
TMT-Based Proteomics Analysis of Senescent Nucleus Pulposus from Patients with Intervertebral Disc Degeneration

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Screening of Senescent NP Samples
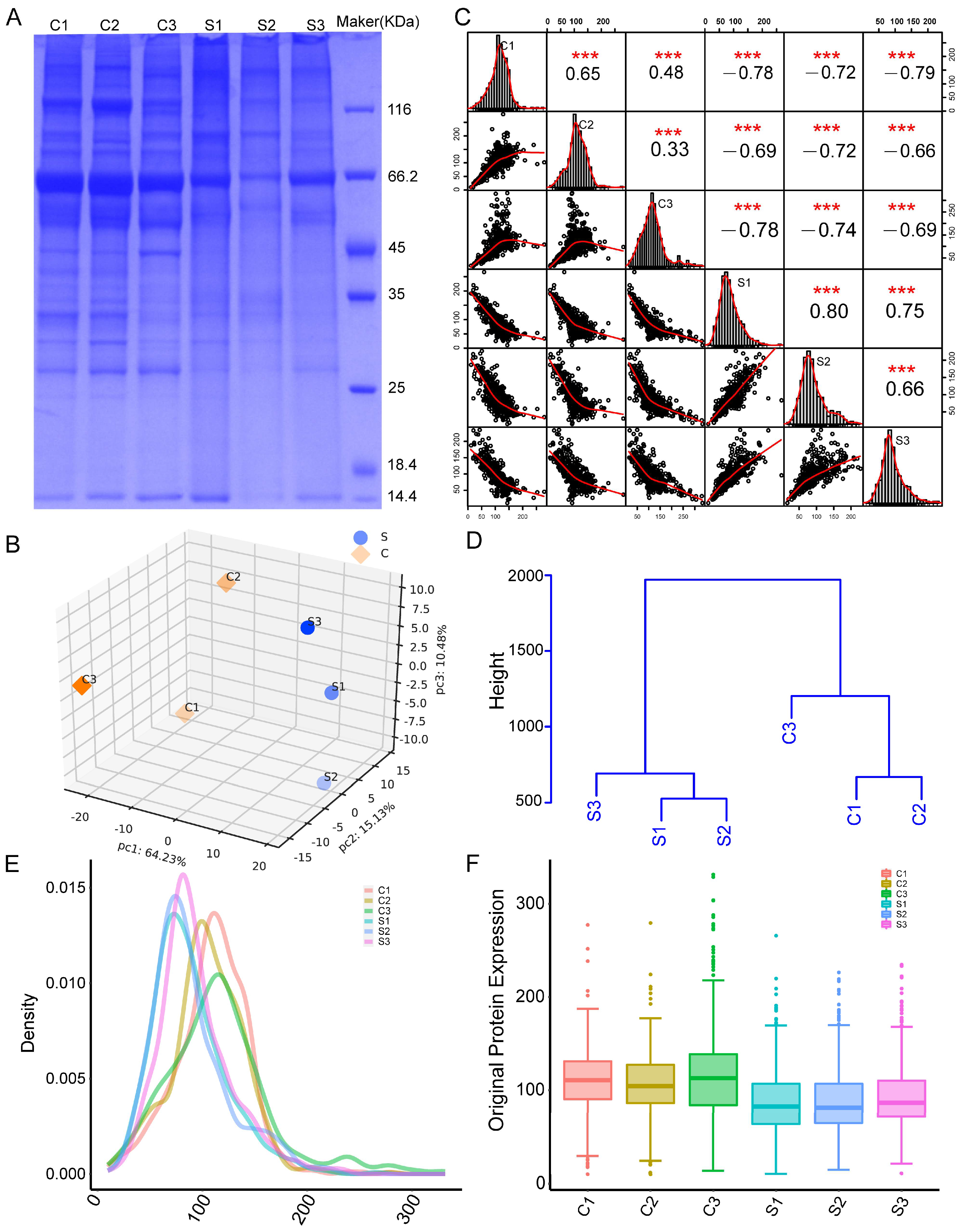
2.2. NP Sample Quality Control
2.3. TMT Quantitative Proteomic Analysis and Quality Control of Data
2.4. Identification of DEPs
2.5. GO and KEGG Annotation Analyses of DEPs
2.6. Protein–Protein Interaction (PPI) Network and Experimental Verification of DEPs
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Histological Evaluation
4.3. Western Blot
4.4. Protein Extraction, Digestion, and Quality Control
4.5. Tandem Mass Tag (TMT) Labeling
4.6. Liquid Chromatography–Mass Spectrometry/Mass Spectrometry (LC-MS/MS) Analysis
4.7. Database Search and Bioinformatics
4.8. Extraction of Human-Derived NP Cells
4.9. Senescence-Associated β-Galactosidase (SA-β-Gal) Staining and Immunofluorescence
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dudek, M.; Morris, H.; Rogers, N.; Pathiranage, D.R.; Raj, S.S.; Chan, D.; Kadler, K.E.; Hoyland, J.; Meng, Q.J. The clock transcription factor BMAL1 is a key regulator of extracellular matrix homeostasis and cell fate in the intervertebral disc. Matrix Biol. 2023, 122, 1–9. [Google Scholar] [CrossRef]
- Ohnishi, T.; Tran, V.; Sao, K.; Ramteke, P.; Querido, W.; Barve, R.A.; van de Wetering, K.; Risbud, M.V. Loss of function mutation in Ank causes aberrant mineralization and acquisition of osteoblast-like-phenotype by the cells of the intervertebral disc. Cell Death Dis. 2023, 14, 447. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.H.; Kim, H.S.; Jang, I.T. Intervertebral Disc Diseases PART 2: A Review of the Current Diagnostic and Treatment Strategies for Intervertebral Disc Disease. Int. J. Mol. Sci. 2020, 21, 2135. [Google Scholar] [CrossRef] [PubMed]
- Silwal, P.; Nguyen-Thai, A.M.; Mohammad, H.A.; Wang, Y.; Robbins, P.D.; Lee, J.Y.; Vo, N.V. Cellular senescence in intervertebral disc aging and degeneration: Moleular mechanisms and potential therapeutic opportunities. Biomolecules 2023, 13, 686. [Google Scholar] [CrossRef]
- Yang, G.; Liu, X.; Jing, X.; Wang, J.; Wang, H.; Chen, F.; Wang, W.; Shao, Y.; Cui, X. Astaxanthin suppresses oxidative stress and calcification in vertebral cartilage endplate via activating Nrf-2/HO-1 signaling pathway. Int. Immunopharmacol. 2023, 119, 110159. [Google Scholar] [CrossRef]
- Välikangas, T.; Suomi, T.; Elo, L.L. A systematic evaluation of normalization methods in quantitative label-free proteomics. Brief. Bioinform. 2018, 19, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ottone, O.K.; Kim, C.J.; Collins, J.A.; Risbud, M.V. Corrigendum: The cGAS-STING pathway affects vertebral bone but does not promote intervertebral disc cell senescence or degeneration. Front. Immunol. 2023, 14, 1201655. [Google Scholar] [CrossRef]
- Yilmaz, I.; Akalan, H.; Oznam, K.; Karaarslan, N.; Yasar Sirin, D.; Ozbek, H. Does oseltamivir protect human chondrocyte and nucleus pulposus cells from degeneration by inhibiting senescence and proinflammation mediated by the NLRP3 inflammasome and NF-κB? Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 4816–4827. [Google Scholar] [PubMed]
- Cherif, H.; Mannarino, M.; Pacis, A.S.; Ragoussis, J.; Rabau, O.; Ouellet, J.A.; Haglund, L. Single-cell RNA-Seq analysis of cells from degenerating and non-degenerating intervertebral discs from the same individual reveals new biomarkers for intervertebral disc degeneration. Int. J. Mol. Sci. 2022, 23, 3993. [Google Scholar] [CrossRef] [PubMed]
- Novais, E.J.; Diekman, B.O.; Shapiro, I.M.; Risbud, M.V. p16(Ink4a) deletion in cells of the intervertebral disc affects their matrix homeostasis and senescence associated secretory phenotype without altering onset of senescence. Matrix Biol. 2019, 82, 54–70. [Google Scholar] [CrossRef]
- Herman, A.B.; Tsitsipatis, D.; Anerillas, C.; Mazan-Mamczarz, K.; Carr, A.E.; Gregg, J.M.; Wang, M.; Zhang, J.; Michel, M.; Henry-Smith, C.A.; et al. DPP4 inhibition impairs senohemostasis to improve plaque stability in atherosclerotic mice. J. Clin. Investig. 2023, 133, e165933. [Google Scholar] [CrossRef] [PubMed]
- Schlett, J.S.; Mettang, M.; Skaf, A.; Schweizer, P.; Errerd, A.; Mulugeta, E.A.; Hein, T.M.; Tsesmelis, K.; Tsesmelis, M.; Büttner, U.F.G.; et al. NF-κB is a critical mediator of post-mitotic senescence in oligodendrocytes and subsequent white matter loss. Mol. Neurodegener. 2023, 18, 24. [Google Scholar] [CrossRef] [PubMed]
- Cherif, H.; Bisson, D.G.; Mannarino, M.; Rabau, O.; Ouellet, J.A.; Haglund, L. Senotherapeutic drugs for human intervertebral disc degeneration and low back pain. eLife 2020, 9, e54693. [Google Scholar] [CrossRef] [PubMed]
- Pinto, A.F.M.; Diedrich, J.K.; Moresco, J.J.; Yates, J.R. Differential precipitation of proteins: A simple protein fractionation strategy to gain biological insights with proteomics. J. Am. Soc. Mass. Spectrom. 2023. [Google Scholar] [CrossRef] [PubMed]
- Escal, J.; Neel, T.; Hodin, S.; Boussoualim, K.; Amouzougan, A.; Coassy, A.; Locrelle, H.; Thomas, T.; Delavenne, X.; Marotte, H. Proteomics analyses of human plasma reveal triosephosphate isomerase as a potential blood marker of methotrexate resistance in rheumatoid arthritis. Rheumatology 2023, 8, kead390. [Google Scholar] [CrossRef]
- Pan, S.; Ding, A.; Li, Y.; Sun, Y.; Zhan, Y.; Ye, Z.; Song, N.; Peng, B.; Li, L.; Huang, W.; et al. Small-molecule probes from bench to bedside: Advancing molecular analysis of drug-target interactions toward precision medicine. Chem. Soc. Rev. 2023, 52, 5706–5743. [Google Scholar] [CrossRef]
- Babu, M.; Snyder, M. Multi-omics profiling for health. Mol. Cell. Proteom. 2023, 22, 100561. [Google Scholar] [CrossRef]
- Ismail, N.H.; Mussa, A.; Al-Khreisat, M.J.; Mohamed Yusoff, S.; Husin, A.; Johan, M.F. Proteomic alteration in the progression of multiple myeloma: A comprehensive review. Diagnostics 2023, 13, 2328. [Google Scholar] [CrossRef]
- Sahu, I.; Zhu, H.; Buhrlage, S.J.; Marto, J.A. Proteomic approaches to study ubiquitinomics. Biochim. Biophys. Acta Gene Regul. Mech. 2023, 1866, 194940. [Google Scholar] [CrossRef]
- Torres, A.; Michea, M.A.; Végvári, Á.; Arce, M.; Morales, A.; Lanyon, E.; Alcota, M.; Fuentes, C.; Vernal, R.; Budini, M.; et al. Proteomic profile of human gingival crevicular fluid reveals specific biological and molecular processes during clinical progression of periodontitis. J. Periodontal. Res. 2023. [Google Scholar] [CrossRef]
- Yadav, R.M.; Marriboina, S.; Zamal, M.Y.; Pandey, J.; Subramanyam, R. High light-induced changes in whole-cell proteomic profile and its correlation with the organization of thylakoid super-complex in cyclic electron transport mutants of Chlamydomonas reinhardtii. Front. Plant Sci. 2023, 14, 1198474. [Google Scholar] [CrossRef] [PubMed]
- Xun, Z.; Yao, X.; Zhu, C.; Ye, Y.; Wu, S.; Chen, T.; Zeng, Y.; Lin, C.; Yang, B.; Ou, Q.; et al. Proteomic characterization of the natural history of chronic HBV infection revealed by tandem mass tag-based quantitative proteomics approach. Mater. Today Bio. 2022, 15, 100302. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, M.; Kokozidou, M.; Gögele, C.; Werner, C.; Auffarth, A.; Kohl, B.; Mrosewski, I.; Schulze-Tanzil, G.G. Does Vitamin K2 Influence the Interplay between Diabetes Mellitus and Intervertebral Disc Degeneration in a Rat Model? Nutrients 2023, 15, 2872. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, H.; Zhang, Z.; Yurube, T.; Takeoka, Y.; Kanda, Y.; Tsujimoto, R.; Miyazaki, K.; Matsuo, T.; Ryu, M.; Kumagai, N.; et al. Anti-inflammatory effects of adiponectin receptor agonist AdipoRon against intervertebral disc degeneration. Int. J. Mol. Sci. 2023, 24, 8566. [Google Scholar] [CrossRef] [PubMed]
- Horvatić, A.; Gelemanović, A.; Pirkić, B.; Smolec, O.; Beer Ljubić, B.; Rubić, I.; Eckersall, P.D.; Mrljak, V.; McLaughlin, M.; Samardžija, M.; et al. Multi-omics approach to elucidate cerebrospinal fluid changes in dogs with intervertebral disc herniation. Int. J. Mol. Sci. 2021, 22, 11678. [Google Scholar] [CrossRef]
- Liao, Z.; Li, S.; Lu, S.; Liu, H.; Li, G.; Ma, L.; Luo, R.; Ke, W.; Wang, B.; Xiang, Q.; et al. Metformin facilitates mesenchymal stem cell-derived extracellular nanovesicles release and optimizes therapeutic efficacy in intervertebral disc degeneration. Biomaterials 2021, 274, 120850. [Google Scholar] [CrossRef]
- Liang, H.; Luo, R.; Li, G.; Zhang, W.; Song, Y.; Yang, C. The proteolysis of ECM in intervertebral disc degeneration. Int. J. Mol. Sci. 2022, 23, 1715. [Google Scholar] [CrossRef]
- Bhujel, B.; Shin, H.E.; Choi, D.J.; Han, I. Mesenchymal stem cell-derived exosomes and intervertebral disc regeneration: Review. Int. J. Mol. Sci. 2022, 23, 7306. [Google Scholar] [CrossRef]
- Wu, Y.; Shen, S.; Shi, Y.; Tian, N.; Zhou, Y.; Zhang, X. Senolytics: Eliminating senescent cells and alleviating intervertebral disc degeneration. Front. Bioeng. Biotechnol. 2022, 10, 823945. [Google Scholar] [CrossRef]
- Dudek, M.; Yang, N.; Ruckshanthi, J.P.; Williams, J.; Borysiewicz, E.; Wang, P.; Adamson, A.; Li, J.; Bateman, J.F.; White, M.R.; et al. The intervertebral disc contains intrinsic circadian clocks that are regulated by age and cytokines and linked to degeneration. Ann. Rheum. Dis. 2017, 76, 576–584. [Google Scholar] [CrossRef]
- Sakai, D.; Nakamura, Y.; Nakai, T.; Mishima, T.; Kato, S.; Grad, S.; Alini, M.; Risbud, M.V.; Chan, D.; Cheah, K.S.; et al. Exhaustion of nucleus pulposus progenitor cells with ageing and degeneration of the intervertebral disc. Nat. Commun. 2012, 3, 1264. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Lei, L.; Li, Z.; Chen, F.; Huang, Y.; Jiang, G.; Guo, X.; Zhao, Z.; Liu, H.; Wang, H.; et al. Grem1 accelerates nucleus pulposus cell apoptosis and intervertebral disc degeneration by inhibiting TGF-β-mediated Smad2/3 phosphorylation. Exp. Mol. Med. 2022, 54, 518–530. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, M.; Derynck, R.; Miyazono, K. TGF-β and the TGF-β family: Context-dependent roles in cell and tissue physiology. Cold Spring Harb. Perspect. Biol. 2016, 8, a021873. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Kokubo, Y.; Nakajima, H.; Honjoh, K.; Watanabe, S.; Matsumine, A. Distribution and polarization of hematogenous macrophages associated with the progression of intervertebral disc degeneration. Spine (Phila Pa 1976) 2022, 47, E149–E158. [Google Scholar] [CrossRef]
- Guo, Z.; Su, W.; Zhou, R.; Zhang, G.; Yang, S.; Wu, X.; Qiu, C.; Cong, W.; Shen, N.; Guo, J.; et al. Exosomal MATN3 of urine-derived stem cells ameliorates intervertebral disc degeneration by antisenescence effects and promotes NPC proliferation and ECM synthesis by activating TGF-β. Oxid. Med. Cell. Longev. 2021, 2021, 5542241. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Li, X.; Jia, X.; Zhu, Z.; Lu, J.; Feng, H.; Shen, B.; Guo, K.; Li, Y.; Wang, Q.; et al. Krüppel like factor 10 prevents intervertebral disc degeneration via TGF-β signaling pathway both in vitro and in vivo. J. Orthop. Translat. 2021, 29, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, C.; Xue, L.; Zhang, F.; Liu, J. Melatonin suppresses apoptosis of nucleus pulposus cells through inhibiting autophagy via the PI3K/Akt Pathway in a high-glucose culture. Biomed. Res. Int. 2021, 2021, 4604258. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, Z.; Hao, J.; Shen, J. Low intensity pulsed ultrasound promotes the extracellular matrix synthesis of degenerative human nucleus pulposus cells through FAK/PI3K/Akt pathway. Spine (Phila Pa 1976) 2016, 41, E248–E254. [Google Scholar] [CrossRef]
- Tan, Y.; Yao, X.; Dai, Z.; Wang, Y.; Lv, G. Bone morphogenetic protein 2 alleviated intervertebral disc degeneration through mediating the degradation of ECM and apoptosis of nucleus pulposus cells via the PI3K/Akt pathway. Int. J. Mol. Med. 2019, 43, 583–592. [Google Scholar] [CrossRef]
- Cheng, C.; Uchiyama, Y.; Hiyama, A.; Gajghate, S.; Shapiro, I.M.; Risbud, M.V. PI3K/AKT regulates aggrecan gene expression by modulating Sox9 expression and activity in nucleus pulposus cells of the intervertebral disc. J. Cell. Physiol. 2009, 221, 668–676. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, F.; Xie, Z.; Chen, L.; Wu, X. The hippo pathway orchestrates cytoskeletal organisation during intervertebral disc degeneration. Acta Histochem. 2021, 123, 151770. [Google Scholar] [CrossRef] [PubMed]
- Shamji, M.F.; Setton, L.A.; Jarvis, W.; So, S.; Chen, J.; Jing, L.; Bullock, R.; Isaacs, R.E.; Brown, C.; Richardson, W.J. Proinflammatory cytokine expression profile in degenerated and herniated human intervertebral disc tissues. Arthritis Rheum. 2010, 62, 1974–1982. [Google Scholar] [PubMed]
- Lerner, A.; Neidhöfer, S.; Reuter, S.; Matthias, T. MMP3 is a reliable marker for disease activity, radiological monitoring, disease outcome predictability, and therapeutic response in rheumatoid arthritis. Best Pract. Res. Clin. Rheumatol. 2018, 32, 550–562. [Google Scholar] [CrossRef] [PubMed]
- Kümper, M.; Zamek, J.; Steinkamp, J.; Pach, E.; Mauch, C.; Zigrino, P. Role of MMP3 and fibroblast-MMP14 in skin homeostasis and repair. Eur. J. Cell Biol. 2022, 101, 151276. [Google Scholar] [CrossRef] [PubMed]
- Aripaka, S.S.; Bech-Azeddine, R.; Jørgensen, L.M.; Mikkelsen, J.D. The expression of metalloproteinases in the lumbar disc correlates strongly with Pfirrmann MRI grades in lumbar spinal fusion patients. Brain Spine 2022, 2, 100872. [Google Scholar] [CrossRef]
- Hu, S.; Fu, Y.; Yan, B.; Shen, Z.; Lan, T. Analysis of key genes and pathways associated with the pathogenesis of intervertebral disc degeneration. J. Orthop. Surg. Res. 2020, 15, 371. [Google Scholar] [CrossRef]
- Hayes, A.J.; Melrose, J. Immunolocalization of keratan sulfate in rat spinal tissues using the keratanase generated BKS-1(+) Neoepitope: Correlation of expression patterns with the class II SLRPs, lumican and keratocan. Cells 2020, 9, 826. [Google Scholar] [CrossRef]
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef]
- Eckfeld, C.; Häußler, D.; Schoeps, B.; Hermann, C.D.; Krüger, A. Functional disparities within the TIMP family in cancer: Hints from molecular divergence. Cancer Metastasis Rev. 2019, 38, 469–481. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, T.; Ma, X.X.; Xiang, H.F.; Hu, Y.G.; Chen, B.H. Lentivirus-mediated TGF-β3, CTGF and TIMP1 gene transduction as a gene therapy for intervertebral disc degeneration in an in vivo rabbit model. Exp. Ther. Med. 2016, 11, 1399–1404. [Google Scholar] [CrossRef]
- Farhan, H.; Hsu, V.W. Cdc42 and cellular polarity: Emerging roles at the golgi. Trends Cell Biol. 2016, 26, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Umbayev, B.; Safarova Yantsen, Y.; Yermekova, A.; Nessipbekova, A.; Syzdykova, A.; Askarova, S. Role of a small GTPase Cdc42 in aging and age-related diseases. Biogerontology 2023, 24, 27–46. [Google Scholar] [CrossRef] [PubMed]
- Pfirrmann, C.W.; Metzdorf, A.; Zanetti, M.; Hodler, J.; Boos, N. Magnetic resonance classification of lumbar intervertebral disc degeneration. Spine (Phila Pa 1976) 2001, 26, 1873–1878. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S.; et al. The STRING database in 2023: Protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2023, 51, D638–D646. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Absorbance 1 | Absorbance 2 | Absorbance 3 | Average Absorbance | Measured Concentration (μg/μL) | True Concentration (μg/μL) |
|---|---|---|---|---|---|---|
| C1 | 0.212 | 0.188 | 0.204 | 0.201 | 0.161 | 1.607 |
| C2 | 0.171 | 0.162 | 0.182 | 0.172 | 0.133 | 1.326 |
| C3 | 0.302 | 0.292 | 0.280 | 0.291 | 0.246 | 2.460 |
| S1 | 0.323 | 0.293 | 0.285 | 0.300 | 0.255 | 2.545 |
| S2 | 0.265 | 0.223 | 0.260 | 0.249 | 0.206 | 2.062 |
| S3 | 0.197 | 0.218 | 0.170 | 0.195 | 0.155 | 1.547 |
| Items | Settings |
|---|---|
| Static modification | TMT (N-term, K); Carbamidomethyl (C) |
| Dynamic modification | Oxidation (M), Acetyl (N-term) |
| Digestion | Trypsin |
| Instrument | Q Exactive HF |
| MS1 tolerance | 10 ppm |
| MS2 tolerance | 0.02 Da |
| Missed cleavages | 2 |
| Database | Uniprot-taxonomy_9606.fasta |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, G.; Li, L.; Yang, Z.; Zhang, C.; Kang, X. TMT-Based Proteomics Analysis of Senescent Nucleus Pulposus from Patients with Intervertebral Disc Degeneration. Int. J. Mol. Sci. 2023, 24, 13236. https://doi.org/10.3390/ijms241713236
Zhang G, Li L, Yang Z, Zhang C, Kang X. TMT-Based Proteomics Analysis of Senescent Nucleus Pulposus from Patients with Intervertebral Disc Degeneration. International Journal of Molecular Sciences. 2023; 24(17):13236. https://doi.org/10.3390/ijms241713236
Chicago/Turabian StyleZhang, Guangzhi, Lei Li, Zhili Yang, Cangyu Zhang, and Xuewen Kang. 2023. "TMT-Based Proteomics Analysis of Senescent Nucleus Pulposus from Patients with Intervertebral Disc Degeneration" International Journal of Molecular Sciences 24, no. 17: 13236. https://doi.org/10.3390/ijms241713236
APA StyleZhang, G., Li, L., Yang, Z., Zhang, C., & Kang, X. (2023). TMT-Based Proteomics Analysis of Senescent Nucleus Pulposus from Patients with Intervertebral Disc Degeneration. International Journal of Molecular Sciences, 24(17), 13236. https://doi.org/10.3390/ijms241713236