
The Effect of Stimulants on Nectar Composition, Flowering, and Seed Yield of Common Buckwheat (Fagopyrum esculentum Moench)





Abstract
:1. Introduction
2. Results
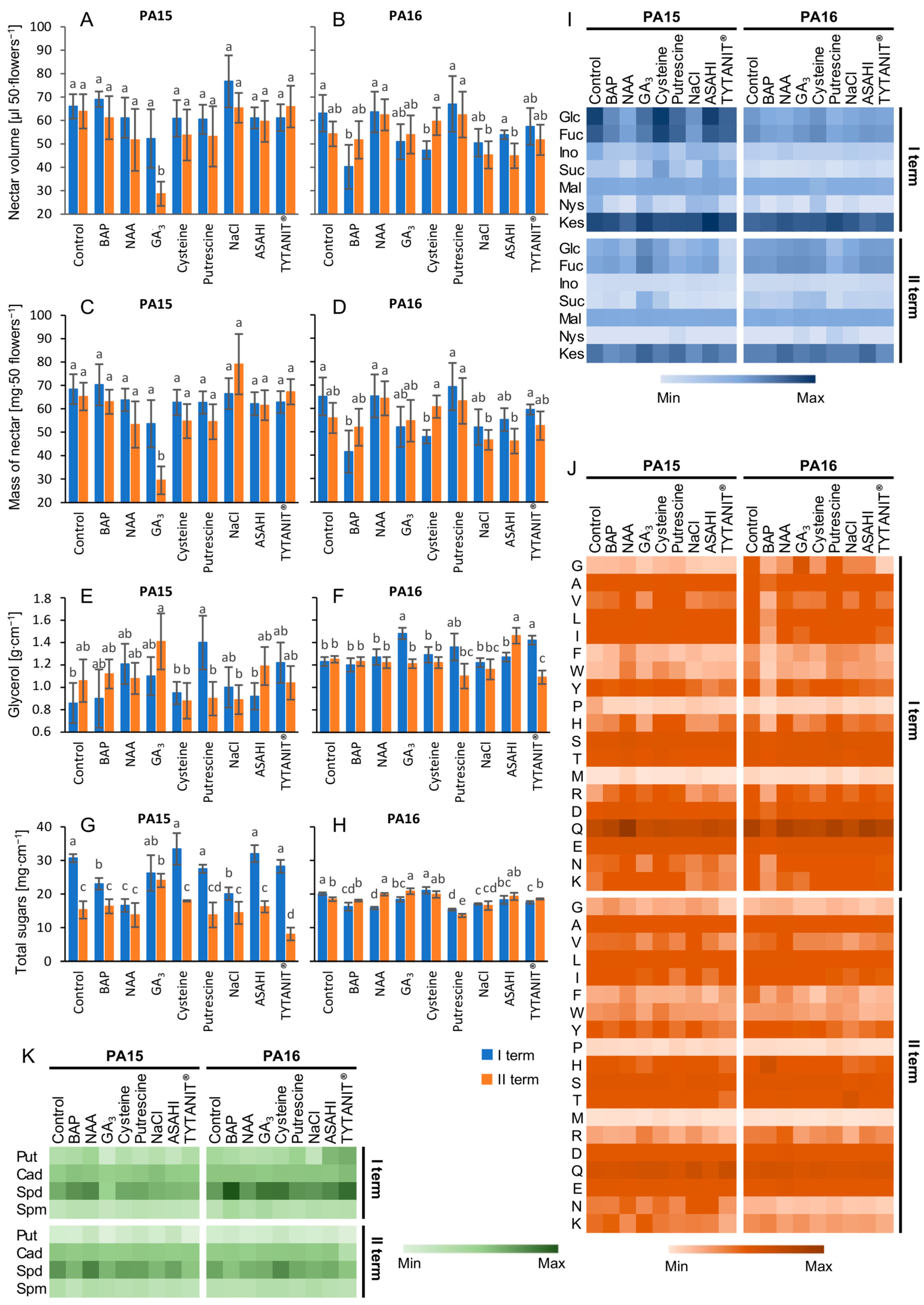
2.1. Mass, Volume, and Sugar Composition of the Nectar
2.2. Amino Acid Content
2.3. Polyamine Content
2.4. Flower Number and Seed Yield
3. Discussion
4. Materials and Methods
4.1. Plant Material and Cultivation
4.2. Stimulant Application
4.3. Analysis of Nectar Composition
4.3.1. Preparation of Buckwheat Flower Nectar Samples
4.3.2. Soluble Carbohydrates
4.3.3. Amino Acids
4.3.4. Polyamines
4.4. Flower and Seed Production
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Amino acids: | |
| A | Alanine |
| D | Aspartic acid |
| E | Glutamic acid |
| F | Phenylalanine |
| G | Glycine |
| H | Histidine |
| I | Isoleucine |
| K | Lysine |
| L | Leucine |
| M | Methionine |
| N | Asparagine |
| P | Proline |
| Q | Glutamine |
| R | Arginine |
| S | Serine |
| T | Threonine |
| W | Tryptophan |
| V | Valine |
| Y | Tyrosine |
| Other: | |
| BAP | 6-benzylaminopurine |
| Cad | Cadaverine |
| Fuc | Fructose |
| GA3 | Gibberellic acid |
| Glc | Glucose |
| Ino | Inositol |
| Kes | Kestose |
| Mal | Maltose |
| MTS | Mass of a thousand seeds |
| NAA | 1-Naphthaleneacetic acid |
| NaCl | Sodium chloride |
| Nys | Nystose |
| Put | Putrescine |
| Spd | Spermidine |
| Spm | Spermine |
References
- Verma, A.K. Influence of climate change on balanced ecosystem, biodiversity and sustainable development: An overview. Int. J. Biol. Innov. 2021, 3, 331–337. [Google Scholar] [CrossRef]
- Khairuddin, M.A.N.; Lasekan, O. Gluten-free cereal products and beverages: A review of their health benefits in the last five years. Foods 2021, 10, 2523. [Google Scholar] [CrossRef]
- Aubert, L.; Konrádová, D.; Kebbas, S.; Barris, S.; Quinet, M. Comparison of high temperature resistance in two buckwheat species Fagopyrum esculentum and Fagopyrum tataricum. J. Plant Physiol. 2020, 251, 153222. [Google Scholar] [CrossRef]
- Domingos, I.F.N.; Bilsborrow, P.E. The effect of variety and sowing date on the growth, development, yield and quality of common buckwheat (Fagopyrum esculentum Moench). Eur. J. Agron. 2021, 126, 126264. [Google Scholar] [CrossRef]
- Katar, D.; Olgun, M.; Turan, M. Analysis of morphological and biochemical characteristics of buckwheat (Fagopyrum esculentum Moench) in comparison with cereals. CyTA-J. Food 2016, 14, 176–185. [Google Scholar] [CrossRef]
- Nešović, M.; Gašić, U.; Tosti, T.; Horvacki, N.; Šikoparija, B.; Nedić, N.; Blagojević, S.; Ignjatović, L.; Tešić, Ž. Polyphenol profile of buckwheat honey, nectar and pollen: Polyphenolics in buckwheat. R. Soc. Open Sci. 2020, 7, 201576. [Google Scholar] [CrossRef]
- Vojtíšková, P.; Švec, P.; Kubán, V.; Krejzová, E.; Bittová, M.; Krácmar, S.; Svobodová, B. Chemical composition of buckwheat plant parts and selected buckwheat products. Potravinarstvo 2014, 8, 247–253. [Google Scholar] [CrossRef]
- Farkas, Á.; Zajácz, E.; Spring, E.; Pasture, B.E.E.; Summer, E. Nectar production for the Hungarian honey industry. Eur. J. Plant Sci. Biotechnol. 2007, 1, 125–151. [Google Scholar]
- Jacquemart, A.; Cawoy, V.; Kinet, J.M.; Ledent, J.F.; Quinet, M. Is buckwheat (Fagopyrum esculentum Moench) still a valuable crop today? Eur. J. Plant Sci. Biotechnol. 2012, 6, 1–10. [Google Scholar]
- FAOSTAT. FAO Statistics; Food and Agriculture Organization of the United Nations: Rome, Italy, 2022; Available online: http://faostat.fao.org/ (accessed on 9 April 2023).
- The Buckwheat Production in the World. Available online: https://knoema.com/data/agriculture-indicators-production+buckwheat (accessed on 21 July 2023).
- Płażek, A.; Słomka, A.; Kopeć, P.; Dziurka, M.; Hornyák, M.; Sychta, K.; Pastuszak, J.; Dubert, F. Effects of high temperature on embryological development and hormone profile in flowers and leaves of common buckwheat (Fagopyrum esculentum Moench). Int. J. Mol. Sci. 2019, 20, 1705. [Google Scholar] [CrossRef]
- Hornyák, M.; Płażek, A.; Kopeć, P.; Dziurka, M.; Pastuszak, J.; Szczerba, A.; Hura, T. Photosynthetic activity of common buckwheat (Fagopyrum esculentum Moench) exposed to thermal stress. Photosynthetica 2020, 58, 45–53. [Google Scholar] [CrossRef]
- Cawoy, V.; Kinet, J.M.; Jacquemart, A.L. Morphology of nectaries and biology of nectar production in the distylous species Fagopyrum esculentum. Ann. Bot. 2008, 102, 675–684. [Google Scholar] [CrossRef]
- Woo, S.-H.; Hena, A.; Kamal, M.; Suzuki, T.; Campbell, C.G.; Adachi, T.; Yun, Y.-H.; Chung, K.-Y.; Choi, J.-S. Buckwheat (Fagopyrum esculentum Moench.): Concepts, prospects and potential. Eur. J. Plant Sci. Biotechnol. 2009, 4, 1–16. [Google Scholar]
- Matsui, K.; Yasui, Y. Genetic and genomic research for the development of an efficient breeding system in heterostylous self-incompatible common buckwheat (Fagopyrum esculentum). Theor. Appl. Genet. 2020, 133, 1641–1653. [Google Scholar] [CrossRef]
- Baker, H.G. Floral nectar sugar constituents in relation to pollinator type. In Handbook of Experimental Pollination Biology; Jones, C.E., Little, R.J., Eds.; Van Nostrand Reynold Comp.: New York, NY, USA, 1983; pp. 117–141. [Google Scholar]
- Brzosko, E.; Bajguz, A.; Burzyńska, J.; Chmur, M. Does reproductive success in natural and anthropogenic populations of generalist Epipactis helleborine depend on flower morphology and nectar composition? Int. J. Mol. Sci. 2023, 24, 4276. [Google Scholar] [CrossRef]
- Alekseyeva, E.; Bureyko, L. Bee visitation, nectar productivity and pollen efficiency of common buckwheat. Fagopyrum 2000, 17, 77–80. [Google Scholar]
- Hornyák, M.; Słomka, A.; Sychta, K.; Dziurka, M.; Kopeć, P.; Pastuszak, J.; Szczerba, A.; Płażek, A. Reducing flower competition for assimilates by half results in higher yield of Fagopyrum esculentum. Int. J. Mol. Sci. 2020, 21, 8953. [Google Scholar] [CrossRef]
- Płażek, A.; Hura, K.; Hura, T.; Słomka, A.; Hornyák, M.; Sychta, K. Synthesis of heat-shock proteins HSP-70 and HSP-90 in flowers of common buckwheat (Fagopyrum esculentum) under thermal stress. Crop Pasture Sci. 2020, 71, 760. [Google Scholar] [CrossRef]
- Brown, P.; Saa, S. Biostimulants in agriculture. Front. Plant Sci. 2015, 6, 671. [Google Scholar] [CrossRef]
- Inoue, N.; Hagiwara, M. Analysis of the yielding process based on the module concept in common buckwheat. Fagopyrum 1999, 16, 73–77. [Google Scholar]
- Słomka, A.; Michno, K.; Dubert, F.; Dziurka, M.; Kopeć, P.; Płażek, A. Embryological background of low seed set in distylous common buckwheat (Fagopyrum esculentum Moench) with biased morph ratios, and stimulant- induced improvement of it. Crop Pasture Sci. 2017, 68, 680–690. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Stimulants in plant science: A global perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef]
- Ma, Y.; Freitas, H.; Dias, M.C. Strategies and prospects for biostimulants to alleviate abiotic stress in plants. Front. Plant Sci. 2022, 13, 1024243. [Google Scholar] [CrossRef]
- Bernier, G.; Havelange, A.; Houssa, C.; Petitjean, A.; Lejeune, P. Physiological signals that induce flowering. Plant Cell 1993, 5, 1147–1155. [Google Scholar] [CrossRef]
- Ahmad, P.; Kumar, A.; Gupta, A.; Hu, X.; Hakeem, K.R.; Azooz, M.M.; Sharma, S. Polyamines: Role in plants under abiotic stress. In Crop Production for Agricultural Improvement; Ashraf, M., Öztürk, M., Ahmad, M., Aksoy, A., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 491–512. [Google Scholar] [CrossRef]
- Shelp, B.J.; Bozzo, G.G.; Trobacher, C.P.; Zarei, A.; Deyman, K.L.; Brikis, C.J. Hypothesis/review: Contribution of putrescine to 4-aminobutyrate (GABA) production in response to abiotic stress. Plant Sci. 2012, 193, 130–135. [Google Scholar] [CrossRef]
- Zhao, B.; Liu, Q.; Wang, B.; Yuan, F. Roles of phytohormones and their signaling pathways in leaf development and stress responses. J. Agric. Food Chem. 2021, 69, 3566–3584. [Google Scholar] [CrossRef]
- Żur, I.; Klein, M.; Dubert, F.; Samek, L.; Waligórska, H.; Żuradzka, I.; Zawiślak, E. Environmental factors and genotypic variation of self-incompatibility in Brassica oleracea L. var capitata. Acta Biol. Cracov. Ser. Bot. 2003, 45, 49–52. [Google Scholar]
- Bianchi, M.B.; Gibbs, P.E. Bud pollination and other techniques are ineffective in breaking late-acting self-incompatibility in Ceiba chodatii (Malvaceae-Bombacoideae). Acta Bot. Brasil. 2021, 35, 381–388. [Google Scholar] [CrossRef]
- Przybysz, A.; Wrochna, M.; Słowiński, A.; Gawrońska, H. Stimulatory effect of Asahi SL on selected plant species. Acta Sci. Pol. Hortorum Cultus 2010, 9, 53–64. [Google Scholar]
- Gupta, S.; Doležal, K.; Kulkarni, M.G.; Balázs, E.; van Staden, J. Role of non-microbial stimulants in regulation of seed germination and seedling establishment. Plant Growth Regul. 2022, 97, 271–313. [Google Scholar] [CrossRef]
- Tian, Z.; Wang, J.W.; Li, J.; Han, B. Designing future crops: Challenges and strategies for sustainable agriculture. Plant J. 2021, 105, 1165–1178. [Google Scholar] [CrossRef]
- Nepi, M. Beyond nectar sweetness: The hidden ecological role of non-protein amino acids in nectar. J. Ecol. 2014, 102, 108–115. [Google Scholar] [CrossRef]
- Cawoy, V.; Lutts, S.; Kinet, J.M. Osmotic stress at seedling stage impairs reproductive development in buckwheat (Fagopyrum esculentum). Physiol. Plant. 2006, 128, 689–700. [Google Scholar] [CrossRef]
- Taylor, D.P.; Obendorf, R.L. Quantitative assessment of some factors limiting seed set in buckwheat. Crop Sci. 2001, 41, 1792–1799. [Google Scholar] [CrossRef]
- Halbrecq, B.; Romedenne, P.; Ledent, J.F. Evolution of flowering, ripening and seed set in buckwheat (Fagopyrum esculentum Moench): Quantitative analysis. Eur. J. Agron. 2005, 23, 209–224. [Google Scholar] [CrossRef]
- Farooq, S.; Rehman, R.U.; Pirzadah, T.B.; Malik, B.; Dar, F.A.; Tahir, I. Cultivation, agronomic practices, and growth performance of buckwheat. In Molecular Breeding and Nutritional Aspects of Buckwheat; Zhou, M., Kreft, I., Woo, S.-H., Chrungoo, N., Wieslander, G., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 299–319. [Google Scholar] [CrossRef]
- Petanidou, T.; Van Laere, A.; Ellis, W.N.; Smets, E. What shapes amino acid and sugar composition in Mediterranean floral nectars? Oikos 2006, 115, 155–169. [Google Scholar] [CrossRef]
- Singaravelan, N.; Man, G.N.E.E.; Inbar, M. Feeding responses of free-flying honeybees to secondary compounds mimicking floral nectars. J. Chem. Ecol. 2005, 31, 2791–2804. [Google Scholar] [CrossRef]
- Cheng, S. The Putrescine Puzzle: How do Manduca Sexta Hawkmoths Respond to Scented Nectar in Datura wrightii Flowers? Honors Thesis, Cornel University, Ithaca, NY, USA, 2022. Available online: https://ecommons.cornell.edu/bitstream/handle/1813/111444/ChengSheri_2022_Thesis.pdf?sequence=1& (accessed on 21 July 2023).
- Aloisi, I.; Distefano, G.; Antognoni, F.; Potente, G.; Parrotta, L.; Faleri, C.; Del Duca, S. Temperature-dependent compatible and incompatible pollen-style interactions in Citrus clementina Hort. ex Tan. Show different transglutaminase features and polyamine pattern. Front. Plant Sci. 2020, 11, 550424. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Univ. Calif. Agric. Exp. Station Circ. 1938, 347, 29–32. [Google Scholar]
- Hura, T.; Dziurka, M.; Hura, K.; Ostrowska, A.; Dziurka, K. Different allocation of carbohydrates and phenolics in dehydrated leaves of triticale. J. Plant Physiol. 2016, 202, 1–9. [Google Scholar] [CrossRef]
- Schuster, R. Determination of amino acids in biological, pharmaceutical, plant and food samples by automated precolumn derivatization and high-performance liquid chromatography. J. Chromatogr. B Biomed. Sci. Appl. 1988, 431, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Woodward, C.; Henderson, J.W.; Wielgos, T. High-Speed Amino Acid Analysis (AAA) on 1.8 µm Reversed-Phase (RP) Columns; Application note; Agilent Technologies, Inc.: Santa Clara, CA, USA, 2007; p. 5989-6297EN. [Google Scholar]
- Hura, T.; Dziurka, M.; Hura, K.; Ostrowska, A.; Dziurka, K. Free and cell wall-bound polyamines under long-term water stress applied at different growth stages of ×Triticosecale Wittm. PLoS ONE 2015, 10, e0135002. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Effects | Mass | Volume | Sugars | Amino Acids | Put | Cad | Spd | Spm |
|---|---|---|---|---|---|---|---|---|
| Line (L) | ns | *** | *** | ns | ** | ns | ** | *** |
| Stimulant (S) | * | ns | *** | ns | ns | *** | ns | ns |
| Term of application (T) | ns | ns | *** | *** | *** | *** | ** | ns |
| L × S | * | ns | ns | ns | ** | *** | ns | ns |
| L × T | ns | * | ** | ns | *** | *** | *** | ** |
| S × T | ns | ns | *** | ns | *** | *** | ns | ns |
| L × S × T | ns | ns | ns | ns | *** | *** | ns | ns |
| Effects | Flower Productivity | Seed Yield |
|---|---|---|
| Line (L) | *** | *** |
| Stimulant (S) | ns | *** |
| Term of application (T) | *** | ns |
| L × S | * | ns |
| L × T | ns | ns |
| S × T | ns | * |
| L × S × T | * | ns |
| Stimulant | PA15 | PA16 | ||||||
|---|---|---|---|---|---|---|---|---|
| No. of Flowers | No. of Empty Seeds | No. of Ripe Seeds | MTS | No. of Flowers | No. of Empty Seeds | No. of Ripe Seeds | MTS | |
| Term I | ||||||||
| Control | 616.1 c | 22.6 a | 117.9 c | 30.86 a | 699.3 a | 21.6 a | 145.7 a | 28.68 b |
| BAP | 632.7 c | 16.0 b | 133.4 b | 33.24 a | 444.4 c | 14.7 b | 134.1 b | 28.00 b |
| NAA | 715.3 b | 18.1 ab | 130.5 b | 30.30 a | 413.7 c | 14.9 b | 133.6 b | 29.97 b |
| GA3 | 624.5 c | 13.5 b | 130.0 b | 30.50 a | 539.7 b | 9.8 c | 137.5 b | 26.37 b |
| Cysteine | 876.0 a | 28.0 a | 160.0 a | 28.33 a | 490.2 bc | 11.9 b | 139.0 b | 28.27 b |
| NaCl | 630.0 c | 15.5 b | 132.0 b | 28.62 a | 461.0 c | 7.0 c | 149.9 a | 29.12 b |
| ASAHI SL | 653.7 c | 25.7 a | 168.9 a | 30.83 a | 398.4 cd | 13.3 b | 98.8 c | 38.22 a |
| TYTANIT® | 657.7 c | 25.5 a | 162.8 a | 31.84 a | 500.4 b | 11.0 b | 110.9 c | 27.13 b |
| Putrescine | 466.8 d | 23.4 a | 138.6 b | 32.71 a | 451.0 c | 16.0 b | 148.1 a | 26.53 b |
| Term II | ||||||||
| Control | 616.1 e | 22.6 b | 117.9 a | 30.86 a | 699.3 c | 21.6 a | 145.7 a | 28.68 b |
| BAP | 610.6 e | 23.9 bc | 112.1 a | 29.94 a | 880.6 a | 17.1 a | 101.3 c | 27.55 bc |
| NAA | 932.4 c | 27.2 b | 107.2 b | 31.54 a | 900.2 a | 19.7 a | 130.0 b | 26.44 c |
| GA3 | 1072.8 b | 26.1 b | 95.6 c | 31.83 a | 807.8 b | 13.7 b | 116.3 c | 27.70 bc |
| Cysteine | 935.8 c | 25.9 b | 104.4 b | 29.35 a | 564.4 d | 14.3 b | 84.2 d | 31.00 a |
| NaCl | 933.4 c | 28.1 b | 100.7 bc | 30.41 a | 833.8 b | 18.0 a | 108.9 c | 27.35 bc |
| ASAHI SL | 512.8 f | 18.2 c | 85.3 d | 34.08 a | 917.2 a | 14.8 b | 103.2 c | 28.72 b |
| TYTANIT® | 1268.8 a | 16.2 c | 90.6 cd | 28.17 a | 550.6 d | 14.2 b | 67.5 e | 29.17 ab |
| Putrescine | 910.6 c | 36.8 a | 99.7 c | 32.63 a | 822.7 b | 18.7 a | 107.4 c | 27.72 bc |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Płażek, A.; Dziurka, M.; Słomka, A.; Kopeć, P. The Effect of Stimulants on Nectar Composition, Flowering, and Seed Yield of Common Buckwheat (Fagopyrum esculentum Moench). Int. J. Mol. Sci. 2023, 24, 12852. https://doi.org/10.3390/ijms241612852
Płażek A, Dziurka M, Słomka A, Kopeć P. The Effect of Stimulants on Nectar Composition, Flowering, and Seed Yield of Common Buckwheat (Fagopyrum esculentum Moench). International Journal of Molecular Sciences. 2023; 24(16):12852. https://doi.org/10.3390/ijms241612852
Chicago/Turabian StylePłażek, Agnieszka, Michał Dziurka, Aneta Słomka, and Przemysław Kopeć. 2023. "The Effect of Stimulants on Nectar Composition, Flowering, and Seed Yield of Common Buckwheat (Fagopyrum esculentum Moench)" International Journal of Molecular Sciences 24, no. 16: 12852. https://doi.org/10.3390/ijms241612852