

Brassinosteroids Regulate the Water Deficit and Latex Yield of Rubber Trees

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
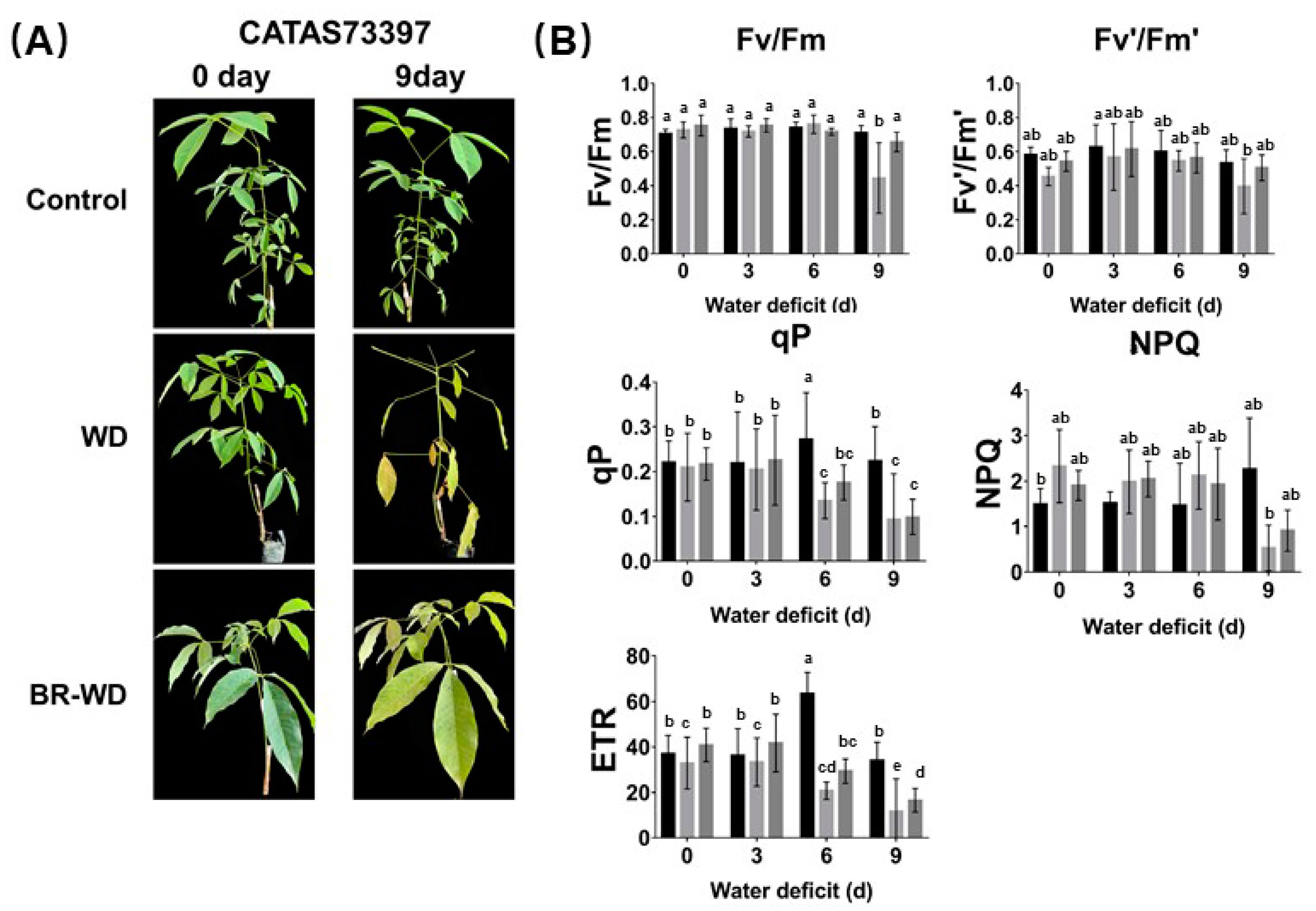
2.1. Effects of Exogenous BR on Photosynthetic Characteristics of Rubber Tree Leaves under Water Deficit
2.2. Effects of Exogenous BR on Physiological Characteristics of Rubber Tree Leaves under Water Deficit
2.3. Effect of BR on Latex Physiological Parameters of Rubber Trees
2.4. GO Enrichment and KEEG Pathway Enrichment Analysis of DEGs
2.5. Difference Analysis of Metabolite Accumulation
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatment
4.2. Determination the Physiological Parameters of Latex
4.3. Observation of Photosynthetic Fluorescence Parameters in the Leaves of Rubber Trees
4.4. Observation of Physiological Parameters of Rubber Trees
4.5. RNA Isolation, cDNA Synthesis, and qRT-PCR Analysis
4.6. RNA-Seq Analysis
4.7. Quantification of Phytohormones
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Planas-Riverola, A.; Gupta, A.; Betegón-Putze, I.; Bosch, N.; Ibañes, M.; Caño-Delgado, A.I. Brassinosteroid signaling in plant development and adaptation to stress. Development 2019, 146, dev151894. [Google Scholar] [CrossRef]
- Franck-Duchenne, M.; Wang, Y.; Ben Tahar, S.; Beachy, R.N. In vitro stem elongation of sweet pepper in media containing 24-epi-brassinolide. Plant Cell 1998, 53, 79–84. [Google Scholar]
- Li, Z.; He, Y. Roles of Brassinosteroids in Plant Reproduction. Int. J. Mol. Sci. 2020, 21, 872. [Google Scholar] [CrossRef]
- Kurepin, L.V.; Bey, M.A.; Back, T.G.; Pharis, R.P. Structure-Function Relationships of Four Stereoisomers of a Brassinolide Mimetic on Hypocotyl and Root Elongation of the Brassinosteroid-Deficient det2-1 Mutant of Arabidopsis. J. Plant Growth Regul. 2016, 35, 215–221. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Mohammadkhani, N. Enhancement of Chilling Stress Tolerance of Tomato Fruit by Postharvest Brassinolide Treatment. Food Bioprocess Technol. 2014, 7, 909–914. [Google Scholar] [CrossRef]
- Tanveer, M.; Shahzad, B.; Sharma, A.; Biju, S.; Bhardwaj, R. 24-Epibrassinolide; an active brassinolide and its role in salt stress tolerance in plants: A review. Plant Physiol. Biochem. 2018, 130, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Kumar, A.; Bhardwaj, R. Plant steroidal hormone epibrassinolide regulate—Heavy metal stress tolerance in Oryza sativa L. by modulating antioxidant defense expression. Environ. Exp. Bot. 2016, 122, 1–9. [Google Scholar] [CrossRef]
- Anjum, S.A.; Wang, L.; Farooq, M.A.; Hussain, M.; Xue, L.; Zou, C. Brassinolide Application Improves the Drought Tolerance in Maize Through Modulation of Enzymatic Antioxidants and Leaf Gas Exchange. J. Agron. Crop Sci. 2011, 197, 177–185. [Google Scholar] [CrossRef]
- Yuan, L.; Zhu, S.; Shu, S.; Sun, J.; Guo, S. Regulation of 2,4-epibrassinolide on mineral nutrient uptake and ion distribution in Ca(NO3)2 stressed cucumber plants. J. Plant physiol. 2015, 188, 29–36. [Google Scholar] [CrossRef]
- Zhao, G.W.; Xu, H.L.; Zhang, P.J.; Su, X.Y.; Zhao, H.J. Effects of 2,4-epibrassinolide on photosynthesis and Rubisco activase gene expression in Triticum aestivum L. seedlings under a combination of drought and heat stress. Plant Growth Regul. 2017, 81, 377–384. [Google Scholar] [CrossRef]
- Li, M.H.; Zhang, Y.F.; Xu, X.H.; Chen, Y.; Chu, J.Z.; Yao, X.Q. The combined treatments of brassinolide and zeaxanthin better alleviate oxidative damage and improve hypocotyl length, biomass, and the quality of radish sprouts stored at low temperature. Food Chem. X 2022, 15, 100394. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.; Ma, X.H.; Hussain, Q.; Asim, M.; Iqbal, A.; Ren, X.C.; Shah, S.H.; Chen, K.L.; Shi, Y. Application of 2,4-Epibrassinolide Improves Drought Tolerance in Tobacco through Physiological and Biochemical Mechanisms. Biology 2022, 11, 1192. [Google Scholar] [CrossRef]
- Xia, X.J.; Zhang, Y.; Wu, J.X.; Wang, J.T.; Zhou, Y.H.; Shi, K.; Yu, Y.L.; Yu, J.Q. Brassinosteroids Promote Metabolism of Pesticides in Cucumber. J. Agric. Food Chem. 2009, 57, 8406–8413. [Google Scholar] [CrossRef]
- Fariduddin, Q.; Yusuf, M.; Hayat, S.; Ahmad, A. Effect of 28-homobrassinolide on antioxidant capacity and photosynthesis in Brassica juncea plants exposed to different levels of copper. Environ. Exp. Bot. 2009, 66, 418–424. [Google Scholar] [CrossRef]
- Sahni, S.; Prasad, B.D.; Liu, Q.; Grbic, V.; Sharpe, A.; Singh, S.P.; Krishna, P. Overexpression of the brassinosteroid biosynthetic gene DWF4 in Brassica napus simultaneously increases seed yield and stress tolerance. Sci. Rep. 2016, 6, 28298. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.R.; Yu, D.Q. BRASSINOSTEROID INSENSITIVE2 Interacts with ABSCISIC ACID INSENSITIVE5 to Mediate the Antagonism of Brassinosteroids to Abscisic Acid during Seed Germination in Arabidopsis. Plant Cell 2014, 26, 4394–4408. [Google Scholar] [CrossRef] [PubMed]
- Goda, H.; Sawa, S.; Asami, T.; Fujioka, S.; Shimada, Y.; Yoshida, S. Comprehensive comparison of auxin-regulated and brassinosteroid-regulated genes in Arabidopsis. Plant Physiol. 2004, 134, 1555–1573. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.; Maharjan, P.M.; Lee, O.; Fujioka, S.; Jang, S.; Kim, B.; Takatsuto, S.; Tsujimoto, M.; Kim, H.; Cho, S.; et al. Auxin stimulates DWARF4 expression and brassinosteroid biosynthesis in Arabidopsis. Plant J. 2011, 66, 564–578. [Google Scholar] [CrossRef]
- Tong, H.; Xiao, Y.; Liu, D.; Gao, S.; Liu, L.; Yin, Y.; Jin, Y.; Qian, Q.; Chu, C. Brassinosteroid regulates cell elongation by modulating gibberellin metabolism in rice. Plant Cell 2014, 26, 4376–4393. [Google Scholar] [CrossRef]
- Unterholzner, S.J.; Rozhon, W.; Papacek, M.; Ciomas, J.; Lange, T.; Kugler, K.G.; Mayer, K.F.; Sieberer, T.; Poppenberger, B. Brassinosteroids Are Master Regulators of Gibberellin Biosynthesis in Arabidopsis. Plant Cell 2015, 27, 2261–2272. [Google Scholar] [CrossRef]
- Jiroutova, P.; Oklestkova, J.; Strnad, M. Crosstalk between Brassinosteroids and Ethylene during Plant Growth and under Abiotic Stress Conditions. Int. J. Mol. Sci. 2018, 19, 3283. [Google Scholar] [CrossRef] [PubMed]
- Muday, G.K.; Rahman, A.; Binder, B.M. Auxin and ethylene: Collaborators or competitors? Trends Plant Sci. 2012, 17, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Dai, L.; Yang, H.; Zhao, X.; Liu, M.; Wang, L. Comprehensive Analysis of BR Receptor Expression under Hormone Treatment in the Rubber Tree (Hevea brasiliensis Muell. Arg.). Plants 2023, 12, 1280. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Yang, H.; Dai, L.; Zhao, X.; Wang, L.F. Genome-wide identification and response stress expression analysis of the BES1 family in rubber tree (Hevea brasiliensis Muell. Arg.). PeerJ 2022, 10, e13189. [Google Scholar] [CrossRef] [PubMed]
- Guerra, N.B.; Sant’Ana Pegorin, G.; Boratto, M.H.; de Barros, N.R.; de Oliveira Graeff, C.F.; Herculano, R.D. Biomedical applications of natural rubber latex from the rubber tree Hevea brasiliensis. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 126, 112126. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, M.D.C.; Santoyo, G.; Glick, B.R. Recent Advances in the Bacterial Phytohormone Modulation of Plant Growth. Plants 2023, 12, 606. [Google Scholar] [CrossRef]
- Quintas-Nunes, F.; Brandao, P.R.; Barreto Crespo, M.T.; Glick, B.R.; Nascimento, F.X. Plant Growth Promotion, Phytohormone Production and Genomics of the Rhizosphere-Associated Microalga, Micractinium rhizosphaerae sp. nov. Plants 2023, 12, 651. [Google Scholar] [CrossRef]
- Li, X.; Liu, Y.; He, Q.; Li, S.; Liu, W.; Lin, C.; Miao, W. A Candidate Secreted Effector Protein of Rubber Tree Powdery Mildew Fungus Contributes to Infection by Regulating Plant ABA Biosynthesis. Front. Microbiol. 2020, 11, 591387. [Google Scholar] [CrossRef]
- Nakano, Y.; Mitsuda, N.; Ide, K.; Mori, T.; Mira, F.R.; Rosmalawati, S.; Watanabe, N.; Suzuki, K. Transcriptome analysis of Para rubber tree (H. brasiliensis) seedlings under ethylene stimulation. BMC Plant Biol. 2021, 21, 420. [Google Scholar] [CrossRef]
- Deng, X.; Guo, D.; Yang, S.; Shi, M.; Chao, J.; Li, H.; Peng, S.; Tian, W. Jasmonate signalling in the regulation of rubber biosynthesis in laticifer cells of rubber tree, Hevea brasiliensis. J. Exp. Bot. 2018, 69, 3559–3571. [Google Scholar] [CrossRef]
- Zhou, H.; Ashworth, K.; Dodd, I.C. Exogenous monoterpenes mitigate H2O2-induced lipid damage but do not attenuate photosynthetic decline during water deficit in tomato. J. Exp. Bot. 2023, erad219. [Google Scholar] [CrossRef]
- Zhang, H.H.; Xu, N.; Wu, X.Y.; Wang, J.R.; Ma, S.L.; Li, X.; Sun, G.Y. Effects of four types of sodium salt stress on plant growth and photosynthetic apparatus in sorghum leaves. J. Plant Interact. 2018, 13, 506–513. [Google Scholar] [CrossRef]
- Dias, M.C.; Bruggemann, W. Limitations of photosynthesis in Phaseolus vulgaris under drought stress: Gas exchange, chlorophyll fluorescence and Calvin cycle enzymes. Photosynthetica 2010, 48, 96–102. [Google Scholar] [CrossRef]
- Li, X.J.; Guo, X.; Zhou, Y.H.; Shi, K.; Zhou, J.; Yu, J.Q.; Xia, X.J. Overexpression of a brassinosteroid biosynthetic gene Dwarf enhances photosynthetic capacity through activation of Calvin cycle enzymes in tomato. BMC Plant Biol. 2016, 16, 33. [Google Scholar] [CrossRef] [PubMed]
- Ahammed, G.J.; Choudhary, S.P.; Chen, S.; Xia, X.; Shi, K.; Zhou, Y.; Yu, J. Role of brassinosteroids in alleviation of phenanthrene-cadmium co-contamination-induced photosynthetic inhibition and oxidative stress in tomato. J. Exp. Bot. 2013, 64, 199–213. [Google Scholar] [CrossRef] [PubMed]
- Verslues, P.E.; Agarwal, M.; Katiyar-Agarwal, S.; Zhu, J.; Zhu, J.K. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J. 2006, 45, 523–539. [Google Scholar] [CrossRef]
- Altuntas, C.; Demiralay, M.; Sezgin Muslu, A.; Terzi, R. Proline-stimulated signaling primarily targets the chlorophyll degradation pathway and photosynthesis associated processes to cope with short-term water deficit in maize. Photosynth. Res. 2020, 144, 35–48. [Google Scholar] [CrossRef]
- Jungklang, J.; Saengnil, K.; Uthaibutra, J. Effects of water-deficit stress and paclobutrazol on growth, relative water content, electrolyte leakage, proline content and some antioxidant changes in Curcuma alismatifolia Gagnep. cv. Chiang Mai Pink. Saudi J. Biol. Sci. 2017, 24, 1505–1512. [Google Scholar] [CrossRef]
- Yousfi, N.; Slama, I.; Ghnaya, T.; Sayoure, A.; Abdelly, C. Effects of water deficit stress on growth, water relations and osmolyte accumulation in Medicago truncatula and M. laciniata populations. C. R. Biol. 2010, 333, 205–213. [Google Scholar] [CrossRef]
- Gao, X.; Ma, J.; Tie, J.; Li, Y.; Hu, L.; Yu, J. BR-Mediated Protein S-Nitrosylation Alleviated Low-Temperature Stress in Mini Chinese Cabbage (Brassica rapa ssp. pekinensis). Int. J. Mol. Sci. 2022, 23, 10964. [Google Scholar] [CrossRef]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef]
- Xia, X.J.; Chen, Z.; Yu, J.Q. ROS mediate brassinosteroids-induced plant stress responses. Plant Signal. Behav. 2010, 5, 532–534. [Google Scholar] [CrossRef]
- Chaudhuri, A.; Halder, K.; Abdin, M.Z.; Majee, M.; Datta, A. Abiotic Stress Tolerance in Plants: Brassinosteroids Navigate Competently. Int. J. Mol. Sci. 2022, 23, 14577. [Google Scholar] [CrossRef]
- Gao, L.; Sun, Y.; Wu, M.; Wang, D.; Wei, J.; Wu, B.; Wang, G.; Wu, W.; Jin, X.; Wang, X.; et al. Physiological and Proteomic Analyses of Molybdenum- and Ethylene-Responsive Mechanisms in Rubber Latex. Front. Plant Sci. 2018, 9, 621. [Google Scholar] [CrossRef]
- Pakianathan, S.W.; Samsidar, H.; Sivakumaran, S.; Gomez, J.B. Physiological and anatomical investigation on long term ethephon-stimulated tress. J. Rubber Res. 1982, 30, 63–79. [Google Scholar]
- Wititsuwannakul, R.; Pasitkul, P.; Kanokwiroon, K.; Wititsuwannakul, D. A role for a Hevea latex lectin-like protein in mediating rubber particle aggregation and latex coagulation. Phytochemistry 2008, 69, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Gidrol, X.; Chrestin, H.; Tan, H.L.; Kush, A. Hevein, a lectin-like protein from Hevea brasiliensis (rubber tree) is involved in the coagulation of latex. J. Biol. Chem. 1994, 269, 9278–9283. [Google Scholar] [CrossRef]
- Kim, W.S.; Eum, C.H.; Molnar, A.; Yu, J.S.; Lee, S. Repeatability and reproducibility of thermal field-flow fractionation in molecular weight determination of processed natural rubber. Analyst 2006, 131, 429–433. [Google Scholar] [CrossRef] [PubMed]
- Santner, A.; Estelle, M. Recent advances and emerging trends in plant hormone signalling. Nature 2009, 459, 1071–1078. [Google Scholar] [CrossRef]
- Yang, J.; Duan, G.H.; Li, C.Q.; Liu, L.; Han, G.Y.; Zhang, Y.L.; Wang, C.M. The Crosstalks between Jasmonic Acid and Other Plant Hormone Signaling Highlight the Involvement of Jasmonic Acid as a Core Component in Plant Response to Biotic and Abiotic Stresses. Front. Plant Sci. 2019, 10, 1349. [Google Scholar] [CrossRef]
- Santner, A.; Calderon-Villalobos, L.I.A.; Estelle, M. Plant hormones are versatile chemical regulators of plant growth. Nat. Chem. Biol. 2009, 5, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Zander, M.; Huang, S.C.; Xie, M.; Song, L.; Guzm, N.J.; Hann, E.; Shanbhag, B.K.; Ng, S.; Jain, S.; et al. Transcription Factor Dynamics in Cross-Regulation of Plant Hormone Signaling Pathways. bioRxiv 2023. [Google Scholar] [CrossRef]
- Li, Q.; Xu, F.; Chen, Z.; Teng, Z.; Sun, K.; Li, X.; Yu, J.; Zhang, G.; Liang, Y.; Huang, X.; et al. Synergistic interplay of ABA and BR signal in regulating plant growth and adaptation. Nat. Plants 2021, 7, 1108–1118. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Liu, Q.C.; Mou, Z.L. A direct link between BR and SA signaling: Negative regulation of TGA4 by BIN2. Mol. Plant 2022, 15, 1254–1256. [Google Scholar] [CrossRef] [PubMed]
- Zu, S.H.; Jiang, Y.T.; Chang, J.H.; Zhang, Y.J.; Xue, H.W.; Lin, W.H. Interaction of brassinosteroid and cytokinin promotes ovule initiation and increases seed number per silique in Arabidopsis. J. Integr. Plant Biol. 2022, 64, 702–716. [Google Scholar] [CrossRef]
- Hu, J.L.; Huang, J.; Xu, H.S.; Wang, Y.S.; Li, C.; Wen, P.Z.; You, X.M.; Zhang, X.; Pan, G.; Li, Q.; et al. Rice stripe virus suppresses jasmonic acid-mediated resistance by hijacking brassinosteroid signaling pathway in rice. PLoS Pathog. 2020, 16, e1008801. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Deng, X.G.; Zhou, X.; Zhu, L.S.; Zou, L.J.; Li, P.X.; Zhang, D.W.; Lin, H.H. Ethylene and hydrogen peroxide are involved in brassinosteroid-induced salt tolerance in tomato. Sci. Rep. 2016, 6, 35392. [Google Scholar] [CrossRef]
- Li, Q.F.; Zhou, Y.; Xiong, M.; Ren, X.Y.; Han, L.; Wang, J.D.; Zhang, C.Q.; Fan, X.L.; Liu, Q.Q. Gibberellin recovers seed germination in rice with impaired brassinosteroid signalling. Plant Sci. 2020, 293, 110435. [Google Scholar] [CrossRef]
- Depuydt, S.; Hardtke, C.S. Hormone Signalling Crosstalk in Plant Growth Regulation. Curr. Biol. 2011, 21, R365–R373. [Google Scholar] [CrossRef]
- Jaillais, Y.; Chory, J. Unraveling the paradoxes of plant hormone signaling integration. Nat. Struct. Mol. Biol. 2010, 17, 642–645. [Google Scholar] [CrossRef]
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R.; et al. Arabidopsis transcription factors: Genome-wide comparative analysis among eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar] [CrossRef] [PubMed]
- Hao, B.W.; Wu, J.L. Laticifer Differentiation in Hevea brasiliensis: Induction by Exogenous Jasmonic Acid and Linolenic Acid. Ann. Bot. 2000, 85, 37–43. [Google Scholar] [CrossRef]
- Junaidi Nuringtyas, T.R.; Clément-Vidal, A.; Flori, A.; Syafaah, A.; Oktavia, F.; Ismawanto, S.; Aji, M.; Subandiyah, S.; Montoro, P. Analysis of reduced and oxidized antioxidants in Hevea brasiliensis latex reveals new insights into the regulation of antioxidants in response to harvesting stress and tapping panel dryness. Heliyon 2022, 8, e09840. [Google Scholar] [CrossRef]
- Herlinawati, E.; Montoro, P.; Ismawanto, S.; Syafaah, A.; Aji, M.; Giner, M.; Flori, A.; Gohet, E.; Oktavia, F. Dynamic analysis of Tapping Panel Dryness in Hevea brasiliensis reveals new insights on this physiological syndrome affecting latex production. Heliyon 2022, 8, e10920. [Google Scholar] [CrossRef]
- Wang, L.F. Physiological and molecular responses to variation of light intensity in rubber Tree (Hevea brasiliensis Muell. Arg.). PLoS ONE 2014, 9, e89514. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, B.; Liu, M.; Yang, H.; Dai, L.; Wang, L. Brassinosteroids Regulate the Water Deficit and Latex Yield of Rubber Trees. Int. J. Mol. Sci. 2023, 24, 12857. https://doi.org/10.3390/ijms241612857
Guo B, Liu M, Yang H, Dai L, Wang L. Brassinosteroids Regulate the Water Deficit and Latex Yield of Rubber Trees. International Journal of Molecular Sciences. 2023; 24(16):12857. https://doi.org/10.3390/ijms241612857
Chicago/Turabian StyleGuo, Bingbing, Mingyang Liu, Hong Yang, Longjun Dai, and Lifeng Wang. 2023. "Brassinosteroids Regulate the Water Deficit and Latex Yield of Rubber Trees" International Journal of Molecular Sciences 24, no. 16: 12857. https://doi.org/10.3390/ijms241612857
APA StyleGuo, B., Liu, M., Yang, H., Dai, L., & Wang, L. (2023). Brassinosteroids Regulate the Water Deficit and Latex Yield of Rubber Trees. International Journal of Molecular Sciences, 24(16), 12857. https://doi.org/10.3390/ijms241612857