
Targeting SARS-CoV-2 Non-Structural Proteins



Abstract
:1. Introduction
2. Description of the NSPs Present in the SARS-CoV-2 Genome
2.1. NSP1
2.2. NSP2
2.3. NSP3
2.4. NSP4
2.5. NSP5
2.6. NSP6
2.7. NSP7
2.8. NSP8
2.9. NSP9
2.10. NSP10
2.11. NSP11
2.12. NSP12
2.13. NSP13
2.14. NSP14
2.15. NSP15
2.16. NSP16
3. Targeting NSPs with Small Molecule Inhibitors
3.1. NSP1
3.2. NSP3
3.3. NSP5
3.4. NSP6
3.5. NSP9
3.6. NSP10
3.7. NSP13
3.8. NSP14
3.9. NSP15
4. Approved Treatments
4.1. Remdesivir
4.2. Paxlovid
4.3. Molnupiravir
4.4. Casirivimab/Imdevinmab
4.5. Tocilizumab/Sarilumab
4.6. Baricitinib
5. Similarities between Approved Treatments and Natural Products
6. Treatment Management
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bchetnia, M.; Girard, C.; Duchaine, C.; Laprise, C. The outbreak of the novel severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2): A review of the current global status. J. Infect. Public Health 2020, 13, 1601–1610. [Google Scholar] [CrossRef]
- Brant, A.C.; Tian, W.; Majerciak, V.; Yang, W.; Zheng, Z.-M. SARS-CoV-2: From its discovery to genome structure, transcription, and replication. Cell Biosci. 2021, 11, 136. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.; Chaudhary, J.K.; Jain, N.; Chaudhary, P.K.; Khanra, S.; Dhamija, P.; Sharma, A.; Kumar, A.; Handu, S. Role of structural and non-structural proteins and therapeutic targets of SARS-CoV-2 for COVID-19. Cells 2021, 10, 821. [Google Scholar] [CrossRef]
- Yan, R.; Zhang, Y.; Li, Y.; Ye, F.; Guo, Y.; Xia, L.; Zhong, X.; Chi, X.; Zhou, Q. Structural basis for the different states of the spike protein of SARS-CoV-2 in complex with ACE2. Cell Res. 2021, 31, 717–719. [Google Scholar] [CrossRef]
- Low, Z.Y.; Zabidi, N.Z.; Yip, A.J.W.; Puniyamurti, A.; Chow, V.T.K.; Lal, S.K. SARS-CoV-2 non-structural proteins and their roles in host immune evasion. Viruses 2022, 14, 1991. [Google Scholar] [CrossRef] [PubMed]
- Redondo, N.; Zaldívar-López, S.; Garrido, J.J.; Montoya, M. SARS-CoV-2 Accessory Proteins in Viral Pathogenesis: Knowns and Unknowns. Front Immunol. 2021, 12, 708264. [Google Scholar] [CrossRef]
- Chandel, V.; Sharma, P.P.; Raj, S.; Choudhari, R.; Rathi, B.; Kumar, D. Structure-based drug repurposing for targeting Nsp9 replicase and spike proteins of severe acute respiratory syndrome coronavirus 2. J. Biomol. Struct. Dyn. 2022, 40, 249–262. [Google Scholar] [CrossRef]
- Raj, R. Analysis of non-structural proteins, NSPs of SARS-CoV-2 as targets for computational drug designing. Biochem. Biophys. Rep. 2021, 25, 100847. [Google Scholar] [CrossRef]
- Wang, B.; Svetlov, D.; Artsimovitch, I. NMPylation and de-NMPylation of SARS-CoV-2 nsp9 by the NiRAN domain. Nucleic Acids Res. 2021, 49, 8822–8835. [Google Scholar] [CrossRef]
- Yapasert, R.; Khaw-On, P.; Banjerdpongchai, R. Coronavirus infection-associated cell death signaling and potential therapeutic targets. Molecules 2021, 26, 7459. [Google Scholar] [CrossRef] [PubMed]
- Wolff, G.; Melia, C.E.; Snijder, E.J.; Bárcena, M. Double-Membrane Vesicles as Platforms for Viral Replication. Trends Microbiol. 2020, 28, 1022–1033. [Google Scholar] [CrossRef]
- Klein, S.; Cortese, M.; Winter, S.L.; Wachsmuth-Melm, M.; Neufeldt, C.J.; Cerikan, B.; Stanifer, M.L.; Boulant, S.; Bartenschlager, R.; Chlanda, P. SARS-CoV-2 structure and replication characterized by in situ cryo-electron tomography. Nat. Commun. 2020, 11, 5885. [Google Scholar] [CrossRef] [PubMed]
- Ahmed-Belkacem, R.; Sutto-Ortiz, P.; Guiraud, M.; Canard, B.; Vasseur, J.-J.; Decroly, E.; Debart, F. Synthesis of adenine dinucleosides SAM analogs as specific inhibitors of SARS-CoV nsp14 RNA cap guanine-N7-methyltransferase. Eur. J. Med. Chem. 2020, 201, 112557. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.; Mukherjee, R.; Grewe, D.; Bojkova, D.; Baek, K.; Bhattacharya, A.; Schulz, L.; Widera, M.; Mehdipour, A.R.; Tascher, G.; et al. Papain-like protease regulates SARS-CoV-2 viral spread and innate immunity. Nature 2020, 587, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Tazikeh-Lemeski, E.; Moradi, S.; Raoufi, R.; Shahlaei, M.; Janlou, M.A.M.; Zolghadri, S. Targeting SARS-CoV-2 non-structural protein 16: A virtual drug repurposing study. J. Biomol. Struct. Dyn. 2021, 39, 4633–4646. [Google Scholar] [CrossRef] [PubMed]
- Santerre, M.; Arjona, S.P.; Allen, C.N.; Shcherbik, N.; Sawaya, B.E. Why do SARS-CoV-2 NSPs rush to the ER? J. Neurol. 2021, 268, 2013–2022. [Google Scholar] [CrossRef]
- Parthasarathy, H.; Tandel, D.; Siddiqui, A.H.; Harshan, K.H. Metformin suppresses SARS-CoV-2 in cell culture. Virus Res. 2022, 323, 199010. [Google Scholar] [CrossRef]
- Pitsillou, E.; Liang, J.; Ververis, K.; Lim, K.W.; Hung, A.; Karagiannis, T.C. Identification of small molecule inhibitors of the deubiquitinating activity of the SARS-CoV-2 papain-like protease: In silico molecular docking studies and in vitro enzymatic activity assay. Front. Chem. 2020, 8, 623971. [Google Scholar] [CrossRef]
- Rao, P.; Patel, R.; Shukla, A.; Parmar, P.; Rawal, R.M.; Saraf, M.; Goswami, D. Identifying structural-functional analogue of GRL0617, the only well-established inhibitor for papain-like protease (PLpro) of SARS-CoV2 from the pool of fungal metabolites using docking and molecular dynamics simulation. Mol. Divers. 2022, 26, 309–329. [Google Scholar] [CrossRef]
- Xian, H.; Liu, Y.; Rundberg Nilsson, A.; Gatchalian, R.; Crother, T.R.; Tourtellotte, W.G.; Zhang, Y.; Aleman-Muench, G.R.; Lewis, G.; Chen, W.; et al. Metformin inhibition of mitochondrial ATP and DNA synthesis abrogates NLRP3 inflammasome activation and pulmonary inflammation. Immunity 2021, 54, 1463–1477.e11. [Google Scholar] [CrossRef]
- Afsar, M.; Narayan, R.; Akhtar, M.N.; Das, D.; Rahil, H.; Nagaraj, S.K.; Eswarappa, S.M.; Tripathi, S.; Hussain, T. Drug targeting Nsp1-ribosomal complex shows antiviral activity against SARS-CoV-2. eLife 2022, 11, e74877. [Google Scholar] [CrossRef] [PubMed]
- Makiyama, K.; Hazawa, M.; Kobayashi, A.; Lim, K.; Voon, D.C.; Wong, R.W. NSP9 of SARS-CoV-2 attenuates nuclear transport by hampering nucleoporin 62 dynamics and functions in host cells. Biochem. Biophys. Res Commun. 2022, 586, 137–142. [Google Scholar] [CrossRef]
- Lapointe, C.P.; Grosely, R.; Johnson, A.G.; Wang, J.; Fernández, I.S.; Puglisi, J.D. Dynamic competition between SARS-CoV-2 NSP1 and mRNA on the human ribosome inhibits translation initiation. Proc. Natl. Acad. Sci. USA 2021, 118, e2017715118. [Google Scholar] [CrossRef] [PubMed]
- Cornillez-Ty, C.T.; Liao, L.; Yates, J.R.; Kuhn, P.; Buchmeier, M.J. Severe Acute Respiratory Syndrome Coronavirus Nonstructural Protein 2 Interacts with a Host Protein Complex Involved in Mitochondrial Biogenesis and Intracellular Signaling. J. Virol. 2009, 83, 10314–10318. [Google Scholar] [CrossRef]
- Angelini, M.M.; Akhlaghpour, M.; Neuman, B.W.; Buchmeier, M.J. Severe Acute Respiratory Syndrome Coronavirus Nonstructural Proteins 3, 4, and 6 Induce Double-Membrane Vesicles. mBio 2013, 4, e00524-13. [Google Scholar] [CrossRef] [PubMed]
- Roe, M.K.; Junod, N.A.; Young, A.R.; Beachboard, D.C.; Stobart, C.C. Targeting novel structural and functional features of coronavirus protease nsp5 (3CLpro, Mpro) in the age of COVID-19. J. Gen. Virol. 2021, 102, 001558. [Google Scholar] [CrossRef]
- Kirchdoerfer, R.N.; Ward, A.B. Structure of the SARS-CoV nsp12 polymerase bound to nsp7 and nsp8 co-factors. Nat. Commun. 2019, 10, 2342. [Google Scholar] [CrossRef]
- Sutton, G.; Fry, E.; Carter, L.; Sainsbury, S.; Walter, T.; Nettleship, J.; Berrow, N.; Owens, R.; Gilbert, R.; Davidson, A.; et al. The nsp9 Replicase Protein of SARS-Coronavirus, Structure and Functional Insights. Structure 2004, 12, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Bouvet, M.; Imbert, I.; Subissi, L.; Gluais, L.; Canard, B.; Decroly, E. RNA 3′-end mismatch excision by the severe acute respiratory syndrome coronavirus nonstructural protein nsp10/nsp14 exoribonuclease complex. Proc. Natl. Acad. Sci. USA 2012, 109, 9372–9377. [Google Scholar] [CrossRef] [PubMed]
- Gadhave, K.; Kumar, P.; Kumar, A.; Bhardwaj, T.; Garg, N.; Giri, R. Conformational dynamics of 13 amino acids long NSP11 of SARS-CoV-2 under membrane mimetics and different solvent conditions. Microb. Pathog. 2021, 158, 105041. [Google Scholar] [CrossRef]
- Suryawanshi, R.K.; Koganti, R.; Agelidis, A.; Patil, C.D.; Shukla, D. Dysregulation of Cell Signaling by SARS-CoV-2. Trends Microbiol. 2021, 29, 224–237. [Google Scholar] [CrossRef]
- Newman, J.A.; Douangamath, A.; Yadzani, S.; Yosaatmadja, Y.; Aimon, A.; Brandão-Neto, J.; Dunnett, L.; Gorrie-stone, T.; Skyner, R.; Fearon, D.; et al. Structure, mechanism and crystallographic fragment screening of the SARS-CoV-2 NSP13 helicase. Nat. Commun. 2021, 12, 4848. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.C.-C.; Laurent-Rolle, M.; Pawlak, J.B.; Wilen, C.B.; Cresswell, P. Translational shutdown and evasion of the innate immune response by SARS-CoV-2 NSP14 protein. Proc. Natl. Acad. Sci. USA 2021, 118, e2101161118. [Google Scholar] [CrossRef]
- Kim, Y.; Jedrzejczak, R.; Maltseva, N.I.; Wilamowski, M.; Endres, M.; Godzik, A.; Michalska, K.; Joachimiak, A. Crystal structure of Nsp15 endoribonuclease NendoU from SARS-CoV-2. Protein Sci. 2020, 29, 1596–1605. [Google Scholar] [CrossRef]
- Sk, M.F.; Jonniya, N.A.; Roy, R.; Poddar, S.; Kar, P. Computational Investigation of Structural Dynamics of SARS-CoV-2 Methyltransferase-Stimulatory Factor Heterodimer nsp16/nsp10 Bound to the Cofactor SAM. Front. Mol. Biosci. 2020, 7, 590165. [Google Scholar] [CrossRef]
- Thoms, M.; Buschauer, R.; Ameismeier, M.; Koepke, L.; Denk, T.; Hirschenberger, M.; Kratzat, H.; Hayn, M.; Mackens-Kiani, T.; Cheng, J.; et al. Structural basis for translational shutdown and immune evasion by the Nsp1 protein of SARS-CoV-2. Science 2020, 369, 1249–1255. [Google Scholar] [CrossRef] [PubMed]
- Schubert, K.; Karousis, E.D.; Jomaa, A.; Scaiola, A.; Echeverria, B.; Gurzeler, L.-A.; Leibundgut, M.; Thiel, V.; Mühlemann, O.; Ban, N. SARS-CoV-2 Nsp1 binds the ribosomal mRNA channel to inhibit translation. Nat. Struct. Mol. Biol. 2020, 27, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Kao, H.-T.; Orry, A.; Palfreyman, M.G.; Porton, B. Synergistic interactions of repurposed drugs that inhibit Nsp1, a major virulence factor for COVID-19. Sci. Rep. 2022, 12, 10174. [Google Scholar] [CrossRef]
- Vora, S.M.; Fontana, P.; Mao, T.; Leger, V.; Zhang, Y.; Fu, T.-M.; Lieberman, J.; Gehrke, L.; Shi, M.; Wang, L.; et al. Targeting stem-loop 1 of the SARS-CoV-2 5′ UTR to suppress viral translation and Nsp1 evasion. Proc. Natl. Acad. Sci USA 2022, 119, e2117198119. [Google Scholar] [CrossRef] [PubMed]
- Gaglia, M.M.; Covarrubias, S.; Wong, W.; Glaunsinger, B.A. A Common Strategy for Host RNA Degradation by Divergent Viruses. J. Virol. 2012, 86, 9527–9530. [Google Scholar] [CrossRef]
- Zhang, K.; Miorin, L.; Makio, T.; Dehghan, I.; Gao, S.; Xie, Y.; Zhong, H.; Esparza, M.; Kehrer, T.; Kumar, A.; et al. Nsp1 protein of SARS-CoV-2 disrupts the mRNA export machinery to inhibit host gene expression. Sci. Adv. 2021, 7, eabe7386. [Google Scholar] [CrossRef] [PubMed]
- Tatsuta, T.; Model, K.; Langer, T. Formation of membrane-bound ring complexes by prohibitins in mitochondria. Mol. Biol. Cell. 2005, 16, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Gamble, S.C.; Chotai, D.; Odontiadis, M.; Dart, D.A.; Brooke, G.N.; Powell, S.M.; Reebye, V.; Varela-Carver, A.; Kawano, Y.; Waxman, J.; et al. Prohibitin, a protein downregulated by androgens, represses androgen receptor activity. Oncogene 2007, 26, 1757–1768. [Google Scholar] [CrossRef]
- Wintachai, P.; Wikan, N.; Kuadkitkan, A.; Jaimipuk, T.; Ubol, S.; Pulmanausahakul, R.; Auewarakul, P.; Kasinrerk, W.; Weng, W.-Y.; Panyasrivanit, M.; et al. Identification of prohibitin as a Chikungunya virus receptor protein. J. Med. Virol. 2012, 84, 1757–1770. [Google Scholar] [CrossRef]
- Kuadkitkan, A.; Wikan, N.; Fongsaran, C.; Smith, D.R. Identification and characterization of prohibitin as a receptor protein mediating DENV-2 entry into insect cells. Virology 2010, 406, 149–161. [Google Scholar] [CrossRef]
- Milligan, J.C.; Zeisner, T.U.; Papageorgiou, G.; Joshi, D.; Soudy, C.; Ulferts, R.; Wu, M.; Lim, C.T.; Tan, K.W.; Weissmann, F.; et al. Identifying SARS-CoV-2 antiviral compounds by screening for small molecule inhibitors of Nsp5 main protease. Biochem. J. 2021, 478, 2499–2515. [Google Scholar] [CrossRef]
- Cottam, E.M.; Maier, H.J.; Manifava, M.; Vaux, L.C.; Chandra-Schoenfelder, P.; Gerner, W.; Britton, P.; Ktistakis, N.T.; Wileman, T. Coronavirus nsp6 proteins generate autophagosomes from the endoplasmic reticulum via an omegasome intermediate. Autophagy 2011, 7, 1335–1347. [Google Scholar] [CrossRef] [PubMed]
- Cottam, E.M.; Whelband, M.C.; Wileman, T. Coronavirus NSP6 restricts autophagosome expansion. Autophagy 2014, 10, 1426–1441. [Google Scholar] [CrossRef]
- Sun, X.; Liu, Y.; Huang, Z.; Xu, W.; Hu, W.; Yi, L.; Liu, Z.; Chan, H.; Zeng, J.; Liu, X.; et al. SARS-CoV-2 non-structural protein 6 triggers NLRP3-dependent pyroptosis by targeting ATP6AP1. Cell Death Differ. 2022, 29, 1240–1254. [Google Scholar] [CrossRef] [PubMed]
- Te Velthuis, A.J.W.; van den Worm, S.H.E.; Snijder, E.J. The SARS-coronavirus nsp7+nsp8 complex is a unique multimeric RNA polymerase capable of both de novo initiation and primer extension. Nucleic Acids Res. 2012, 40, 1737–1747. [Google Scholar] [CrossRef]
- Xia, H.; Cao, Z.; Xie, X.; Zhang, X.; Chen, J.Y.-C.; Wang, H.; Menachery, V.D.; Rajsbaum, R.; Shi, P.-Y. Evasion of Type I Interferon by SARS-CoV-2. Cell Rep. 2020, 33, 108234. [Google Scholar] [CrossRef]
- Decroly, E.; Debarnot, C.; Ferron, F.; Bouvet, M.; Coutard, B.; Imbert, I.; Gluais, L.; Papageorgiou, N.; Sharff, A.; Bricogne, G.; et al. Crystal Structure and Functional Analysis of the SARS-Coronavirus RNA Cap 2′-O-Methyltransferase nsp10/nsp16 Complex. PLoS Pathog. 2011, 7, e1002059. [Google Scholar] [CrossRef]
- Ma, Y.; Wu, L.; Shaw, N.; Gao, Y.; Wang, J.; Sun, Y.; Lou, Z.; Yan, L.; Zhang, R.; Rao, Z. Structural basis and functional analysis of the SARS coronavirus nsp14–nsp10 complex. Proc. Natl. Acad. Sci. USA 2015, 112, 9436–9441. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Peng, R.; Yuan, B.; Zhao, J.; Wang, M.; Wang, X.; Wang, Q.; Sun, Y.; Fan, Z.; Qi, J.; et al. Structural and Biochemical Characterization of the nsp12-nsp7-nsp8 Core Polymerase Complex from SARS-CoV-2. Cell Rep. 2020, 31, 107774. [Google Scholar] [CrossRef] [PubMed]
- Hillen, H.S.; Kokic, G.; Farnung, L.; Dienemann, C.; Tegunov, D.; Cramer, P. Structure of replicating SARS-CoV-2 polymerase. Nature 2020, 584, 154–156. [Google Scholar] [CrossRef]
- Subissi, L.; Posthuma, C.C.; Collet, A.; Zevenhoven-Dobbe, J.C.; Gorbalenya, A.E.; Decroly, E.; Snijder, E.J.; Canard, B.; Imbert, I. One severe acute respiratory syndrome coronavirus protein complex integrates processive RNA polymerase and exonuclease activities. Proc. Natl. Acad. Sci. USA 2014, 111, E3900–E3909. [Google Scholar] [CrossRef]
- Chen, J.; Malone, B.; Llewellyn, E.; Grasso, M.; Shelton, P.M.M.; Olinares, P.D.B.; Maruthi, K.; Eng, E.T.; Vatandaslar, H.; Chait, B.T.; et al. Structural Basis for Helicase-Polymerase Coupling in the SARS-CoV-2 Replication-Transcription Complex. Cell 2020, 182, 1560–1573.e13. [Google Scholar] [CrossRef]
- Yan, L.; Zhang, Y.; Ge, J.; Zheng, L.; Gao, Y.; Wang, T.; Jia, Z.; Wang, H.; Huang, Y.; Li, M.; et al. Architecture of a SARS-CoV-2 mini replication and transcription complex. Nat. Commun. 2020, 11, 5874. [Google Scholar] [CrossRef]
- Ivanov, K.A.; Ziebuhr, J. Human Coronavirus 229E Nonstructural Protein 13: Characterization of Duplex-Unwinding, Nucleoside Triphosphatase, and RNA 5′-Triphosphatase Activities. J. Virol. 2004, 78, 7833–7838. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Cai, H.; Pan, J.; Xiang, N.; Tien, P.; Ahola, T.; Guo, D. Functional screen reveals SARS coronavirus nonstructural protein nsp14 as a novel cap N7 methyltransferase. Proc. Natl. Acad. Sci. USA 2009, 106, 3484–3489. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Fei, C.-Y.; Chen, Y.-P.; Sargsyan, K.; Chang, C.-P.; Yuan, H.S.; Lim, C. Synergistic Inhibition of SARS-CoV-2 Replication Using Disulfiram/Ebselen and Remdesivir. ACS Pharmacol. Transl. Sci. 2021, 4, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Seo, S.H.; Woo, S.-J.; Kwon, Y.; Song, M.; Ha, N.-C. Epigallocatechin gallate inhibits the uridylate-specific endoribonuclease Nsp15 and efficiently neutralizes the SARS-CoV-2 strain. J. Agric. Food Chem. 2021, 69, 5948–5954. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.R.; Misdary, C.; Yegya-Raman, N.; Kim, S.; Narayanan, N.; Siddiqui, S.; Salgame, P.; Radbel, J.; Groote, F.D.; Michel, C.; et al. Montelukast in hospitalized patients diagnosed with COVID-19. J. Asthma 2022, 59, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Wower, J.; Maltseva, N.; Chang, C.; Jedrzejczak, R.; Wilamowski, M.; Kang, S.; Nicolaescu, V.; Randall, G.; Michalska, K.; et al. Tipiracil binds to uridine site and inhibits Nsp15 endoribonuclease NendoU from SARS-CoV-2. Commun. Biol. 2021, 4, 193. [Google Scholar] [CrossRef]
- Nakagawa, K.; Narayanan, K.; Wada, M.; Makino, S. Inhibition of Stress Granule Formation by Middle East Respiratory Syndrome Coronavirus 4a Accessory Protein Facilitates Viral Translation, Leading to Efficient Virus Replication. J. Virol. 2018, 92, e00902-18. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.; Wu, A.; Wang, Y.; Xu, S.; Tang, Y.; Jin, X.; Wang, S.; Qin, L.; Sun, Y.; Fan, C.; et al. Identification and Characterization of a Ribose 2′-O-Methyltransferase Encoded by the Ronivirus Branch of Nidovirales. J. Virol. 2016, 90, 6675–6685. [Google Scholar] [CrossRef]
- Kumar, P.; Bhardwaj, T.; Giri, R. Mitoxantrone dihydrochloride, an FDA approved drug, binds with SARS-CoV-2 NSP1 C-terminal. RSC Adv. 2022, 12, 5648–5655. [Google Scholar] [CrossRef]
- Vankadari, N.; Jeyasankar, N.N.; Lopes, W.J. Structure of the SARS-CoV-2 Nsp1/5′-untranslated region complex and implications for potential therapeutic targets, a vaccine, and virulence. J. Phys. Chem. Lett. 2020, 11, 9659–9668. [Google Scholar] [CrossRef]
- Bujanic, L.; Shevchuk, O.; von Kügelgen, N.; Kalinina, A.; Ludwik, K.; Koppstein, D.; Zerna, N.; Sickmann, A.; Chekulaeva, M. The key features of SARS-CoV-2 leader and NSP1 required for viral escape of NSP1-mediated repression. RNA 2022, 28, 766–779. [Google Scholar] [CrossRef]
- Yan, W.; Zheng, Y.; Zeng, X.; He, B.; Cheng, W. Structural biology of SARS-CoV-2: Open the door for novel therapies. Signal Transduct. Target Ther. 2022, 7, 26. [Google Scholar] [CrossRef] [PubMed]
- Rut, W.; Lv, Z.; Zmudzinski, M.; Patchett, S.; Nayak, D.; Snipas, S.J.; El Oualid, F.; Huang, T.T.; Bekes, M.; Drag, M.; et al. Activity profiling and crystal structures of inhibitor-bound SARS-CoV-2 papain-like protease: A framework for anti-COVID-19 drug design. Sci. Adv. 2020, 6, eabd4596. [Google Scholar] [CrossRef]
- Hognon, C.; Marazzi, M.; García-Iriepa, C. Atomistic-level description of the covalent inhibition of SARS-CoV-2 papain-like protease. Int. J. Mol. Sci. 2022, 23, 5855. [Google Scholar] [CrossRef]
- Banerjee, S. An insight into the interaction between α-ketoamide- based inhibitor and coronavirus main protease: A detailed in silico study. Biophys. Chem. 2021, 269, 106510. [Google Scholar] [CrossRef] [PubMed]
- Frecer, V.; Miertus, S. Antiviral agents against COVID-19: Structure-based design of specific peptidomimetic inhibitors of SARS-CoV-2 main protease. RSC Adv. 2020, 10, 40244–40263. [Google Scholar] [CrossRef] [PubMed]
- Khater, S.; Kumar, P.; Dasgupta, N.; Das, G.; Ray, S.; Prakash, A. Combining SARS-CoV-2 proofreading exonuclease and RNA-dependent RNA polymerase inhibitors as a strategy to combat COVID-19: A high-throughput in silico screening. Front. Microbiol. 2021, 12, 647693. [Google Scholar] [CrossRef]
- Lin, L.; Wang, Y.; Li, Q.; Hu, M.; Shi, Y. Novel SARS-CoV-2 therapeutic targets: RNA proofreading complex and virus-induced senescence. Cell Death Differ. 2022, 29, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Rona, G.; Zeke, A.; Miwatani-Minter, B.; de Vries, M.; Kaur, R.; Schinlever, A.; Garcia, S.F.; Goldberg, H.V.; Wang, H.; Hinds, T.R.; et al. The NSP14/NSP10 RNA repair complex as a Pan-coronavirus therapeutic target. Cell Death Differ. 2022, 29, 285–292. [Google Scholar] [CrossRef]
- Lu, L.; Peng, Y.; Yao, H.; Wang, Y.; Li, J.; Yang, Y.; Lin, Z. Punicalagin as an allosteric NSP13 helicase inhibitor potently suppresses SARS-CoV-2 replication in vitro. Antivir. Res. 2022, 206, 105389. [Google Scholar] [CrossRef]
- Fan, H.; He, S.-T.; Han, P.; Hong, B.; Liu, K.; Li, M.; Wang, S.; Tong, Y. Cepharanthine: A promising old drug against SARS-CoV-2. Adv. Biol. 2022, 6, e2200148. [Google Scholar] [CrossRef]
- Narayanan, N.; Nair, D.T. Ritonavir may inhibit exoribonuclease activity of nsp14 from the SARS-CoV-2 virus and potentiate the activity of chain terminating drugs. Int. J. Biol. Macromol. 2021, 168, 272–278. [Google Scholar] [CrossRef]
- Mouffak, S.; Shubbar, Q.; Saleh, E.; El-Awady, R. Recent advances in management of COVID-19: A review. Biomed. Pharmacother. Biomed. Pharmacother. 2021, 143, 112107. [Google Scholar] [CrossRef]
- Kokic, G.; Hillen, H.S.; Tegunov, D.; Dienemann, C.; Seitz, F.; Schmitzova, J.; Farnung, L.; Siewert, A.; Höbartner, C.; Cramer, P. Mechanism of SARS-CoV-2 polymerase stalling by remdesivir. Nat. Commun. 2021, 12, 279. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Yin, W.; Xu, H.E. RNA-dependent RNA polymerase: Structure, mechanism, and drug discovery for COVID-19. Biochem. Biophys. Res. Commun. 2021, 538, 47–53. [Google Scholar] [CrossRef]
- Marzi, M.; Vakil, M.K.; Bahmanyar, M.; Zarenezhad, E. Paxlovid: Mechanism of action, synthesis, and in silico study. BioMed. Res. Int. 2022, 2022, 7341493. [Google Scholar] [CrossRef]
- Joyce, R.P.; Hu, V.W.; Wang, J. The history, mechanism, and perspectives of nirmatrelvir (PF-07321332): An orally bioavailable main protease inhibitor used in combination with ritonavir to reduce COVID-19-related hospitalizations. Med. Chem. Res. 2022, 31, 1637–1646. [Google Scholar] [CrossRef]
- Loos, N.H.C.; Beijnen, J.H.; Schinkel, A.H. The mechanism-based inactivation of CYP3A4 by ritonavir: What mechanism? Int. J. Mol. Sci. 2022, 23, 9866. [Google Scholar] [CrossRef] [PubMed]
- Kabinger, F.; Stiller, C.; Schmitzová, J.; Dienemann, C.; Kokic, G.; Hillen, H.S.; Höbartner, C.; Cramer, P. Mechanism of molnupiravir-induced SARS-CoV-2 mutagenesis. Nat. Struct. Mol. Biol. 2021, 28, 740–746. [Google Scholar] [CrossRef]
- Saravolatz, L.D.; Depcinski, S.; Sharma, M. Molnupiravir and nirmatrelvir-ritonavir: Oral coronavirus disease 2019 antiviral drugs. Clin. Infect. Dis. 2023, 76, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.J.; Tchesnokov, E.P.; Schinazi, R.F.; Götte, M. Molnupiravir promotes SARS-CoV-2 mutagenesis via the RNA template. J. Biol. Chem. 2021, 297, 100770. [Google Scholar] [CrossRef]
- Tian, L.; Pang, Z.; Li, M.; Lou, F.; An, X.; Zhu, S.; Song, L.; Tong, Y.; Fan, H.; Fan, J. Molnupiravir and its antiviral activity against COVID-19. Front. Immunol. 2022, 13, 855496. [Google Scholar] [CrossRef]
- Deeks, E.D. Casirivimab/Imdevimab: First Approval. Drugs 2021, 81, 2047–2055. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Lee, J.Y.; Yang, J.W.; Lee, K.H.; Effenberger, M.; Szpirt, W.; Kronbichler, A.; Shin, J.I. Immunopathogenesis and treatment of cytokine storm in COVID-19. Theranostics 2021, 11, 316–329. [Google Scholar] [CrossRef]
- Marconi, V.C.; Ramanan, A.V.; de Bono, S.; Kartman, C.E.; Krishnan, V.; Liao, R.; Piruzeli, M.L.B.; Goldman, J.D.; Alatorre-Alexander, J.; de Cassia Pellegrini, R.; et al. Efficacy and safety of baricitinib for the treatment of hospitalised adults with COVID-19 (COV-BARRIER): A randomised, double-blind, parallel-group, placebo-controlled phase 3 trial. Lancet Respir. Med. 2021, 9, 1407–1418. [Google Scholar] [CrossRef] [PubMed]
- Hoang, T.N.; Pino, M.; Boddapati, A.K.; Viox, E.G.; Starke, C.E.; Upadhyay, A.A.; Gumber, S.; Nekorchuk, M.; Busman-Sahay, K.; Strongin, Z.; et al. Baricitinib treatment resolves lower-airway macrophage inflammation and neutrophil recruitment in SARS-CoV-2-infected rhesus macaques. Cell 2021, 184, 460–475.e21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, Y.; Qiao, W.; Zhang, J.; Qi, Z. Baricitinib, a drug with potential effect to prevent SARS-CoV-2 from entering target cells and control cytokine storm induced by COVID-19. Int. Immunopharmacol. 2020, 86, 106749. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.T.S.; Šudomová, M.; Berchová-Bímová, K.; Šmejkal, K.; Echeverría, J. Psoromic Acid, a Lichen-Derived Molecule, Inhibits the Replication of HSV-1 and HSV-2, and Inactivates HSV-1 DNA Polymerase: Shedding Light on Antiherpetic Properties. Molecules 2019, 24, 2912. [Google Scholar] [CrossRef]
- Elion, G.B. Mechanism of action and selectivity of acyclovir. Am. J. Med. 1982, 73, 7–13. [Google Scholar] [CrossRef]
- Jones, G.; Prosser, D.E.; Kaufmann, M. Cytochrome P450-mediated metabolism of vitamin D. J. Lipid Res. 2014, 55, 13–31. [Google Scholar] [CrossRef]
- Campbell, G.R.; Spector, S.A. Hormonally Active Vitamin D3 (1α,25-Dihydroxycholecalciferol) Triggers Autophagy in Human Macrophages That Inhibits HIV-1 Infection *. J. Biol. Chem. 2011, 286, 18890–18902. [Google Scholar] [CrossRef]
- Lin, L.-T.; Chen, T.-Y.; Lin, S.-C.; Chung, C.-Y.; Lin, T.-C.; Wang, G.-H.; Anderson, R.; Lin, C.-C.; Richardson, C.D. Broad-spectrum antiviral activity of chebulagic acid and punicalagin against viruses that use glycosaminoglycans for entry. BMC Microbiol. 2013, 13, 187. [Google Scholar] [CrossRef]
- Yang, Y.; Xiu, J.; Liu, J.; Zhang, L.; Li, X.; Xu, Y.; Qin, C.; Zhang, L. Chebulagic Acid, a Hydrolyzable Tannin, Exhibited Antiviral Activity in Vitro and in Vivo against Human Enterovirus 71. Int. J. Mol. Sci. 2013, 14, 9618–9627. [Google Scholar] [CrossRef] [PubMed]
- Duncan, M.C.; Onguéné, P.A.; Kihara, I.; Nebangwa, D.N.; Naidu, M.E.; Williams, D.E.; Balgi, A.D.; Andrae-Marobela, K.; Roberge, M.; Andersen, R.J.; et al. Virtual Screening Identifies Chebulagic Acid as an Inhibitor of the M2(S31N) Viral Ion Channel and Influenza A Virus. Molecules 2020, 25, 2903. [Google Scholar] [CrossRef] [PubMed]
- Du, R.; Cooper, L.; Chen, Z.; Lee, H.; Rong, L.; Cui, Q. Discovery of chebulagic acid and punicalagin as novel allosteric inhibitors of SARS-CoV-2 3CLpro. Antivir. Res. 2021, 190, 105075. [Google Scholar] [CrossRef]
- Li, P.; Du, R.; Wang, Y.; Hou, X.; Wang, L.; Zhao, X.; Zhan, P.; Liu, X.; Rong, L.; Cui, Q. Identification of Chebulinic Acid and Chebulagic Acid as Novel Influenza Viral Neuraminidase Inhibitors. Front. Microbiol. 2020, 11, 182. [Google Scholar] [CrossRef]
- Liu, C.; Cai, D.; Zhang, L.; Tang, W.; Yan, R.; Guo, H.; Chen, X. Identification of hydrolyzable tannins (punicalagin, punicalin and geraniin) as novel inhibitors of hepatitis B virus covalently closed circular DNA. Antivir. Res. 2016, 134, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, G.; Nishioka, I.; Nishizawa, M.; Yamagishi, T.; Kashiwada, Y.; Dutschman, G.E.; Bodner, A.J.; Kilkuskie, R.E.; Cheng, Y.C.; Lee, K.H. Anti-AIDS agents, 2: Inhibitory effects of tannins on HIV reverse transcriptase and HIV replication in H9 lymphocyte cells. J. Nat. Prod. 1990, 53, 587–595. [Google Scholar] [CrossRef]


{kind=link}
| Protein | MW (kDa) | Amino Acid Length | Function | References |
|---|---|---|---|---|
| NSP1 | 19.8 | 180 | Binding 40S ribosome | [23] |
| NSP2 | 70.5 | 638 | Potential binding to prohibitins | [24] |
| NSP3 | 217.3 | 1945 | Papain-like protease | [14] |
| NSP4 | 56.2 | 500 | Formation of double-membrane vesicles (DMVs) replication organelles | [25] |
| NSP5 | 33.8 | 306 | 3C-like protease | [26] |
| NSP6 | 33.0 | 290 | Promotes the formation of double-membrane vesicles in infected cells, ER/cytosolic protein | [25] |
| NSP7 | 9.2 | 83 | Essential cofactor that binds to NSP12, forming a complex that stabilizes the polymerase domain | [27] |
| NSP8 | 21.9 | 198 | Extends the template RNA-binding surface | [27] |
| NSP9 | 12.4 | 113 | Involved in viral RNA synthesis | [28] |
| NSP10 | 14.8 | 139 | Viral replication | [29] |
| NSP11 | 1.3 | 13 | Unknown | [30] |
| NSP12 | 106.7 | 932 | Viral replication | [31] |
| NSP13 | 66.9 | 601 | Helicase | [32] |
| NSP14 | 59.8 | 527 | 3′ to 5′ ExoN activity | [33] |
| NSP15 | 38.8 | 346 | RNA processing | [34] |
| NSP16 | 33.3 | 298 | mRNA capping | [35] |
| Compound No. | Structure | Similar No. | Similar Structures |
|---|---|---|---|
| 1 Glycyrrhizic acid CID: 14982. CHEMBL441687. CAS: 1405-86-3. SMILE:C[C@@]12CC[C@@H]3[C@](C)(CC[C@H](O[C@H]4O[C@H](C(=O)O)[C@@H](O)[C@H](O)[C@H]4O[C@@H]4O[C@H](C(=O)O)[C@@H](O)[C@H](O)[C@H]4O)C3(C)C)[C@H]1C(=O)C=C1[C@@H]3C[C@](C)(CC[C@]3(C)CC[C@]12C)C(=O)O | 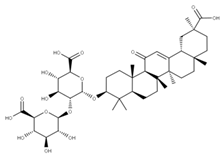 | 28 28-O-β-D-Glucopyranosyloleanolic acid 3-O-β-D-galactopyranosyl (1->3)-β-D-glucuronopyranoside-6-O-n-butyl ester. NPC104372. CID: 16099401. CHEMBL493640. SMILE: CCCCOC(=O)[C@@H]1[C@H]([C@@H]([C@H]([C@H](O[C@H]2CC[C@@]3(C)[C@@H](CC[C@]4(C)[C@@H]3CC=C3[C@@H]5CC(C)(C)CC[C@@]5(CC[C@@]43C)C(=O)O[C@H]3[C@@H]([C@H]([C@@H]([C@@H](CO)O3)O)O)O)C2(C)C)O1)O)O[C@H]1[C@@H]([C@H]([C@H]([C@@H](CO)O1)O)O)O)O | 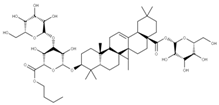 |
| 2 Lobaric acid CID: 73157. CHEMBL551842. CAS: 522-53-2 SMILE:O=C1Oc2cc(O)c(C(=O)O)c(CCCCC)c2Oc2cc(OC)cc(c12)C(=O)CCCC | 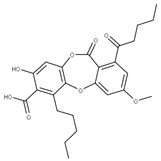 | 29 Psoromic acid NPC10576. CID: 23725. CHEMBL176570. CAS: 7299-11-8 SMILE: Cc1cc(c(C=O)c2c1C(=O)Oc1c(C)c(cc(c1O2)C(=O)O)OC)[O-] | 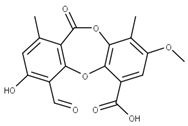 |
| 3 Garcinolic acid CID: 6857794. CHEMBL1456870. SMILE: CC1(Oc2c(C\C=C(/C)C)c3OC45C(CCC=C4C(=O)c3c(O)c2C=C1)C(C)(C)O[C@@]5(C\C=C(/C)C(=O)O)C(=O)O)CC\C=C(/C)C | 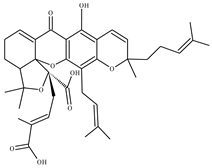 | 30 Neogambogic acid CID: 125071. CAS: 93772-31-7. SMILE: CC(=CCCC1(CC(C2=C(C3=C(C(=C2O1)CC=C(C)C)OC45C6CC(C=C4C3=O)C(=O)C5(OC6(C)C)CC=C(C)C(=O)O)O)O)C)C | 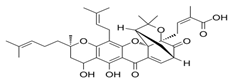 |
| 31 10-α-Hydroxyepigambogic acid CID: 74429469. SMILE: CC(=CCCC1(C=CC2=C(C3=C(C(=C2O1)CC=C(C)C)OC45C6CC(C(C4C3=O)O)C(=O)C5(OC6(C)C)CC=C(C)C(=O)O)O)C)C | 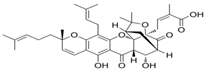 | ||
| 32 Gambogenific acid NPC162248. CID: 25208911. CHEMBL558353. SMILE: C/C(=CCc1c2O[C@H](Cc2c2c(c1O)C(=O)C1=C[C@@H]3C[C@@H]4[C@@]1(O2)[C@@](C/C=C(C(=O)O)/C)(OC4(C)C)C3=O)C(O)(C)C)/CCC=C(C)C. | 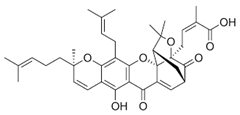 | ||
| 33 Nimbolinin C CID: 10794274. LTS0151616. SMILE: CC1=C2[C@H](C[C@H]1C3=COC=C3)O[C@H](C[C@H]4[C@]2([C@@H]([C@H]5[C@@H]6[C@@]4([C@H](C[C@H]([C@]6(CO5)C)OC(=O)C)OC(=O)/C=C/C7=CC=CC=C7)C)O)C)OC |  | ||
| 34 Scortechinone R NPC473607. CID: 44559271. CHEMBL447070. SMILE: CO[C@]12C=C3C(=O)c4c(O[C@@]53[C@H](C1)C(O[C@@]5(C2=O)C/C=C(C(=O)O)/C)(C)C)c1c(c(c4O)CC(C(=C)C)O)O[C@@H](C1(C)C)C | 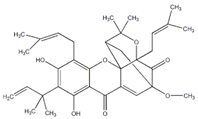 | ||
| 5 Montelukast CID: 5281040. CHEMBL787. CAS: 158966-92-8. SMILE: OC(C)(C)c1ccccc1CC[C@@H](SCC1(CC(=O)O)CC1)c1cccc(c1)/C=C/c1nc2cc(Cl)ccc2cc1 | 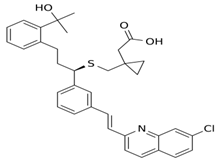 | No compound found | |
| 6 Mitoxantrone dihydrochloride CID: 164946820. CHEMBL5078683. SMILE: O=C(N[C@@H](CN)C(=O)NCC(=O)NC/C=C/C(=O)OC)[C@H](CCc1sc2ccccc2n1)NC(=O)C |  | No compound found | |
| 7 VIR251 CID: 164946820. CHEMBL5078683. SMILE: O=C(N[C@@H](CN)C(=O)NCC(=O)NC/C=C/C(=O)OC)[C@H](CCc1sc2ccccc2n1)NC(=O)C |  | No compound found | |
| 8 GRL-0617 CID: 24941262. CHEMBL549695. CAS: 1093070-16-6 SMILE: O=C(N[C@H](C)c1cccc2ccccc12)c1cc(N)ccc1C |  | No compound found | |
| 9 Vinyl sulfone-2CN115 CID: 156621389. SMILE: CS(=O)(=O)\C=C\[C@H](CCC(=O)N)NC(=O)[C@@H](NC(=O)OCc1cccc(C#C)c1)CC(C)C | 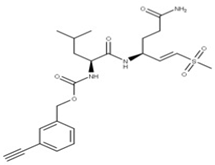 | No compound found | |
| 10 Calpain inhibitor I CID: 443118. CHEMBL304784. CAS: 110044-82-1. SMILE: CC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCC)C=O | 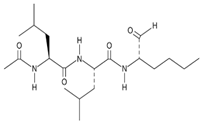 | No compound found | |
| 11 α-ketoamide 13b CID: 146026181. CHEMBL5075209. SMILE: O=C(OC(C)(C)C)NC1=CC=CN(C1=O)[C@@H](CC1CC1)C(=O)N[C@@H](C[C@@H]1CCNC1=O)C(=O)C(=O)NCc1ccccc1 | 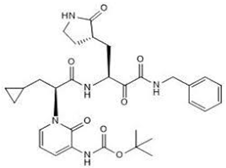 | No compound found | |
| 12 Metformin CID: 152743144. CAS: 657-24-9. SMILE: N=C(\N=C(/N)N)N(C)C | 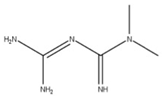 | No compound found | |
| 13 Vitamin D3 derivative CID: 52080453. CHEMBL846. CAS: 32222-06-3. SMILE: C[C@]12CCC\C(=C/C=C3/C[C@@H](O)C[C@H](O)C3=C)[C@@H]1CC[C@@H]2[C@H](C)CCCC(C)(O)C | 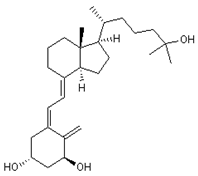 | 35 25-Hydroxycholecalciferol (calcifediol) NPC27395. CID: 5283731. CHEMBL1040. CAS: 19356-17-3 SMILE: O[C@H]1CCC(=C)/C(=CC=C2/CCC[C@]3([C@H]2CC[C@@H]3[C@@H](CCCC(O)(C)C)C)C)/C1 | 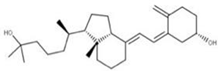 |
| 36 1α,24R,25-Trihydroxylcholecalciferol NPC320548. CID: 6446280. CHEMBL3351075. CAS: 56142-94-0. SMILE: O[C@H]1C[C@H](O)C(=C)/C(=CC=C2/CCC[C@]3([C@H]2CC[C@@H]3[C@@H](CC[C@H](C(O)(C)C)O)C)C)/C1 | 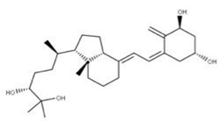 | ||
| 14 Conivaptan CID: 151171. CHEMBL1755. CAS: 210101-16-9. SMILE: O=C(N1CCc2[NH]c(C)nc2c2ccccc12)c1ccc(NC(=O)c2ccccc2c2ccccc2)cc1 | 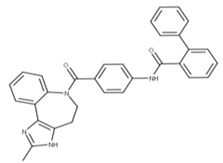 | No compound found | |
| 15 Sofalcone CID: 5282219. CHEMBL1441961. CAS: 64506-49-6. SMILE: C/C(C)=C/COc1cc(OCC(=O)O)c(cc1)C(=O)/C=C/c1ccc(cc1)OC\C=C(/C)C |  | 37 4-Geranyloxy-2,6 dihydroxybenzophenone NPC124269. CID: 44575287. CHEMBL510517. SMILE: C/C(=CCOc1cc(O)c(c(c1)O)C(=O)c1ccccc1)/CCC=C(C)C | 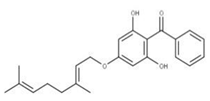 |
| 38 Cordoin NPC240593. CID: 5961410. CHEMBL450771. CAS: 38965-74-1 SMILE: CC(=CCOc1ccc(C(=O)/C=C/c2ccccc2)c(c1)O)C | 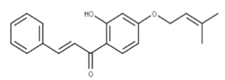 | ||
| 16 Punicalagin CID: 44584733. CAS: 65995-63-3. SMILE: OC1O[C@@H]2COC(=O)c3cc(O)c(O)c(O)c3c3c(O)c(O)c4OC(=O)c5c6c(OC(=O)c3c46)c(O)c(O)c5c3c(cc(O)c(O)c3O)C(=O)O[C@H]2[C@@H]2OC(=O)c3cc(O)c(O)c(O)c3c3c(cc(O)c(O)c3O)C(=O)O[C@@H]12 | 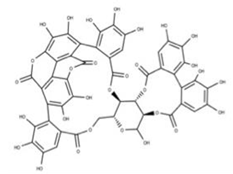 | 39 Chebulagic acid NPC119094. CID: 442674. CHEMBL525240 SMILE: OC(=O)C[C@@H]1C(=O)O[C@@H]2[C@H]3COC(=O)c4cc(O)c(c(c4c4c(C(=O)O[C@@H]2[C@@H](OC(=O)c2c5[C@H]1[C@H](O)C(=O)Oc5c(c(c2)O)O)[C@@H](O3)OC(=O)c1cc(O)c(c(c1)O)O)cc(O)c(c4O)O)O)O | 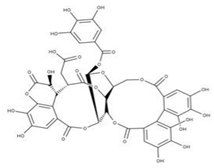 |
| 40 Punicalin NPC123259. CID: 5464368. CHEMBL502440. CAS: 65995-64-4. SMILE: O[C@@H]1O[C@@H]2COC(=O)c3cc(O)c(c(c3c3c(O)c(O)c4c5c3c(=O)oc3c(c(c(c6c(C(=O)O[C@H]2[C@@H]([C@H]1O)O)cc(O)c(c6O)O)c(c(=O)o4)c53)O)O)O)O | 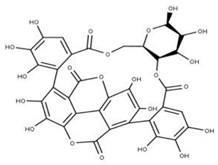 | ||
| 17 Cepharanthine CID: 10206. CHEMBL449782. CAS: 481-49-2 SMILE: COc1cc2CCN(C)[C@@H]3Cc4ccc(OC)c(Oc5ccc(cc5)C[C@@H]5N(C)CCc6cc7OCOc7c(Oc1cc32)c56)c4 | 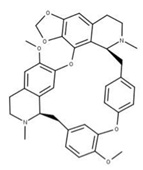 | 41 Isochondodendrine NPC104196. CID: 197726 CHEMBL1169628. CAS: 477-62-3. SMILE: CN1CCc2cc(c(c3c2[C@H]1Cc1ccc(cc1)Oc1c2c(CCN(C)[C@@H]2Cc2ccc(cc2)O3)cc(c1O)OC)O)OC | 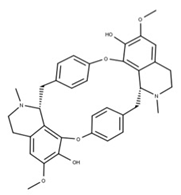 |
| 42 Isotetrandrine NPC10908. CID: 5351212. CHEMBL504757. CAS: 26137-48-4. SMILE: CN1CCc2cc(c3cc2[C@H]1Cc1ccc(cc1)Oc1cc(ccc1OC)C[C@H]1c2c(CCN1C)cc(c(c2O3)OC)OC)OC | 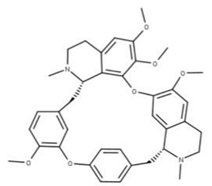 | ||
| 18 Ritonavir CID: 392622. CHEMBL163. CAS: 155213-67-5. SMILE: CN(Cc1nc(sc1)C(C)C)C(=O)N[C@H](C(=O)N[C@H](C[C@H](O)[C@@H](NC(=O)OCc1scnc1)Cc1ccccc1)Cc1ccccc1)C(C)C | 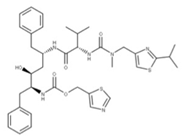 | No compound found | |
| 19 Disulfiram CID: 3117. CHEMBL964. CAS: 97-77-8. SMILE: CCN(CC)C(=S)SSC(=S)N(CC)CC | 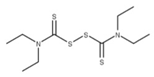 | No compound found | |
| 20 Ebselen CID: 3194. CHEMBL51085. CAS: 60940-34-3. SMILE: O=C1N([Se]c2ccccc12)c1ccccc1 | 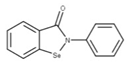 | No compound found | |
| 21 Sinefungin CID: 65482. CHEMBL1214186. CAS: 58944-73-3. SMILE: O=C(O)[C@@H](N)CC[C@H](N)C[C@H]1O[C@@H](n2cnc3c2ncnc3N)[C@H](O)[C@@H]1O |  | 43 S-Adenosylhomocysteine NPC324484. CID: 439155. CHEMBL418052. CAS: 979-92-0. SMILE: C1=NC(=C2C(=N1)N(C=N2)C3C(C(C(O3)CSCCC(C(=O)[O-])[NH3+])O)O)N | 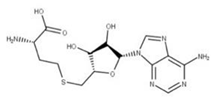 |
| 22 Epigallocatechin gallate CID: 65064. CHEMBL297453. CAS: 989-51-5. SMILE: O=C(O[C@@H]1Cc2c(cc(O)cc2O)O[C@@H]1c1cc(O)c(O)c(O)c1)c1cc(O)c(O)c(O)c1 | 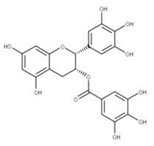 | 44 3,4,5-Trihydroxy-benzoic acid (2R,3S)-5,7-dihydroxy-2-(3,4,5-trihydroxy-phenyl)-chroman-3-Yl ester NPC104983. CID: 5276890. CHEMBL126079. CAS: 5127-64-0. SMILE: Oc1cc(O)c2c(c1)O[C@@H]([C@H](C2)OC(=O)c1cc(O)c(c(c1)O)O)c1cc(O)c(c(c1)O)O |  |
| 45 (-)-Epigallocatechin gallate NPC291948. CID: 2824823. CHEMBL338988 SMILE: Oc1cc(O)c2c(c1)O[C@H]([C@H](C2)OC(=O)c1cc(O)c(c(c1)O)O)c1cc(O)c(c(c1)O)O |  | ||
| 23 Tipiracil CID: 6323266. CHEMBL235668. CAS: 183204-74-2. SMILE: O=C1NC(=O)C(Cl)=C(CN2CCCC2=N)N1 |  | No compound found | |
| 24 Remdesivir CID: 121304016. CHEMBL4065616. CAS: 1809249-37-3. SMILE: O=[P@](Oc1ccccc1)(OC[C@H]1O[C@](C#N)([C@H](O)[C@@H]1O)c1ccc2n1ncnc2N)N[C@@H](C)C(=O)OCC(CC)CC | 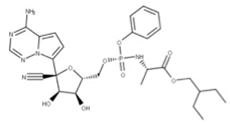 | No compound found | |
| 25 Nirmatrelvir CID: 155903259. CHEMBL4802135. CAS: 2628280-40-8. SMILE: CC1(C)[C@@H]2[C@H](N(C[C@@H]21)C(=O)[C@@H](NC(=O)C(F)(F)F)C(C)(C)C)C(=O)N[C@H](C#N)C[C@@H]1CCNC1=O | 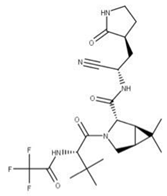 | No compound found | |
| 26 Molnupiravir CID: 145996610. CHEMBL4650320. CAS: 2492423-29-5. SMILE: ONC=1C=CN(C(=O)N=1)[C@@H]1O[C@H](COC(=O)C(C)C)[C@@H](O)[C@H]1O | 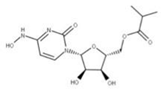 | No compound found | |
| 27 Baricitinib CID: 44205240. CHEMBL2105759. CAS: 1187594-09-7. SMILE: O=S(=O)(CC)N1CC(CC#N)(C1)n1cc(cn1)c1ncnc2[NH]ccc21 | 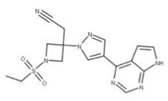 | No compound found |
| Compound | Similar Structures | Source |
|---|---|---|
| 28 | 28-O-b-D-Glucopyranosyloleanolic acid 3-O-b-D-galactopyranosyl(1->3)-b-D-glucuronopyranoside-6-O-n-butyl ester | Calendula officinalis (P) |
| Licorice saponins (several compounds) | Glycyrrhiza uralensis (P) | |
| 29 | Psoromic acid | Adiantum capillus-veneris (P) Aesculus pavia (P) Antrodia sinuosa (F) Calea nelsonii (P) Colletotrichum acutatum (F) Cephalocroton cordofanus (P) Dendrobium moniliforme (P) Lamium galeobdolon subsp. montanum (P) |
| 30 | Neogambogic acid | Garcinia hanburyi (P) |
| 31 | 10-a-Hydroxyepigambogic acid | Garcinia hanburyi (P) |
| 32 | Gambogenific acid | Garcinia hanburyi (P) |
| 33 | Nimbolinin C | Melia azedarach (P) |
| 34 | Scortechinone R | Garcinia scortechinii (P) |
| 35 | 25-Hydroxycholecalciferol (calcifediol) | Homo sapiens (M) |
| 36 | 1-a,24R,25-Trihydroxylcholecalciferol | Homo sapiens (M) |
| 37 | 4-Geranyloxy-2,6-dihydroxybenzophenone | Cratoxylum sumatranum (P) Pseudosuberites hyalinus (S) Ribes rubrum (P) Scorzonera judaica (P) Tovomita longifolia (P) Verbena brasiliensis (P) |
| 38 | Cordoin | Crotalaria dura (P) Duguetia staudtii (P) Fasciospongia cavernosa (S) Liquidambar orientalis (P) Millettia erythrocalyx (P) Salvia phlomoides (P) Solidago arguta (P) |
| 39 | Chebulagic acid | Bupleurum chinense (P) Excoecaria agallocha (P) Leonurus sibiricus (P) Lumbricus terrestris (W) Phyllanthus emblica (P) |
| 40 | Punicalin | Aspergillus candidus (F) Beauveria felina (F) Conyza canadensis (P) Cirsium japonicum (P) Cladiella krempfi (C) Garcinia hombroniana (P) Punica granatum (P) |
| 41 | Isochondodendrine | Coffea canephora (P) Collybia albuminosa (F) Cyclea barbata (P) Cyclea racemose (P) Isolona ghesquiereina (P) Megaselia halterata (F) Mustela putorius furo (M) |
| 42 | Isotetrandrine | Cornus officinalis (P) Cyclea barbata (P) Melianthus major (P) Pestalotiopsis disseminate (F) Stephania cepharantha (P) Stephania tetrandra (P) Thalictrum faberi (P) |
| 43 | S-Adenosylhomocysteine | Homo sapiens (M) |
| 44 | 3,4,5-Trihydroxy-benzoic acid (2R,3S)-5,7-dihydroxy-2-(3,4,5-trihydroxy-phenyl)-chroman-3-Yl ester | Corylus avellana (P) Morus alba (P) Paeonia lactiflora (P) |
| 45 | (-)-Epigallocatechin gallate | Camellia sinensis (P) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tam, D.; Lorenzo-Leal, A.C.; Hernández, L.R.; Bach, H. Targeting SARS-CoV-2 Non-Structural Proteins. Int. J. Mol. Sci. 2023, 24, 13002. https://doi.org/10.3390/ijms241613002
Tam D, Lorenzo-Leal AC, Hernández LR, Bach H. Targeting SARS-CoV-2 Non-Structural Proteins. International Journal of Molecular Sciences. 2023; 24(16):13002. https://doi.org/10.3390/ijms241613002
Chicago/Turabian StyleTam, Donald, Ana C. Lorenzo-Leal, Luis Ricardo Hernández, and Horacio Bach. 2023. "Targeting SARS-CoV-2 Non-Structural Proteins" International Journal of Molecular Sciences 24, no. 16: 13002. https://doi.org/10.3390/ijms241613002
APA StyleTam, D., Lorenzo-Leal, A. C., Hernández, L. R., & Bach, H. (2023). Targeting SARS-CoV-2 Non-Structural Proteins. International Journal of Molecular Sciences, 24(16), 13002. https://doi.org/10.3390/ijms241613002