
Targeting NKG2DL with Bispecific NKG2D–CD16 and NKG2D–CD3 Fusion Proteins on Triple–Negative Breast Cancer

 , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Characterization of NKG2DL Expression in Triple–Negative Breast Cancer Cells
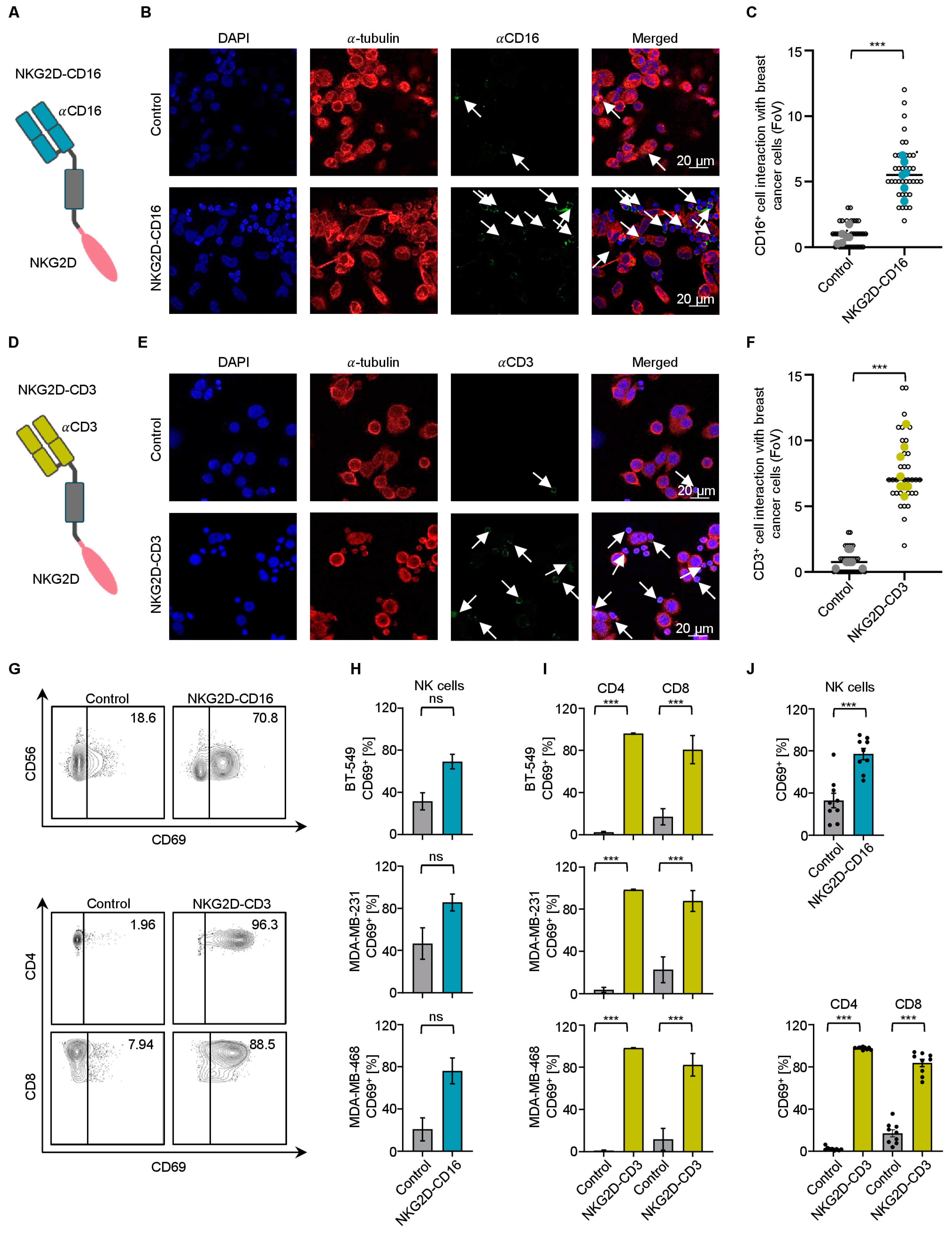
2.2. Effector Cell Recruitment by NKG2D BFPs against TNBC Cells
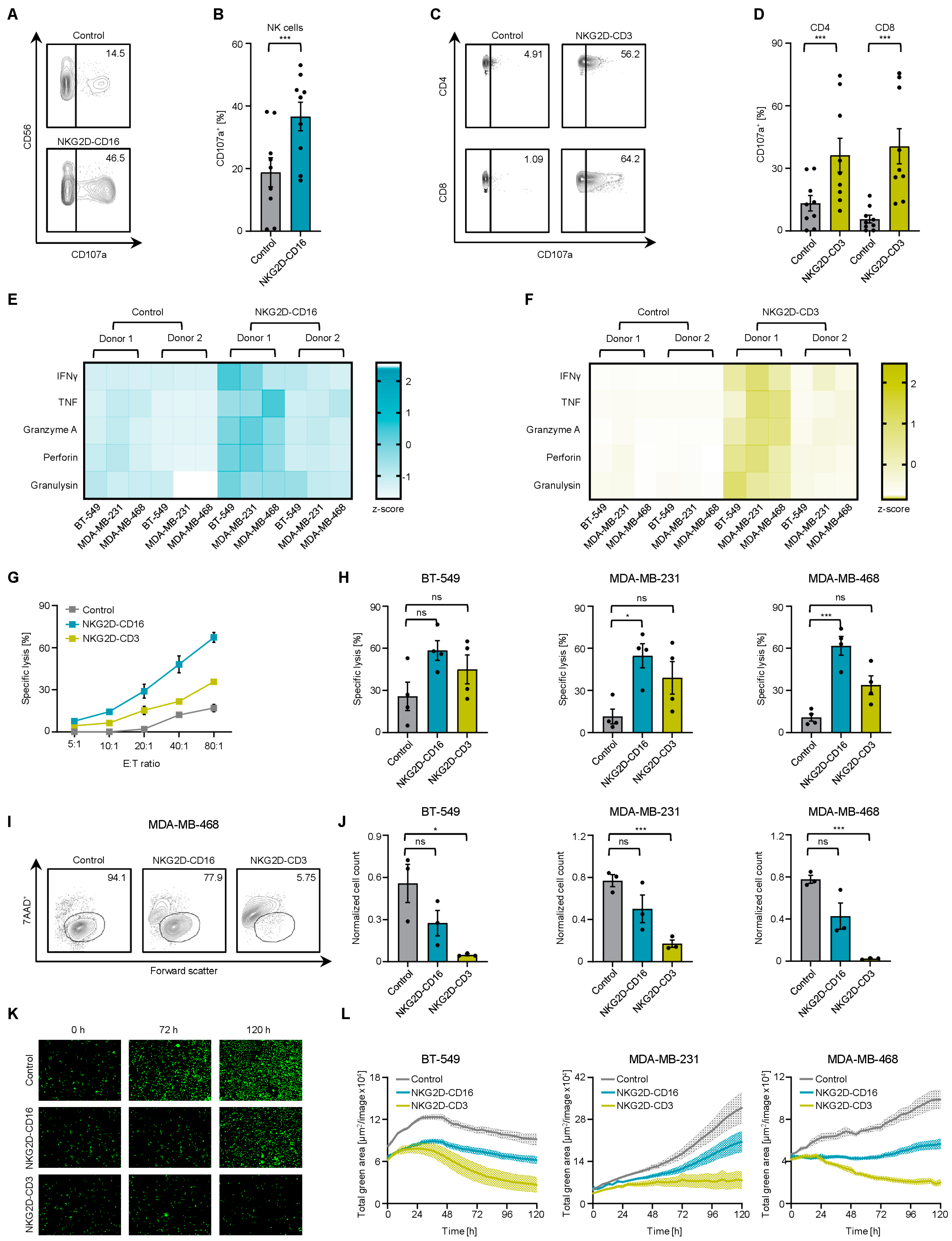
2.3. Modulation of NK Cell and T Cell Reactivity against TNBC Cells with PBMCs from Healthy Donors
2.4. Induction of Target Cell Lysis by Bispecific NKG2D Fusion Proteins
2.5. Reactivity of NK Cells and T Cells from TNBC Patients against TNBC Cells
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. qRT–PCR
4.3. Flow Cytometry
4.4. Production and Purification of Bispecific NKG2D Fusion Proteins
4.5. Immunofluorescence
4.6. Analysis of Cytokine Secretion
4.7. Cytotoxicity Assay
4.8. Primary Material
4.9. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Giaquinto, A.N.; Sung, H.; Miller, K.D.; Kramer, J.L.; Newman, L.A.; Minihan, A.; Jemal, A.; Siegel, R.L. Breast Cancer Statistics, 2022. CA Cancer J. Clin. 2022, 72, 524–541. [Google Scholar] [CrossRef] [PubMed]
- Li, X.X.; Yang, J.; Peng, L.M.; Sahin, A.A.; Huo, L.; Ward, K.C.; O’Regan, R.; Torres, M.A.; Meisel, J.L. Triple Negative Breast Cancer Has Worse Overall Survival and Cause–Specific Survival Than Non–Triple Negative Breast Cancer. Mod. Pathol. 2017, 30, 54a. [Google Scholar] [CrossRef] [PubMed]
- Anurag, M.; Jaehnig, E.J.; Krug, K.; Lei, J.T.; Bergstrom, E.J.; Kim, B.J.; Vashist, T.D.; Huynh, A.M.T.; Dou, Y.; Gou, X.; et al. Proteogenomic Markers of Chemotherapy Resistance and Response in Triple–Negative Breast Cancer. Cancer Discov. 2022, 12, 2586–2605. [Google Scholar] [CrossRef]
- Yao, H.; He, G.; Yan, S.; Chen, C.; Song, L.; Rosol, T.J.; Deng, X. Triple–negative breast cancer: Is there a treatment on the horizon? Oncotarget 2017, 8, 1913–1924. [Google Scholar] [CrossRef] [PubMed]
- Bagegni, N.A.; Davis, A.A.; Clifton, K.K.; Ademuyiwa, F.O. Targeted Treatment for High–Risk Early–Stage Triple–Negative Breast Cancer: Spotlight on Pembrolizumab. Breast Cancer 2022, 14, 113–123. [Google Scholar] [CrossRef]
- Raulet, D.H.; Gasser, S.; Gowen, B.G.; Deng, W.; Jung, H. Regulation of ligands for the NKG2D activating receptor. Annu. Rev. Immunol. 2013, 31, 413–441. [Google Scholar] [CrossRef]
- Orr, M.T.; Lanier, L.L. Natural killer cell education and tolerance. Cell 2010, 142, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Zingoni, A.; Fionda, C.; Borrelli, C.; Cippitelli, M.; Santoni, A.; Soriani, A. Natural Killer Cell Response to Chemotherapy–Stressed Cancer Cells: Role in Tumor Immunosurveillance. Front. Immunol. 2017, 8, 1194. [Google Scholar] [CrossRef]
- Han, Y.; Xie, W.; Song, D.G.; Powell, D.J., Jr. Control of triple–negative breast cancer using ex vivo self–enriched, costimulated NKG2D CAR T cells. J. Hematol. Oncol. 2018, 11, 92. [Google Scholar] [CrossRef] [PubMed]
- Sallman, D.A.; Brayer, J.; Sagatys, E.M.; Lonez, C.; Breman, E.; Agaugue, S.; Verma, B.; Gilham, D.E.; Lehmann, F.F.; Davila, M.L. NKG2D–based chimeric antigen receptor therapy induced remission in a relapsed/refractory acute myeloid leukemia patient. Haematologica 2018, 103, e424–e426. [Google Scholar] [CrossRef] [PubMed]
- Sallman, D.A.; Kerre, T.; Havelange, V.; Poire, X.; Lewalle, P.; Wang, E.S.; Brayer, J.B.; Davila, M.L.; Moors, I.; Machiels, J.P.; et al. CYAD–01, an autologous NKG2D–based CAR T–cell therapy, in relapsed or refractory acute myeloid leukaemia and myelodysplastic syndromes or multiple myeloma (THINK): Haematological cohorts of the dose escalation segment of a phase 1 trial. Lancet Haematol. 2023, 10, e191–e202. [Google Scholar] [CrossRef] [PubMed]
- Raab, S.; Steinbacher, J.; Schmiedel, B.J.; Kousis, P.C.; Steinle, A.; Jung, G.; Grosse-Hovest, L.; Salih, H.R. Fc–optimized NKG2D–Fc constructs induce NK cell antibody–dependent cellular cytotoxicity against breast cancer cells independently of HER2/neu expression status. J. Immunol. 2014, 193, 4261–4272. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, O.A.; Gonzalez-Hinojosa, M.D.R.; Arakawa-Hoyt, J.S.; Millan, A.J.; Gotthardt, D.; Nabekura, T.; Lanier, L.L. The CD16 and CD32b Fc–gamma receptors regulate antibody–mediated responses in mouse natural killer cells. J. Leukoc. Biol. 2023, 113, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Hagelstein, I.; Lutz, M.S.; Schmidt, M.; Heitmann, J.S.; Malenke, E.; Zhou, Y.; Clar, K.L.; Kopp, H.G.; Jung, G.; Salih, H.R.; et al. Bispecific NKG2D–CD3 and NKG2D–CD16 Fusion Proteins as Novel Treatment Option in Advanced Soft Tissue Sarcomas. Front. Immunol. 2021, 12, 653081. [Google Scholar] [CrossRef] [PubMed]
- Marklin, M.; Hagelstein, I.; Koerner, S.P.; Rothfelder, K.; Pfluegler, M.S.; Schumacher, A.; Grosse-Hovest, L.; Jung, G.; Salih, H.R. Bispecific NKG2D–CD3 and NKG2D–CD16 fusion proteins for induction of NK and T cell reactivity against acute myeloid leukemia. J. Immunother. Cancer 2019, 7, 143. [Google Scholar] [CrossRef]
- Rosenberg, J.; Huang, J. CD8+ T Cells and NK Cells: Parallel and Complementary Soldiers of Immunotherapy. Curr. Opin. Chem. Eng. 2018, 19, 9–20. [Google Scholar] [CrossRef]
- Thacker, G.; Henry, S.; Nandi, A.; Debnath, R.; Singh, S.; Nayak, A.; Susnik, B.; Boone, M.M.; Zhang, Q.; Kesmodel, S.B.; et al. Immature natural killer cells promote progression of triple–negative breast cancer. Sci. Transl. Med. 2023, 15, eabl4414. [Google Scholar] [CrossRef] [PubMed]
- Moretta, L. Dissecting CD56dim human NK cells. Blood 2010, 116, 3689–3691. [Google Scholar] [CrossRef]
- Sheppard, S.; Ferry, A.; Guedes, J.; Guerra, N. The Paradoxical Role of NKG2D in Cancer Immunity. Front. Immunol. 2018, 9, 1808. [Google Scholar] [CrossRef] [PubMed]
- Paczulla, A.M.; Rothfelder, K.; Raffel, S.; Konantz, M.; Steinbacher, J.; Wang, H.; Tandler, C.; Mbarga, M.; Schaefer, T.; Falcone, M.; et al. Absence of NKG2D ligands defines leukaemia stem cells and mediates their immune evasion. Nature 2019, 572, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Beverley, P.C.; Callard, R.E. Distinctive functional characteristics of human “T” lymphocytes defined by E rosetting or a monoclonal anti–T cell antibody. Eur. J. Immunol. 1981, 11, 329–334. [Google Scholar] [CrossRef]
- Fleit, H.B.; Wright, S.D.; Unkeless, J.C. Human neutrophil Fc gamma receptor distribution and structure. Proc. Natl. Acad. Sci. USA 1982, 79, 3275–3279. [Google Scholar] [CrossRef] [PubMed]
- Shields, R.L.; Namenuk, A.K.; Hong, K.; Meng, Y.G.; Rae, J.; Briggs, J.; Xie, D.; Lai, J.; Stadlen, A.; Li, B.; et al. High resolution mapping of the binding site on human IgG1 for Fc gamma RI, Fc gamma RII, Fc gamma RIII, and FcRn and design of IgG1 variants with improved binding to the Fc gamma R. J. Biol. Chem. 2001, 276, 6591–6604. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Oldham, R.J.; Teal, E.; Beers, S.A.; Cragg, M.S. Fc–Engineering for Modulated Effector Functions–Improving Antibodies for Cancer Treatment. Antibodies 2020, 9, 64. [Google Scholar] [CrossRef] [PubMed]
- Herbener, P.; Schonfeld, K.; Konig, M.; Germer, M.; Przyborski, J.M.; Bernoster, K.; Schuttrumpf, J. Functional relevance of in vivo half antibody exchange of an IgG4 therapeutic antibody–drug conjugate. PLoS ONE 2018, 13, e195823. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Characteristics | Total (n = 19) | |
|---|---|---|
| n (%) | Mean (Stdv.) | |
| Gender Female Age, Mean Years | 19 (100) | 59.4 (±10.4) |
| TNM classification | ||
| T1c N0 MX T1c N0 M0 T1c N1 MX T1c N1 M0 T2 N0 MX T2 N0 M0 T2 N1 MX T2 N1 M0 T2 N1 M1 T3 N0 MX T3 N1 M0 T3 N2a M0 T4d N1 MX | 2 (11) 2 (11) 1 (5) 1 (5) 2 (11) 4 (21) 1 (5) 1 (5) 1 (5) 1 (5) 1 (5) 1 (5) 1 (5) | |
| Histological subtype | ||
| ER/PR status negative <10% 10–50% | 15 (79) 3 (16) 1 (5) | |
| Her2 neu status negative + ++ | 12 (63) 3 (16) 2 (11) | |
| Histological grading | ||
| G2 G3 | 8 (42) 11 (58) | |
| Therapy line at the time of blood sampling | ||
| Atezolizumab Carboplatin, Paclitaxel Carboplatin, Paclitaxel, Atezolizumab Carboplatin, Paclitaxel, Pembrolizumab Cyclophosphamide, Epirubicin Denosumab Denosumab, Paclitaxel, Pembrolizumab Sacituzumab | 1 (5) 3 (16) 1 (5) 7 (37) 2 (11) 1 (5) 1 (5) 1 (5) | |
| Days since last treatment dose | ||
| d7 d8 d14 d21 d34 | 12 (63) 2 (11) 1 (5) 2 (11) 1 (5) | |
| Peripheral blood count | ||
| Leukocytes (1000/µL) Lymphocytes (1000/µL) Monocytes (1000/µL) Neutrophils (1000/µL) Platelet count (1000/µL) Hb (g/dL) | 5.2 (±2.4) 1.8 (±0.6) 0.5 (±0.15) 3.1 (±1.9) 275 (±143) 11.1 (±0.9) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaidun, P.; Holzmayer, S.J.; Greiner, S.M.; Seller, A.; Tegeler, C.M.; Hagelstein, I.; Mauermann, J.; Engler, T.; Koch, A.; Hartkopf, A.D.; et al. Targeting NKG2DL with Bispecific NKG2D–CD16 and NKG2D–CD3 Fusion Proteins on Triple–Negative Breast Cancer. Int. J. Mol. Sci. 2023, 24, 13156. https://doi.org/10.3390/ijms241713156
Kaidun P, Holzmayer SJ, Greiner SM, Seller A, Tegeler CM, Hagelstein I, Mauermann J, Engler T, Koch A, Hartkopf AD, et al. Targeting NKG2DL with Bispecific NKG2D–CD16 and NKG2D–CD3 Fusion Proteins on Triple–Negative Breast Cancer. International Journal of Molecular Sciences. 2023; 24(17):13156. https://doi.org/10.3390/ijms241713156
Chicago/Turabian StyleKaidun, Polina, Samuel J. Holzmayer, Sarah M. Greiner, Anna Seller, Christian M. Tegeler, Ilona Hagelstein, Jonas Mauermann, Tobias Engler, André Koch, Andreas D. Hartkopf, and et al. 2023. "Targeting NKG2DL with Bispecific NKG2D–CD16 and NKG2D–CD3 Fusion Proteins on Triple–Negative Breast Cancer" International Journal of Molecular Sciences 24, no. 17: 13156. https://doi.org/10.3390/ijms241713156
APA StyleKaidun, P., Holzmayer, S. J., Greiner, S. M., Seller, A., Tegeler, C. M., Hagelstein, I., Mauermann, J., Engler, T., Koch, A., Hartkopf, A. D., Salih, H. R., & Märklin, M. (2023). Targeting NKG2DL with Bispecific NKG2D–CD16 and NKG2D–CD3 Fusion Proteins on Triple–Negative Breast Cancer. International Journal of Molecular Sciences, 24(17), 13156. https://doi.org/10.3390/ijms241713156