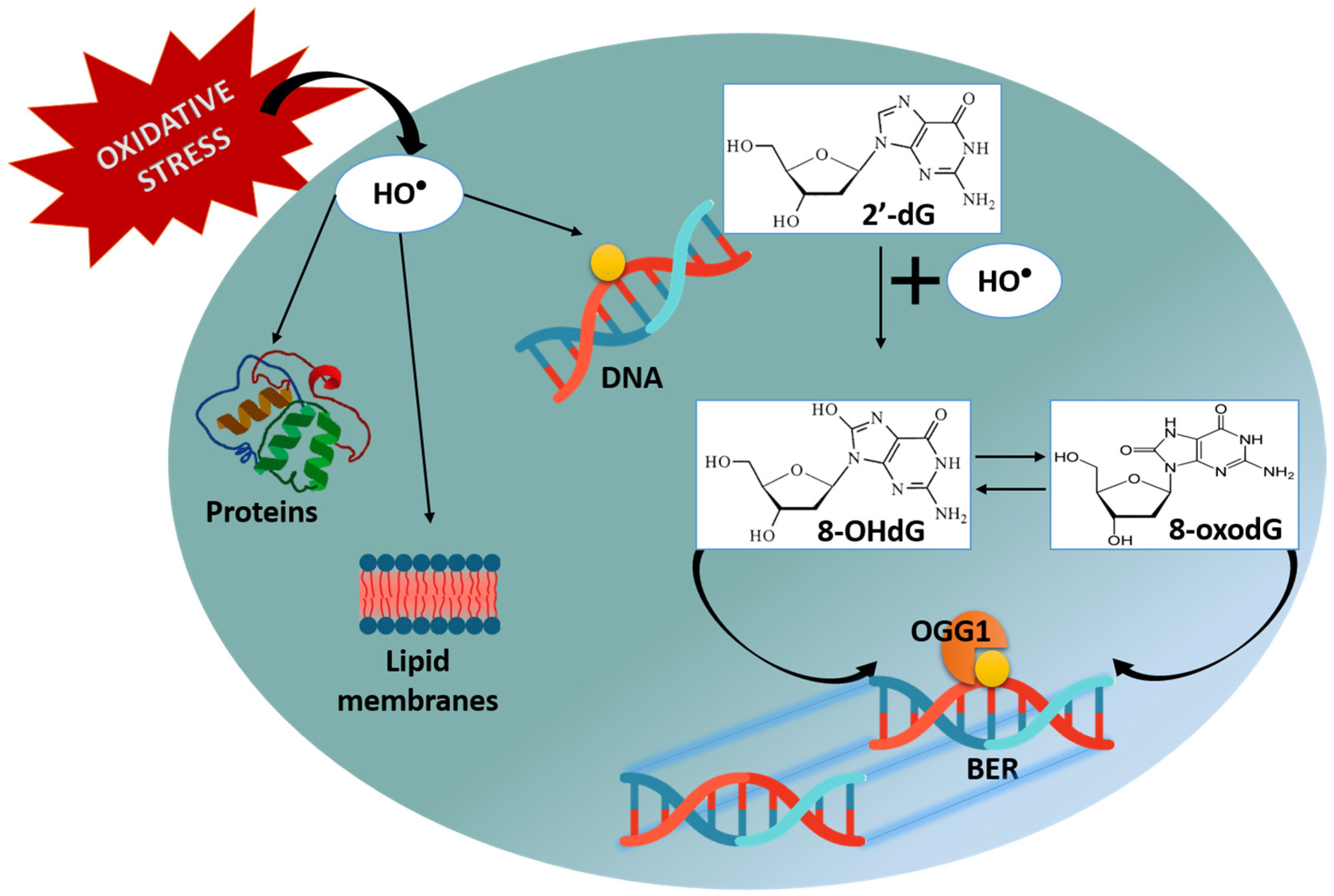
2.2. DNA Damage: AP Sites (Apurinic/Apyrimidinic Sites)
2.3. DNA Damage: 8-OHdG (8-hydroxy-2′-deoxyguanosine)
2.4. Global DNA Methylation
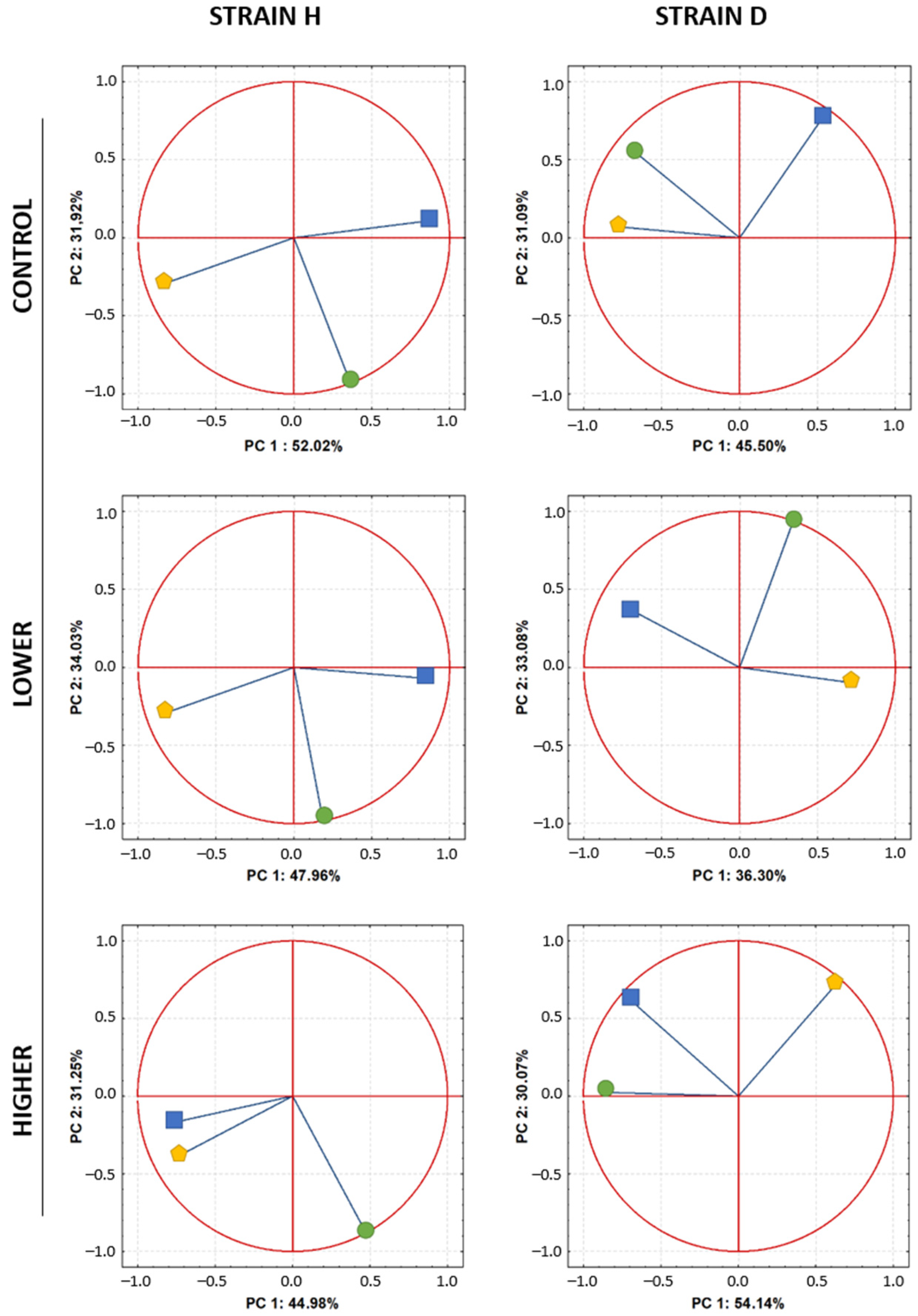
2.5. PCA—Relationships between Variables
3. Discussion
4. Materials and Methods
4.1. Graphene Oxide Characteristics
4.2. Characteristics of the Species
4.3. Food Preparation: Graphene Oxide, Control Food
4.4. Experimental Model and Tissue Preparation
4.5. DNA Isolation
4.6. Measurement of Selected Parameters
4.6.1. AP Sites (Apurinic/Apyrimidinic Sites)
4.6.2. 8-OHdG (8-hydroxy-2′-deoxyguanosine)
4.6.3. Global DNA Methylation
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brisebois, P.P.; Siaj, M. Harvesting Graphene Oxide-Years 1859 to 2019: A Review of Its Structure, Synthesis, Properties and Exfoliation. J. Mater. Chem. C Mater. 2020, 8, 1517–1547. [Google Scholar] [CrossRef]
- Smith, A.T.; LaChance, A.M.; Zeng, S.; Liu, B.; Sun, L. Synthesis, Properties, and Applications of Graphene Oxide/Reduced Graphene Oxide and Their Nanocomposites. Nano Mater. Sci. 2019, 1, 31–47. [Google Scholar] [CrossRef]
- Aliyev, E.; Filiz, V.; Khan, M.M.; Lee, Y.J.; Abetz, C.; Abetz, V. Structural Characterization of Graphene Oxide: Surface Functional Groups and Fractionated Oxidative Debris. Nanomaterials 2019, 9, 1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukhanova, A.; Bozrova, S.; Sokolov, P.; Berestovoy, M.; Karaulov, A.; Nabiev, I. Dependence of Nanoparticle Toxicity on Their Physical and Chemical Properties. Nanoscale Res. Lett. 2018, 13, 44. [Google Scholar] [CrossRef] [Green Version]
- Markovic, M.; Kumar, A.; Andjelkovic, I.; Lath, S.; Kirby, J.K.; Losic, D.; Batley, G.E.; McLaughlin, M.J. Ecotoxicology of Manufactured Graphene Oxide Nanomaterials and Derivation of Preliminary Guideline Values for Freshwater Environments. Environ. Toxicol. Chem. 2018, 37, 1340–1348. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Lu, F.; Tu, Y.; Ren, Z. Glucose Biosensors Based on Carbon Nanotube Nanoelectrode Ensembles. Nano Lett. 2004, 4, 191–195. [Google Scholar] [CrossRef]
- Timur, S.; Anik, U.; Odaci, D.; Gorton, L. Development of a Microbial Biosensor Based on Carbon Nanotube (CNT) Modified Electrodes. Electrochem. Commun. 2007, 9, 1810–1815. [Google Scholar] [CrossRef]
- Maehashi, K.; Katsura, T.; Kerman, K.; Takamura, Y.; Matsumoto, K.; Tamiya, E. Label-Free Protein Biosensor Based on Aptamer-Modified Carbon Nanotube Field-Effect Transistors. Anal. Chem. 2007, 79, 782–787. [Google Scholar] [CrossRef] [PubMed]
- Flasz, B.; Dziewięcka, M.; Kędziorski, A.; Tarnawska, M.; Augustyniak, M. Vitellogenin Expression, DNA Damage, Health Status of Cells and Catalase Activity in Acheta Domesticus Selected According to Their Longevity after Graphene Oxide Treatment. Sci. Total Environ. 2020, 737, 140274. [Google Scholar] [CrossRef]
- Flasz, B.; Dziewięcka, M.; Kędziorski, A.; Tarnawska, M.; Augustyniak, J.; Augustyniak, M. Multigenerational Selection towards Longevity Changes the Protective Role of Vitamin C against Graphene Oxide-Induced Oxidative Stress in House Crickets. Environ. Pollut. 2021, 290, 117996. [Google Scholar] [CrossRef]
- Dziewięcka, M.; Flasz, B.; Rost Roszkowska, M.; Kędziorski, A.; Kochanowicz, A.; Augustyniak, M. Graphene Oxide as a New Anthropogenic Stress Factor Multigenerational Study at the Molecular, Cellular, Individual and Population Level of Acheta Domesticus. J. Hazard. Mater. 2020, 396, 122775. [Google Scholar] [CrossRef] [PubMed]
- Cano, A.M.; Maul, J.D.; Saed, M.; Shah, S.A.; Green, M.J.; Cañas-Carrell, J.E. Bioaccumulation, Stress, and Swimming Impairment in Daphnia Magna Exposed to Multiwalled Carbon Nanotubes, Graphene, and Graphene Oxide. Environ. Toxicol. Chem. 2017, 36, 2199–2204. [Google Scholar] [CrossRef] [PubMed]
- Souza, J.P.; Venturini, F.P.; Santos, F.; Zucolotto, V. Chronic Toxicity in Ceriodaphnia Dubia Induced by Graphene Oxide. Chemosphere 2018, 190, 218–224. [Google Scholar] [CrossRef]
- Lv, X.; Yang, Y.; Tao, Y.; Jiang, Y.; Chen, B.; Zhu, X.; Cai, Z.; Li, B. A Mechanism Study on Toxicity of Graphene Oxide to Daphnia Magna: Direct Link between Bioaccumulation and Oxidative Stress. Environ. Pollut. 2018, 234, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Liu, W. Chronic and Transgenerational Effects of Silver Nanoparticles in Freshwater Gastropod Lymnaea Stagnalis. Chemosphere 2023, 313, 137386. [Google Scholar] [CrossRef] [PubMed]
- McGillicuddy, E.; Murray, I.; Kavanagh, S.; Morrison, L.; Fogarty, A.; Cormican, M.; Dockery, P.; Prendergast, M.; Rowan, N.; Morris, D. Silver Nanoparticles in the Environment: Sources, Detection and Ecotoxicology. Sci. Total Environ. 2017, 575, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Tiede, K.; Boxall, A.B.A.; Tear, S.P.; Lewis, J.; David, H.; Hassellöv, M. Detection and Characterization of Engineered Nanoparticles in Food and the Environment. Food Addit. Contam. Part A 2008, 25, 795–821. [Google Scholar] [CrossRef] [PubMed]
- Peters, R.J.B.; van Bemmel, G.; Milani, N.B.L.; den Hertog, G.C.T.; Undas, A.K.; van der Lee, M.; Bouwmeester, H. Detection of Nanoparticles in Dutch Surface Waters. Sci. Total Environ. 2018, 621, 210–218. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, J.; Cai, Z.; Feng, Y.; Wang, Y.; Zhang, D.; Pan, X. Environmental Science Nano CRITICAL REVIEW Detection of Engineered Nanoparticles in Aquatic Environments: Current Status and Challenges in Enrichment, Separation, and Analysis. Environ. Sci. Nano 2019, 6, 709. [Google Scholar] [CrossRef]
- Guimarães, B.; Römbke, J.; Amorim, M.J.B. On the Importance of Longer-Term Exposure to Stressors A Critical Review and Proposal for Multigenerational Testing in Standard Soil Invertebrates. Sci. Total Environ. 2023, 854, 158680. [Google Scholar] [CrossRef]
- Wamucho, A.; Unrine, J.M.; Kieran, T.J.; Glenn, T.C.; Schultz, C.L.; Farman, M.; Svendsen, C.; Spurgeon, D.J.; Tsyusko, O.V. Genomic Mutations after Multigenerational Exposure of Caenorhabditis Elegans to Pristine and Sulfidized Silver Nanoparticles. Environ. Pollut. 2019, 254, 113078. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, M.J.; Scott-Fordsmand, J.J.; Amorim, M.J.B. Multigenerational Exposure to Cobalt (CoCl2) and WCCo Nanoparticles in Enchytraeus Crypticus. Nanotoxicology 2019, 13, 751–760. [Google Scholar] [CrossRef]
- Kim, S.W.; Kwak, J.I.L.; An, Y.J. Multigenerational Study of Gold Nanoparticles in Caenorhabditis Elegans: Transgenerational Effect of Maternal Exposure. Environ. Sci. Technol. 2013, 47, 5393–5399. [Google Scholar] [CrossRef]
- Moon, J.; Kwak, J.I.L.; Kim, S.W.; An, Y.J. Multigenerational Effects of Gold Nanoparticles in Caenorhabditis Elegans: Continuous versus Intermittent Exposures. Environ. Pollut. 2017, 220, 46–52. [Google Scholar] [CrossRef]
- Luiz, M.; Da Silva, N.; Nogueira, D.J.; Köerich, J.S.; Pereira Vaz, V.; Justino, N.M.; Renato, J.; Schmidt, A.; Schulz Vicentini, D.; Matias, M.S.; et al. Environmental Toxicology Multigenerational Toxic Effects on Daphnia Magna Induced by Silver Nanoparticles and Glyphosate Mixture. Environ. Toxicol. Chem. 2021, 40, 1123–1131. [Google Scholar]
- Flasz, B.; Dziewięcka, M.; Kędziorski, A.; Tarnawska, M.; Augustyniak, M. Multigenerational Graphene Oxide Intoxication Results in Reproduction Disorders at the Molecular Level of Vitellogenin Protein Expression in Acheta Domesticus. Chemosphere 2021, 280, 130772. [Google Scholar] [CrossRef] [PubMed]
- Rossbach, L.M.; Oughton, D.H.; Maremonti, E.; Eide, D.M.; Brede, D.A. Impact of Multigenerational Exposure to AgNO3 or NM300K Ag NPs on Antioxidant Defense and Oxidative Stress in Caenorhabditis Elegans. Ecotoxicol. Environ. Saf. 2021, 216, 112178. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Wang, Q.; Rossi, L.; Ebbs, S.D.; White, J.C. Multigenerational Exposure to Cerium Oxide Nanoparticles: Physiological and Biochemical Analysis Reveals Transmissible Changes in Rapid Cycling Brassica Rapa. NanoImpact 2016, 1, 46–54. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Hou, J.; Zhu, Y.; Lin, D. Multigenerational Exposure to TiO2 Nanoparticles in Soil Stimulates Stress Resistance and Longevity of Survived C. Elegans via Activating Insulin/IGF-like Signaling. Environ. Pollut. 2020, 263, 114376. [Google Scholar] [CrossRef]
- Wamucho, A.; Heffley, A.; Tsyusko, O.V. Epigenetic Effects Induced by Silver Nanoparticles in Caenorhabditis Elegans after Multigenerational Exposure. Sci. Total Environ. 2020, 725, 138523. [Google Scholar] [CrossRef]
- Wamucho, A.; Unrine, J.; May, J.; Tsyusko, O. Global DNA Adenine Methylation in Caenorhabditis Elegans after Multigenerational Exposure to Silver Nanoparticles and Silver Nitrate. Int. J. Mol. Sci. 2023, 24, 6168. [Google Scholar] [CrossRef]
- Schultz, C.L.; Wamucho, A.; Tsyusko, O.V.; Unrine, J.M.; Crossley, A.; Svendsen, C.; Spurgeon, D.J. Multigenerational Exposure to Silver Ions and Silver Nanoparticles Reveals Heightened Sensitivity and Epigenetic Memory in Caenorhabditis Elegans. Proc. R. Soc. B Biol. Sci. 2016, 283, 20152911. [Google Scholar] [CrossRef] [PubMed]
- Bicho, R.C.; Roelofs, D.; Mariën, J.; Scott-Fordsmand, J.J.; Amorim, M.J.B. Epigenetic Effects of (Nano)Materials in Environmental Species Cu Case Study in Enchytraeus Crypticus. Environ. Int. 2020, 136, 105447. [Google Scholar] [CrossRef] [PubMed]
- Moreira, L.; Costa, C.; Pires, J.; Teixeira, J.P.; Fraga, S. How Can Exposure to Engineered Nanomaterials Influence Our Epigenetic Code? A Review of the Mechanisms and Molecular Targets. Mutat. Res. Rev. Mutat. Res. 2021, 788, 108385. [Google Scholar] [CrossRef] [PubMed]
- Dziewięcka, M.; Pawlyta, M.; Majchrzycki, Ł.; Balin, K.; Barteczko, S.; Czerkawska, M.; Augustyniak, M. The Structure–Properties–Cytotoxicity Interplay: A Crucial Pathway to Determining Graphene Oxide Biocompatibility. Int. J. Mol. Sci. 2021, 22, 5401. [Google Scholar] [CrossRef]
- de Juan, C.; Griffell, K.; Calbet, A.; Saiz, E. Multigenerational Physiological Compensation and Body Size Reduction Dampen the Effects of Warming on Copepods. Limnol. Ocean. 2023, 68, 1037–1047. [Google Scholar] [CrossRef]
- Furuta, Y.; Kobayashi, I. Mobility of DNA Sequence Recognition Domains in DNA Methyltransferases Suggests Epigenetics-Driven Adaptive Evolution. Mob. Genet. Elem. 2012, 2, 292–296. [Google Scholar] [CrossRef] [Green Version]
- Swanson, W.J. Adaptive Evolution of Genes and Gene Families. Curr. Opin. Genet. Dev. 2003, 13, 617–622. [Google Scholar] [CrossRef]
- Raimondo, S.; Hemmer, B.L.; Goodman, L.R.; Cripe, G.M. Multigenerational Exposure of the Estuarine Sheepshead Minnow (Cyprinodon Variegatus) to 17β-Estradiol. II. Population-Level Effects through Two Life Cycles. Environ. Toxicol. Chem. 2009, 28, 2409–2415. [Google Scholar] [CrossRef]
- Araujo, G.S.; Abessa, D.M.S.; Soares, A.M.V.M.; Loureiro, S. Multi-Generational Exposure to Pb in Two Monophyletic Daphnia Species: Individual, Functional and Population Related Endpoints. Ecotoxicol. Environ. Saf. 2019, 173, 77–85. [Google Scholar] [CrossRef]
- Narayan, R.K.J. Nuclear DNA Changes, Genome Differentiation and Evolution InNicotiana (Solanaceae). Plant Syst. Evol. 1987, 157, 161–180. [Google Scholar] [CrossRef]
- Kawecki, T.J.; Ebert, D. Conceptual Issues in Local Adaptation. Ecol. Lett. 2004, 7, 1225–1241. [Google Scholar] [CrossRef] [Green Version]
- Becks, L.; Betini, G.S.; Mcadam, A.G.; Griswold, C.K.; Norris, R. A Fitness Trade-off between Seasons Causes Multigenerational Cycles in Phenotype and Population Size. eLife 2017, 6, e18770. [Google Scholar]
- Augustyniak, M.; Tarnawska, M.; Dziewięcka, M.; Kafel, A.; Rost-Roszkowska, M.; Babczyńska, A. DNA Damage in Spodoptera Exigua after Multigenerational Cadmium Exposure A Trade-off between Genome Stability and Adaptation. Sci. Total Environ. 2020, 745, 141048. [Google Scholar] [CrossRef]
- Tu, Q.; Deng, J.; Di, M.; Lin, X.; Chen, Z.; Li, B.; Tian, L.; Zhang, Y. Reproductive Toxicity of Polystyrene Nanoplastics in Drosophila Melanogaster under Multi-Generational Exposure. Chemosphere 2023, 330, 138724. [Google Scholar] [CrossRef]
- Heinlaan, M.; Viljalo, K.; Richter, J.; Ingwersen, A.; Vija, H.; Mitrano, D.M. Multi-Generation Exposure to Polystyrene Nanoplastics Showed No Major Adverse Effects in Daphnia Magna. Environ. Pollut. 2023, 323, 121213. [Google Scholar] [CrossRef]
- Dziewięcka, M.; Witas, P.; Karpeta-Kaczmarek, J.; Kwaśniewska, J.; Flasz, B.; Balin, K.; Augustyniak, M. Reduced Fecundity and Cellular Changes in Acheta Domesticus after Multigenerational Exposure to Graphene Oxide Nanoparticles in Food. Sci. Total Environ. 2018, 635, 947–955. [Google Scholar] [CrossRef]
- Kim, S.W.; Moon, J.; An, Y.-J. Matricidal Hatching Can Induce Multi-Generational Effects in Nematode Caenorhabditis Elegans after Dietary Exposure to Nanoparticles. Environ. Sci. Pollut. Res. 2018, 25, 36394–36402. [Google Scholar] [CrossRef]
- Thompson, P.S.; Cortez, D. New Insights into Abasic Site Repair and Tolerance. DNA Repair 2020, 90, 102866. [Google Scholar] [CrossRef]
- Boiteux, S.; Guillet, M. Abasic Sites in DNA: Repair and Biological Consequences in Saccharomyces Cerevisiae. DNA Repair 2004, 3, 1–12. [Google Scholar] [CrossRef]
- Graille, M.; Wild, P.; Sauvain, J.J.; Hemmendinger, M.; Canu, I.G.; Hopf, N.B. Urinary 8-OHDG as a Biomarker for Oxidative Stress: A Systematic Literature Review and Meta-Analysis. Int. J. Mol. Sci. 2020, 21, 3743. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-Hydroxy-2′ -Deoxyguanosine (8-OHdG): A Critical Biomarker of Oxidative Stress and Carcinogenesis. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2009, 27, 120–139. [Google Scholar] [CrossRef] [Green Version]
- Friedberg, E.C. DNA Damage and Repair. Nature 2003, 421, 436–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scha, O.D.; Jiricny, J. Recent Progress in the Biology, Chemistry and Structural Biology of DNA Glycosylases. Bioessays 2001, 23, 270–281. [Google Scholar]
- Arndt, D.A.; Chen, J.; Moua, M.; Klaper, R.D. Multigeneration Impacts on Daphnia Magna of Carbon Nanomaterials with Differing Core Structures and Functionalizations. Environ. Toxicol. Chem. 2014, 33, 541–547. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Zhang, J.; Cheng, C.; Hou, K.; Wang, X.; Zhu, L.; Li, B.; Du, Z.; Wang, J.; Wang, J. Effects of Ecotoxicity of Penoxsulam Single and Co-Exposure with AgNPs on Eisenia Fetida. Chemosphere 2022, 307, 136134. [Google Scholar] [CrossRef]
- Gurunathan, S.; Kang, M.-H.; Jeyaraj, M.; Kim, J.-H. Differential Cytotoxicity of Different Sizes of Graphene Oxide Nanoparticles in Leydig (TM3) and Sertoli (TM4) Cells. Nanomaterials 2019, 9, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, R.; Liu, H.; Lu, C.; Hou, K.; Wang, X.; Wang, J.; Du, Z.; Li, B.; Zhu, L.; Wang, J. The Effect of TiO2NPs on Cloransulam-Methyl Toxicity to Earthworm (Eisenia Fetida). Chemosphere 2023, 322, 138242. [Google Scholar] [CrossRef]
- Elsaesser, A.; Howard, C.V. Toxicology of Nanoparticles. Adv. Drug Deliv. Rev. 2012, 64, 129–137. [Google Scholar] [CrossRef]
- Jastrzębska, A.M.; Olszyna, A.R. The Ecotoxicity of Graphene Family Materials: Current Status, Knowledge Gaps and Future Needs. J. Nanoparticle Res. 2015, 17, 40. [Google Scholar] [CrossRef]
- Jastrzębska, A.M.; Kurtycz, P.; Olszyna, A.R. Recent Advances in Graphene Family Materials Toxicity Investigations. J. Nanoparticle Res. 2012, 14, 1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmannova, D.; Borsky, P.; Svadlakova, T.; Borska, L.; Fiala, Z. Reproductive and Developmental Nanotoxicity of Carbon Nanoparticles. Nanomaterials 2022, 12, 1716. [Google Scholar] [CrossRef] [PubMed]
- Goutam Mukherjee, A.; Ramesh Wanjari, U.; Renu, K.; Vellingiri, B.; Valsala Gopalakrishnan, A. Heavy Metal and Metalloid Induced Reproductive Toxicity. Environ. Toxicol. Pharmacol. 2022, 92, 103859. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Feng, D.; Xu, Z.; Qi, P.; Yan, X. Acute Benzo[a]Pyrene Exposure Induced Oxidative Stress, Neurotoxicity and Epigenetic Change in Blood Clam Tegillarca Granosa. Sci. Rep. 2021, 11, 18744. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Dong, B.; Pang, N.; Hu, J. ROS Generation and DNA Damage Contribute to Abamectin-Induced Cytotoxicity in Mouse Macrophage Cells. Chemosphere 2019, 234, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Adeel, M.; Ma, C.; Ullah, S.; Rizwan, M.; Hao, Y.; Chen, C.; Jilani, G.; Shakoor, N.; Li, M.; Wang, L.; et al. Exposure to Nickel Oxide Nanoparticles Insinuates Physiological, Ultrastructural and Oxidative Damage: A Life Cycle Study on Eisenia Fetida. Environ. Pollut. 2019, 254, 113032. [Google Scholar] [CrossRef]
- Mizuta, Y.; Tokuda, K.; Guo, J.; Zhang, S.; Narahara, S.; Kawano, T.; Murata, M.; Yamaura, K.; Hoka, S.; Hashizume, M.; et al. Sodium Thiosulfate Prevents Doxorubicin-Induced DNA Damage and Apoptosis in Cardiomyocytes in Mice. Life Sci. 2020, 257, 118074. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, X.; Liu, Y.; Fang, K.; Liu, T. The Toxic Effects of Sulfoxaflor Induced in Earthworms (Eisenia Fetida) under Effective Concentrations. Int. J. Environ. Res. Public Heal. 2020, 17, 1740. [Google Scholar] [CrossRef] [Green Version]
- Lister, K.N.; Lamare, M.D.; Burritt, D.J. Maternal Antioxidant Provisioning Mitigates Pollutant-Induced Oxidative Damage in Embryos of the Temperate Sea Urchin Evechinus Chloroticus. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Costello, J. DNA Methylation: An Epigenetic Mark of Cellular Memory. Exp. Mol. Med. 2017, 49, e322. [Google Scholar] [CrossRef] [Green Version]
- Field, L.M.; Lyko, F.; Mandrioli, M.; Prantera, G. DNA Methylation in Insects. Insect. Mol. Biol. 2004, 13, 109–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyko, F.; Maleszka, R. Insects as Innovative Models for Functional Studies of DNA Methylation. Trends Genet. 2011, 27, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Horch, H.W.; Mito, T.; Popadi, A.; Ohuchi, H.; Noji, S. The Cricket as a Model Organism; Springer: Tokyo, Japan, 2017; ISBN 9784431564768. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effects | DNA Stability Parameters | |
|---|---|---|
| F | p | |
| Generation (G) | 21.664 | <0.0001 |
| Treatment (T) | 1.910 | 0.0791 |
| Strain (S) | 7.486 | 0.0002 |
| G × T | 3.307 | <0.0001 |
| T × S | 2.502 | 0.0225 |
| G × T × S | 6.129 | <0.0001 |
| Strain H | Parameter | Treatment | ||
|---|---|---|---|---|
| Control | Lower | Higher | ||
| (a) | AP sites | |||
| G1 | 2.61 | ↓ | ↑ | |
| G2 | 2.37 | ― | ― | |
| G3 | 3.26 | ↓ | ↓ | |
| G4 | 2.53 | ↓ | ↓ | |
| G5 | 3.33 | ↓ | ↓ | |
| G6 | 3.44 | ↑ | ↑ | |
| (b) | 8-OHdG | |||
| G1 | 30.88 | ↑ | ↓ | |
| G2 | 34.13 | ↓ | ↓ | |
| G3 | 27.96 | ― | ― | |
| G4 | 20.26 | ― | ― | |
| G5 | 23.38 | ↓ | ↓ | |
| G6 | 22.68 | ― | ― | |
| (c) | Global DNA methylation | |||
| G1 | 2.40 | ↓ | ― | |
| G2 | 1.21 | ↑ | ― | |
| G3 | 1.04 | ↑ | ↑ | |
| G4 | 1.76 | ↓ | ↓ | |
| G5 | 2.85 | ↓ | ↓ | |
| G6 | 0.46 | ↑ | ― | |
| Strain D | Parameter | Treatment | ||
|---|---|---|---|---|
| Control | Lower | Higher | ||
| (a) | AP sites | |||
| G1 | 2.82 | ↑ | ― | |
| G2 | 2.31 | ↑ | ― | |
| G3 | 2.50 | ↓ | ↓ | |
| G4 | 1.89 | ↑ | ↑ | |
| G5 | 3.74 | ↓ | ↓ | |
| G6 | 2.71 | ↑ | ↑ | |
| (b) | 8-OHdG | |||
| G1 | 28.50 | ↑ | ↑ | |
| G2 | 35.56 | ― | ↓ | |
| G3 | 31.68 | ↓ | ↓ | |
| G4 | 20.52 | ↑ | ↑ | |
| G5 | 20.14 | ― | ― | |
| G6 | 18.00 | ↑ | ↑ | |
| (c) | Global DNA methylation | |||
| G1 | 2.65 | ↓ | ↓ | |
| G2 | 1.10 | ↓ | ↑ | |
| G3 | 1.68 | ↓ | ↑ | |
| G4 | 1.61 | ↑ | ↑ | |
| G5 | 2.25 | ↓ | ↓ | |
| G6 | 0.76 | ↑ | ↑ | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flasz, B.; Ajay, A.K.; Tarnawska, M.; Babczyńska, A.; Majchrzycki, Ł.; Kędziorski, A.; Napora-Rutkowski, Ł.; Świerczek, E.; Augustyniak, M. Multigenerational Effects of Graphene Oxide Nanoparticles on Acheta domesticus DNA Stability. Int. J. Mol. Sci. 2023, 24, 12826. https://doi.org/10.3390/ijms241612826
Flasz B, Ajay AK, Tarnawska M, Babczyńska A, Majchrzycki Ł, Kędziorski A, Napora-Rutkowski Ł, Świerczek E, Augustyniak M. Multigenerational Effects of Graphene Oxide Nanoparticles on Acheta domesticus DNA Stability. International Journal of Molecular Sciences. 2023; 24(16):12826. https://doi.org/10.3390/ijms241612826
Chicago/Turabian StyleFlasz, Barbara, Amrendra K. Ajay, Monika Tarnawska, Agnieszka Babczyńska, Łukasz Majchrzycki, Andrzej Kędziorski, Łukasz Napora-Rutkowski, Ewa Świerczek, and Maria Augustyniak. 2023. "Multigenerational Effects of Graphene Oxide Nanoparticles on Acheta domesticus DNA Stability" International Journal of Molecular Sciences 24, no. 16: 12826. https://doi.org/10.3390/ijms241612826
APA StyleFlasz, B., Ajay, A. K., Tarnawska, M., Babczyńska, A., Majchrzycki, Ł., Kędziorski, A., Napora-Rutkowski, Ł., Świerczek, E., & Augustyniak, M. (2023). Multigenerational Effects of Graphene Oxide Nanoparticles on Acheta domesticus DNA Stability. International Journal of Molecular Sciences, 24(16), 12826. https://doi.org/10.3390/ijms241612826