Abstract
There is an urgent unmet need for robust and reliable biomarkers for early diagnosis, prognosis, and prediction of response to specific treatments of many aggressive and deadly cancers, such as pancreatic cancer, and liquid biopsy-based miRNA profiling has the potential for this. MiRNAs are a subset of non-coding RNAs that regulate the expression of a multitude of genes post-transcriptionally and thus are potential diagnostic, prognostic, and predictive biomarkers and have also emerged as potential therapeutics. Because miRNAs are involved in the post-transcriptional regulation of their target mRNAs via repressing gene expression, defects in miRNA biogenesis pathway and miRNA expression perturb the expression of a multitude of oncogenic or tumor-suppressive genes that are involved in the pathogenesis of various cancers. As such, numerous miRNAs have been identified to be downregulated or upregulated in many cancers, functioning as either oncomes or oncosuppressor miRs. Moreover, dysregulation of miRNA biogenesis pathways can also change miRNA expression and function in cancer. Profiling of dysregulated miRNAs in pancreatic cancer has been shown to correlate with disease diagnosis, indicate optimal treatment options and predict response to a specific therapy. Specific miRNA signatures can track the stages of pancreatic cancer and hold potential as diagnostic, prognostic, and predictive markers, as well as therapeutics such as miRNA mimics and miRNA inhibitors (antagomirs). Furthermore, identified specific miRNAs and genes they regulate in pancreatic cancer along with downstream pathways can be used as potential therapeutic targets. However, a limited understanding and validation of the specific roles of miRNAs, lack of tissue specificity, methodological, technical, or analytical reproducibility, harmonization of miRNA isolation and quantification methods, the use of standard operating procedures, and the availability of automated and standardized assays to improve reproducibility between independent studies limit bench-to-bedside translation of the miRNA biomarkers for clinical applications. Here I review recent findings on miRNAs in pancreatic cancer pathogenesis and their potential as diagnostic, prognostic, and predictive markers.
1. Introduction
Pancreatic ductal adenocarcinoma (PDAC) is the most common and highly aggressive and deadly pancreatic cancer characterized by an unparalleled mortality-to-incidence ratio and with a poor prognosis, and the number of patients with this disease is increasing [1,2,3]. This is partly due to the rapid course of disease and absence of symptoms at the initial stage of the disease as well as late detection and lack of effective biomarkers for monitoring its progress. The early and aggressive local invasion and high rate of metastasis are the main characteristics of pancreatic cancer with the worst rates of overall survival. The five-year survival rate has only just reached ~11% [4], and the standard of therapy improves survival in a range of months. In addition, approximately 50% of newly diagnosed pancreatic cancers are metastatic, with an average survival of patients less than one year [2]. It is estimated that 64,050 individuals will be diagnosed and 50,550 individuals will die from PDAC in the United States [5]. Due to the rising incidence of disease and lack of or limited effectiveness of treatments, pancreatic cancer is projected to become the second-leading cause of cancer-related deaths by 2030 [3,5]. Because surgery is considered to be the only curative treatment modality for pancreatic cancer, early detection has the potential to significantly improve outcomes [5].
Such stark statistics are driven are driven by the stealthy onset of the disease that often features non-specific symptoms and the absence of effective biomarkers for screening; the majority of pancreatic cancer cases (82%) are detected too late, and most diagnoses are made after the opportunity for surgical intervention has passed, resulting in surgically unresectable, locally advanced or metastatic cancer [5]. Even after complete surgical resection, 80% of patients experience recurrence, and at this stage the disease is almost universally deadly [6,7,8]. On the other hand, in the metastatic setting, the adjuvant standard of care first-line chemotherapy treatment options [i.e., FOLFIRINOX (5-fluorouracil, leucovorin, irinotecan, and oxaliplatin) and the combination of gemcitabine and nab-paclitaxel] have been shown to benefit and reduce recurrence risk (overall survival benefit of 19.4 months over gemcitabine alone, 54.4 months vs. 35.0 months) [5,7]. It has been shown that FOLFIRINOX improves the median overall survival by 4.3 months (11.1 months vs. 6.8 months) as compared to gemcitabine monotherapy [7,9]. Likewise, the combination of nab-paclitaxel with gemcitabine was shown to improve median overall survival by 1.8 months (8.5 months vs. 6.7 months) [10].
Although screening high-risk individuals (e.g., those with predisposing genetic conditions with a lifetime pediatric cancer risk of >5–15%, high-risk pancreatic lesions, and strong family history, obesity, and type 2 diabetes) [2] with magnetic resonance imaging (MRI) or endoscopic ultrasound (EUS) [11] is recommended, the optimal timing of screening and the benefit of the combination of these tests remains to be determined. Likewise, screening tests to ascertain response to treatment and disease recurrence are also limited to MRI imaging and infrequently tumor markers measured in serum that suffer from poor sensitivity and specificity [12,13]. Importantly, while screening high-risk individuals might have some benefits, it has been shown that universal screening of the general public is ineffective [11].
Because of this, there is an urgent need for the discovery and development of easily detectable biomarkers as diagnostic tools to detect disease at earlier stages, guide therapy selection, monitor treatment response, and predict the recurrence of the disease and hence improve the survival rates of patients [14].
Cancer is a complex and heterogeneous disease that evolves through successive genetic and genomic as well as epigenetic changes that support tumorigenesis. It is a disease that is driven by both internal stimuli such as genome alterations leading to germline and somatic alterations and external stimuli that play a role in the introduction of some genome alterations and expression patterns of certain genes and cellular signaling pathways associated with cell growth, migration, and metastasis. Changes in the genome that affect gene function often result from genomic alterations including chromosomal translocations, insertions or deletions, amplifications, and single-nucleotide mutations or from the epigenome leading to activation of oncogenes and suppression of tumor suppressor genes [15]. As described in a recent review [16], the hallmarks of cancer make it even more challenging to tackle this complex disease.
Although the majority of oncology research is still focused on the dynamic variations of proteins and protein-coding RNAs (their corresponding coding sequences only account for ∼2% of the genome (https://www.genomicseducation.hee.nhs.uk/genotes/knowledge-hub/non-coding-dna/ (accessed on 26 July 2023)) [17,18,19]), the role of non-coding RNAs (ncRNAs) transcribed from the remaining 98% of the genome including microRNAs (miRNAs) play key roles in a multitude of biological processes in normal physiological states [20] but also in the development of various types of diseases including cancer [20], underscoring the importance of miRNAs and other ncRNAs in tumor initiation and progression. As such, miRNAs and their altered expression have been recognized as an additional molecular mechanism responsible for the pathological processes of many diseases [21,22,23] including innate immunity [24], autoimmunity and autoimmune diseases [25], viral infections [26,27,28,29], acute hepatitis [30], depression [31], anxiety [32], Alzheimer’s disease [33], Huntington’s disease [34], obesity, metabolic and cardiovascular diseases [35,36,37,38], diabetes [39,40,41,42,43,44], and many cancers [5,14,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73], consequently, these miRNAs can be used to indicate the presence of a pathology and even the stage, progression, or genetic link of the disease. Supporting this, recent findings on miRNAs highlight their roles during tumor pathogenesis as well as in response to various therapies.
Although genetic, genomic, and epigenetic changes affect genome function, dysfunctions of various types of regulators are the primary molecular mechanisms responsible for the pathological processes of many cancers, miRNAs and their altered expression and members of the miRNA biogenesis pathway have also been recognized as additional molecular mechanisms implicated in the pathological processes of many cancers and have received great attention in recent years. To support this, new data on the molecular mechanisms responsible for the dysregulation of miRNA biogenesis and expression in cancer are continuously emerging. For example, it has been shown that genetic deletion or amplification, epigenetic methylation of miRNA genomic loci, and alterations affecting the transcription factor-mediated regulation of primary miRNA (pri-miRNA) as well as components involved in the miRNA biogenesis pathway frequently alter miRNA expression and function in many cancers. In addition, recent data indicate that other factors including oncogenic drivers such as mutations in the KRAS gene can also affect global miRNA biogenesis and effector function, leading to global miRNA dysregulation [74]. Because of this, miRNAs and their dysregulation have gained attention both from academia and industry as a research discipline both for understanding disease biology [59,75,76,77] and to develop them as potential diagnostic, prognostic, and predictive biomarkers as well as drug targets or as therapeutics [47,48,51,59,67,78].
Here, I review how miRNA expression is dysregulated in cancer with a focus on pancreatic cancer and discuss the clinical applications of miRNAs as potential biomarkers that may be used to facilitate patient diagnosis, prognosis, monitoring, and treatment in the oncology field.
2. microRNAs
After the discovery of the first microRNA (miRNA), lin-4, in 1993 in Caenorhabditis elegans [79,80], it was quickly recognized that microRNAs have been detected throughout the animal and plant kingdoms and some were shown to be highly conserved across species [81,82,83], with a conserved mechanism and broad functional significance.
MiRNAs are small non-coding RNAs, with an average of 22 nucleotides in length which are highly conserved and are naturally encoded in the genomes of various species [81,82,83,84] and play key functional roles in the regulation of gene expression at the transcriptional [85,86,87] and post-transcriptional [41,88,89,90,91] levels of their target mRNAs [41,89] including the regulation of cell function by regulating gene expression via the modulation of the stability and translation of mRNA [92] in a broad range of biological processes within cells and organisms, consequently affecting cell differentiation, cell proliferation, angiogenesis, and apoptosis, etc. [48]. In addition, miRNAs exhibit tissue-specific [67,93] and developmental expression patterns [75,76,77].
Emerging data suggest that new miRNAs are still being discovered [94] and curreently, it is estimated that there are >2588 human mature miRNAs in human cells with time- and tissue-dependent expression patterns. Of the >2588 miRNAs, 1115 are currently annotated in miRBase V22 [15,16,21,95,96,97,98]. It is estimated that these >2588 miRNAs regulate over 60% of the expression of human genes, demonstrating that they have important regulatory roles in diverse physiological and developmental processes as well as in the pathogenesis of various human diseases and disorders [21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44], including many cancers [5,14,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73].
As illustrated in Figure 1, most miRNAs are transcribed from DNA sequences into nascent primary miRNAs (pri-miRNAs) which are then initially processed by DROSHA in the nucleus to generate precursor miRNA (pre-miRNA) [41,99]. As many as 40% of miRNA genes may lie in the introns or even exons of other genes [100]. Pre-miRNAs are subsequently exported from the nucleus to the cytoplasm by exportin 5 (XPO5) where they are further processed by DICER, producing small RNA duplexes with 2 nt. 3′ overhangs. These small double-stranded RNA duplexes with distinct 2 nucleotides 3′ overhangs are then loaded onto the Argonaute (AGO) protein, which retains only one strand of mature miRNA by removing the other strand [89]. The complex formed by the AGO and miRNA associates with cofactors such as GW182 (i.e., TNRC6A), and forms the effector complex, the RNA-induced silencing complex (RISC) [91]. The miRNA-RISC complex is responsible for the degradation of mRNA transcripts and translational suppression through interaction with the complementary mRNA target sequences predominantly located within the 3′-untranslated region (3′-UTR) of mRNAs (Figure 1) [101,102,103,104].
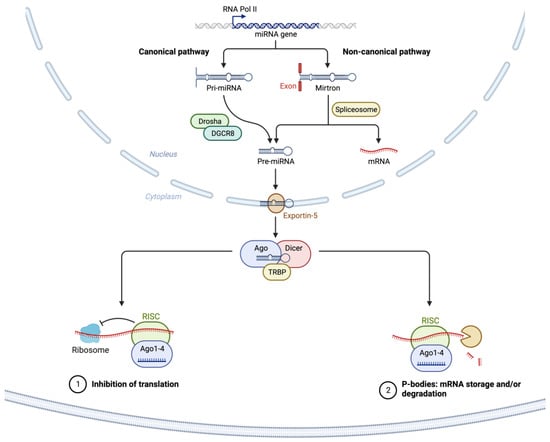
Figure 1.
Schematic of canonical and noncanonical miRNA biogenesis, processing, and target RNA translational suppression or degradation. The canonical miRNA pathway generates pri-miRNAs from transcripts by miRNA genes encoded in intronic, exonic, or intergenic regions. The pri-miRNA transcripts are then processed by Drosha/DGCR8 into pre-miRNAs. Intronic pre-miRNAs of the noncanonical mirtron pathway are formed by splicing, debranching, and trimming short introns without the involvement of Drosha processing. The pre-miRNA transcripts produced by both the canonical and noncanonical pathways are then exported from the nucleus to the cytoplasm by Exportin 5. This is followed by subsequent Dicer cleavage within the RISC loading complex (RLC) and subsequent unwinding of the miRNA/miRNA duplex via Argonaute, and TRBP-dependent loading into the RNA-induced silencing complex (RISC). The binding of target mRNAs to miRNAs in RISC is followed by suppression of translation and/or mRNA degradation within p-bodies in the cytosol. Created with BioRender.com (accessed on 6 July 2023). Abbreviations: AGO2: Argonaute RISC Catalytic Component 2; DICER1: Dicer 1, Ribonuclease III; EGFR: Epidermal growth factor receptor 2; DGCR8: DiGeorge Syndrome Critical Region 8; DROSHA: Drosha Ribonuclease III; mRNA: messenger RNA; miRNA: micro RNA; P-bodies: Processing bodies; pre-miRNA: precursor microRNA; pri-miRNA: primary microRNA; RNA Pol II: RNA polymerase II; TRBP: AR RNA-binding protein; XPO5: Exportin 5.
As discussed in the literature [59], a specific miRNA may target many different mRNAs [105]; while a particular messenger RNA may be targeted by several miRNAs, either simultaneously or in a context-dependent fashion [106], resulting in the cooperative repression effect [107,108].
3. miRNA Dysregulation in Cancer
Cancer is a complex and heterogeneous disease that evolves through successive genetic and genomic changes that support tumorigenesis [109]. Changes in the genome that affect gene function often result from genomic alterations such as chromosomal translocations, insertions or deletions, amplifications, and single-nucleotide mutations in the epigenome. These genetic and epigenetic alterations often trigger the activation of oncogenes and suppression of tumor suppressor genes [110]. To add to this picture, miRNAs also play an important role in organismal development, normal physiology, and disease state, including many hallmarks of various cancer types (Figure 2).
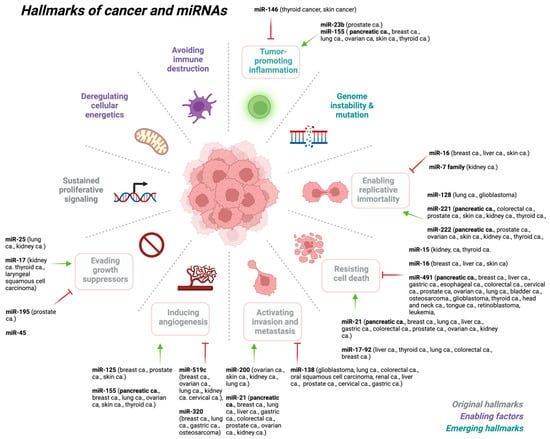
Figure 2.
miRNAs play key roles at various stages of cancer development. Created with BioRender.com (accessed on 6 July 2023). Abbreviations: ca: cancer.
Although genetic, genomic, and epigenetic changes affecting genome function as well as the dysfunctions of various types of regulators are the primary molecular mechanisms responsible for the pathological processes of a multitude of human diseases, including many cancers, among which, miRNAs and their altered expression, and alterations and mutations in members of miRNA biogenesis pathways have also been recognized as additional molecular mechanisms implicated in the pathological processes of many cancers and have received great attention in recent years. Numerous studies have demonstrated the involvement of miRNAs in cancer pathogenesis by regulating the expression of their target mRNAs contributing to tumor growth, invasion, angiogenesis, and immune system evasion [111,112]. Furthermore, studies have shown that tumor-specific miRNA signatures can define cancer subtypes, patient survival, and treatment response [46,113,114], and notably, cancer-associated miRNAs can be detected in biological fluids, allowing less-invasive monitoring [115].
It is widely accepted that genetic deletion or amplification, epigenetic methylation of miRNA genomic loci, and alterations affecting the transcription factor-mediated regulation of primary miRNA (pri-miRNA) as well as factors involved in the miRNA biogenesis pathway frequently alter miRNA expression and function in many cancers.
Alterations throughout miRNA biogenesis can affect the availability of target mRNA, including many mRNAs associated with cancer development (Figure 3). As such, altered or dysregulated miRNAs or miRNA-processing machinery have been linked to many pathological processes, resulting in the failure to maintain the normal homeostatic state and eventually leading to malignant transformation, including many cancers [5,14,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73].
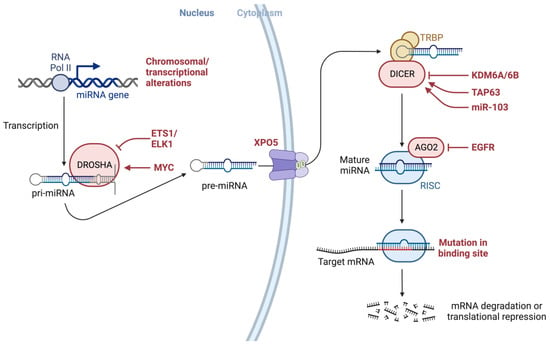
Figure 3.
Alterations throughout miRNA biogenesis can affect the availability of target mRNA, including many mRNAs associated with cancer development. The figure also depicts several modulators, proteins such as EGFR, and transcription factors such as ETS1/ELK1, MYC, epigenetic regulators such as demethylating protein KDM6A, a known tumor suppressor, the tumor suppressor gene TAP63, and miRNAs such as miR-103 that may interact with modulators of miRNA-processing machinery. Both KRAS and EGFR are essential mediators of pancreatic cancer development and interact with AGO2 to perturb its function. Created with BioRender.com (accessed on 6 July 2023). Abbreviations: AGO2: Argonaute RISC Catalytic Component 2; DICER1: Dicer 1, Ribonuclease III; DROSHA: Drosha Ribonuclease III; EGFR: Epidermal growth factor receptor 2; EGFR: Epidermal growth factor receptor, ELK1: ETS Like-1 protein Elk-1; ETS1: ETS Proto-Oncogene 1; KDM6A: Lysine (K)-specific demethylase 6A; miRNA: micro RNA; mRNA: messenger RNA; MYC: MYC Proto-Oncogene; pre-miRNA: precursor microRNA; pri-miRNA: primary microRNA; RNA Pol II: RNA polymerase II; TAP63: Tumor protein p63; XPO5: Exportin 5.
As illustrated in Figure 3, various genetic or epigenetic alterations in key factors involving the miRNA biogenesis process can affect the availability of miRNA targets, including many targets associated with cancer development, as well as several modulators and proteins (e.g., EGFR), transcription factors (e.g., ETS1/ELK1, MYC), epigenetic regulators (e.g., demethylating protein KDM6A, a known tumor suppressor), the tumor suppressor gene TAP63, and miRNAs (e.g., miR-103) [116] that may interact with factors involved in miRNA biogenesis machinery. It has been shown that both KRAS and EGFR are essential mediators of pancreatic cancer development and they were shown to interact with Argonaute 2 (AGO2) to perturb its function [117].
Recent findings indicate that different types of miRNAs have been shown to regulate many of the hallmarks of cancer Figure 2 [71], either as a tumor suppressor (i.e., oncosuppressor miRs) or as oncogenes (i.e., oncomirs) [118,119,120,121,122,123,124,125,126,127,128]. MiRNAs play a critical role in the regulation of the expression of a multitude of genes, such as the cellular responses to environmental stresses including DNA damage, hypoxia, oxidative stress, and starvation.
Supporting this, miRNAs with an oncogenic function as well as tumor-suppressing function have recently been identified in various neoplastic malignancies, and dysregulation of miRNA expression is closely associated with cancer initiation, progression, and metastasis and shown to be associated with the origin, progression, therapeutic response, and patient survival of the disease [47,51,55,62,67,72,73,129,130].
For example, the tissue specificity of miRNAs [67,93], which is required for maintaining normal cell and tissue homeostasis [46], allows them to be used as potential biomarkers in diagnosing cancer of unknown primary origin [97,131].
Supporting these findings, recurrent genetic and epigenetic alterations of individual miRNAs and components involved in the miRNA-processing machinery and biogenesis pathway have been identified in many cancer types, and emerging data from several functional studies provide evidence for the mechanism of action of many potential oncogenic and tumor-suppressor miRNAs [118,119,120,121,122,123,124,125,126,127,128] with potential clinical applications as potential diagnostics and therapeutics.
4. Oncogenic Mutations in Key Factors Involved in the miRNA Biogenesis
As discussed in the literature [74], oncogenic mutations in factors involved in miRNA biosynthesis, processing, and export machinery are associated with many cancers. The majority of these oncogenic mutations in the microprocessing complex and export machinery result in a decrease in miRNA biosynthesis, leading to global dysregulation of miRNAs and the mRNA transcripts they regulate.
For example, mutations (e.g., E518K, rs417309 G/A, p.R32fs, copy-number loss, p.S92fsa) in DGCR8, a member of the nuclear microprocessing complex which binds pri-miRNAs to facilitate processing by DROSHA has been linked to various human cancers including Wilms tumor, thyroid carcinoma, FMGS, laryngeal cancer, breast cancer, and pineoblastoma [132,133,134,135,136].
Similarly, mutations (rs640831 C/A, rs1110386 G/Aa, rs486732 C/Aa, p.E500, p.R277C, p.Q136a) in RNASEN/DROSHA, a member of the nuclear microprocessing complex and a ribonuclease that cleaves pir-miRNA to pre-miRNAs have been linked to several human cancers including lung adenocarcinoma, ovarian cancer, bladder cancer, breast cancer, thyroid carcinoma, and pineoblastoma [136,137,138,139,140,141].
Mutations (p.S625C, p.P89A) in DHX9, a member of the nuclear microprocessing complex and a helicase that modulates nuclear miRNA processing have been linked to breast cancer [142].
Mutations (p.T1182del, rs2257082 A/G) in XPO5, which facilitates pre-miRNA export from the nucleus to the cytoplasm, have been linked to colon cancer, gastric cancer, endometrial cancer, and gastric cancer [143,144].
Likewise, mutations (germline: p.Y1180, p.Y819H, p.D1713Aa, p.R187a, p.G803Ra, p.E503Xa and somatic: p.A872Ta, p.E428Ka, p.E813Qa, p.D1709Na, p.D1709G, p.D1810Y, p.E1813Qa, p.Y1701) in DICER1, a member of the RISC complex, and an endoribonuclease that cleaves the stem-loop from pre-miRNAs, leading to the formation of mature miRNAs, have been linked to fetal lung adenocarcinoma, Sertoli–Leydig ovarian tumor, FMGS, hepatocellular carcinoma, Wilms tumor, pleuropulmonary blastoma, embryonal ovarian cancer, yolk-sac tumor, juvenile granulosa-cell tumor, and teratoma, pineoblastoma [136,145,146,147,148,149,150,151,152].
Mutations (p.P144fs, p.R353fs, p.M145fs, copy-number gain) in TARBP1, a member of the RISC complex and a binding partner of DICER that facilitates RISC assembly, have been linked to colon cancer, Wilms tumor, gastric cancer, and adrenal cancer [63,153,154,155].
Mutations (p.P127L) in PRKRA, a member of the RISC complex and a binding partner of DICER that facilitates RISC assembly, have been linked to ovarian cancer [63,153,154,155].
Mutations (copy-number gain) in AGO2, a member of the RISC complex and an endonuclease that has mRNA cleavage function, have been linked to head and neck cancer, multiple myeloma, breast cancer, and ovarian cancer [156,157,158].
Mutations (p.P115-Q118del, p.R1183fs, p.Trp804fs) in TNRC6A, a member of the RISC complex and a binding partner of AGO2 required for mRNA silencing, have been linked to esophageal cancer, gastric cancer, and colorectal cancer [63,159,160].
As an additional mechanism that promotes malignant transformation, recent data [74] show that mutant KRAS is also involved in the modulation of the activity of many members of miRNA processing and regulatory pathways. Supporting this, previous studies have shown that miRNA biogenesis and global miRNA expression levels are significantly dysregulated in cancers harboring KRAS mutations [161,162,163,164].
The gene encoding KRAS is the most mutated oncogene in many cancers including pancreatic ductal adenocarcinoma (PDAC), non-small-cell lung cancer (NSCLC), and colorectal cancer (CRC) [165,166,167,168]. Activating mutations in the KRAS gene result in the constitutive activation of downstream signaling cascades, consequently leading to constitutive and sustained proliferation, self-renewal, and increased vascularization [166,169,170,171,172]. Notably, the functional effects of KRAS mutations correlate with the oncogenic effects of global miRNA dysregulation [173].
Similar to the downstream effects of mutations in the key factors involved in miRNA biogenesis and processing pathways, oncogenic KRAS affects global miRNA dysregulation. This is achieved by directly regulating the activity of key factors involved in miRNA biogenesis, processing, and regulatory pathways by oncogenic KRAS as an additional mechanism to promote malignant transformation. Thus, this regulatory mechanism disrupted by oncogenic KRAS may represent a novel vulnerability in KRAS-dependent cancers such as pancreatic cancer and others.
Furthermore, data show that mutations in the factors involved in the miRNA biogenesis, processing, and export machinery have been linked to a decrease in miRNA biosynthesis, thus leading to the global dysregulation of miRNAs and the mRNA transcripts they target [74].
Beyond the effect of mutant KRAS on miRNA biogenesis and processing pathways, mutant KRAS also alters how miRNAs regulate their target RNA [74]. For instance, the mutant KRAS/AGO2 interaction was shown to dysregulate the effector function of miRNAs, as AGO2 and the miRNA/RISC are essential for miRNA localization and silencing of miRNA targets. Supporting this, cancer cells with KRAS mutations have been shown to have increased levels of AGO2 phosphorylation at serine 387 (AGO2S387) [174].
Beyond directly regulating members of the miRNA regulatory pathway, mutant KRAS was also shown to alter the coordination and the storage of miRNA-targeted transcripts in intracellular condensates involved in transcript regulation, such as stress granules (SGs) and processing bodies (PBs) [175,176,177,178].
Thus, a better understanding of the global miRNA dysregulation induced by oncogenic KRAS may reveal novel targetable pathways for therapeutic intervention.
Recently, two new drugs, sotorasib (Lumakras) [179] and adagrasib (Krazati) [180], have been approved to treat people with non-small cell lung cancer that has the KRAS G12C mutation, and those in development include MRTX1133 targeting KRAS G12D mutations. As such, concurrent targeting of various forms of mutant KRAS and individual members of the miRNA biogenesis and processing machinery might represent a synergistic therapeutic strategy to treat several types of cancer with KRAS mutation [181].
5. Liquid Biopsy in Pancreatic Cancer
Because pancreatic cancer is a highly aggressive and one of the most lethal types of cancer with a five-year survival rate remaining low (~11%), it is always diagnosed at an advanced stage and is resistant to therapy. Thus, there is an urgent unmet need for a reliable and robust biomarker that can detect asymptomatic premalignant or early malignant tumors and predict the response to treatment for pancreatic cancer patients [5].
Although used extensively for early diagnosis of cancers, carcinoembryonic antigen (CEA) [13] and carbohydrate antigen 19-9 (CA19-9) [13,182] have limitations in clinical practice and a single biopsy of tumor specimens is often not informative enough about tumor progression and metastasis [183]. As such, there is an unmet need for biomarkers that can detect disease in early stages and also predict the response to treatment and recurrence of tumors.
Because of this, it is fundamentally important for the detection of early asymptomatic disease, such that treatments can begin before the disease becomes unmanageable and metastatic and this can be only possible with the help of biomarkers that can be easily measured in real-time and repeatedly sampled to help diagnose disease in early stages and also help with the prediction of response to a specific treatment.
The liquid biopsy in pancreatic cancer, as discussed in detail in the literature [5,184] and illustrated in Figure 4, is a non-invasive or minimally invasive process followed by miRNA profiling that can inform the miRNA signatures for each type of cancer as well as stages of disease.
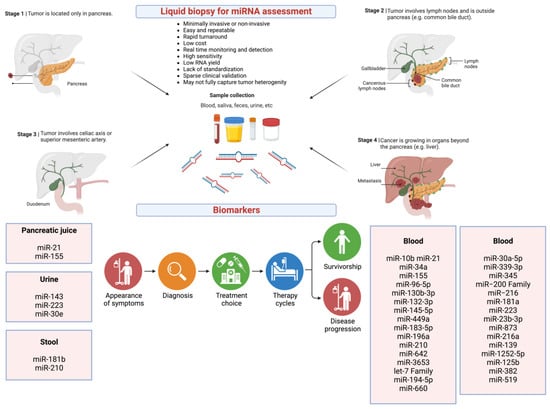
Figure 4.
miRNAs as potential liquid biopsy biomarkers for pancreatic cancer. Created with BioRender.com (accessed on 6 July 2023). Abbreviations: Let-7 family: lethal-7 family of 12 miRNAs; miRNA: micro RNA; miR: micro RNA.
Unlike other methods such as invasive biopsy, liquid biopsy is a minimally invasive or non-invasive procedure depending on the bodily fluid collected (e.g., blood vs. saliva) for biomarker assessment. It gives a real-time and longitudinal assessment of the disease course and enables monitoring of treatment response. Importantly, liquid biopsy can be used to profile circulating miRNAs as biomarkers in real-time and longitudinally, which can inform the miRNA signatures for each type of cancer as well as stages of disease.
For example, as illustrated in Figure 4, in recent years, many circulating miRNAs identified as potential diagnostic and prognostic biomarkers in pancreatic cancer have been reported in the literature [66,185,186,187,188,189].
6. miRNAs as Potential Biomarkers
As discussed in the literature [190], dysregulated expression of miRNAs has been implicated in various human diseases and disorders [21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44], including many cancers [5,14,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73], these miRNAs have been investigated in several clinical trials as indicators of the presence of a pathology and even of the stage, progression, or genetic link of disease. Because of this, miRNAs, especially those in circulation have been exploited as novel, minimally invasive, or non-invasive biomarkers because of their relatively high stability in circulation and other biofluids, sensitivity, and ease of detection using various detection methods in various biological fluids.
Notably, in some situations, one miRNA biomarker may be sufficient to identify a health outcome; however, in many other cases, a panel of well-defined and validated miRNAs has been used for increased diagnostic sensitivity and specificity as biomarkers [190].
Recently, many miRNAs were detected in cell-free conditions in circulation, including biofluids such as whole blood, plasma, serum, saliva, tears, urine, stool, pancreatic juice, breast milk, colostrum, seminal fluid, amniotic fluid, bronchial lavage, cerebrospinal fluid, peritoneal and pleural fluids, and other body-fluid types [191] and they exhibit specific expression patterns that are associated with altered physiological conditions and disease states [37,38,41,42,43,44,51,192,193,194].
miRNAs can be secreted into circulation in extracellular fluids and shuttled to target cells via extracellular vesicles, including exosomes, microvesicles, and apoptotic bodies under various physiologic and pathologic conditions, enabling them to function as chemical messengers to mediate cell–cell communication [193,195,196], or by binding to proteins such as Argonautes (AGO), especially AGO2 [195,197]. In addition, secreted miRNAs trafficked between different subcellular compartments have been shown to regulate translation, and even transcription [198].
The source of miRNAs, how accessible it is as a biomarker, how the miRNA is extracted from the biological specimen, and even how it is analyzed can significantly influence the miRNA analysis results. In biofluids [191], the miRNA content can originate through a few pathways, including extracellular vesicles (exosomes) [199,200,201,202]. Studies are investigating whether the enrichment of exosomes from biofluids or direct cell-free preparations are more reliable in miRNA isolation and measurements and some studies indicate that it may depend on the specific human disease [190].
It is important to note that exosomal vesicle purification can lower the RNA yield and integrity which can lead to more interindividual variability. Furthermore, a disease-specific alteration in exosome release and clearance, or administration of therapeutics can alter blood volume and can skew RNA yield and signature [203].
In addition to being contained within exosomes, significant portions of miRNAs appear to be contained in various cell types such as tumor cells [204], stem cells [205], macrophages [206], and adipocytes [207], all of which release exosomes into circulation with specific miRNA (exomiRs) content.
7. Circulating miRNAs Detected in Different Body Fluids
An ideal biomarker should be accessible via non-invasive techniques, be reasonably sensitive to distinguish early tumor presence before clinical symptoms and should be undetectable or low in healthy people [116,208].
Their non-invasive or minimally invasive accessibility, relative stability, low complexity, and ease of detection via various techniques make circulating miRNAs ideal candidates as biomarkers for several diseases [116].
Although miRNA profiling of tumor tissue has been used as a prognostic test, tumor sampling requires invasive techniques such as biopsy, aspiration biopsy, or excision surgery of prevailing tumors [208,209].
Data indicate that miRNAs can be detected in various body fluids including whole blood, plasma, serum, saliva, tears, urine, stool, pancreatic juice, breast milk, colostrum, seminal fluid, amniotic fluid, bronchial lavage, cerebrospinal fluid, peritoneal and pleural fluids, and others and that these miRNAs relate to different pathophysiological conditions [191].
It is reasonable to argue that circulating miRNAs detected in the above body fluids could be originating from lysed or dying cells [210]; however, data indicate that miRNAs are actively exported from the cells within exosomes [211] or as exosome- or microvesicle-free miRNAs attached to Argonaute2 complexes [212].
Although circulating miRNAs from plasma and serum are the most common method for profiling circulating miRNAs as biomarkers, other biofluids such as urine and saliva are also used as the source for circulating miRNAs [130].
As discussed in the literature [130], a recent report has analyzed miRNA expression data from 40 human healthy tissues and those from human body fluids, including serum, plasma, urine, bile, and feces, and shown a positive correlation between body fluid miRNAs and tissue miRNAs [213].
For example, miR-186-5p was shown to be overexpressed not only in tumor tissues and blood but also in the urine of bladder cancer patients [214].
Similarly, several miRNAs, including miR-210-3p, were shown to be elevated in the urine of patients with transitional cell carcinoma [215].
Another example is members of the let-7 family miRNAs. Data show that all members of let-7 families of miRNAs were significantly upregulated in urine from clear-cell renal cell cancer (ccRCC) patients [216] and a combination of urinary miRNAs including miR-34b-5p, miR-126-3p, and miR-449a-3p could be used for ccRCC diagnosis [217].
A panel of four miRNAs including miR-21-5p, miR-125b-5p, miR-155-5p, and miR-451-5p was found to be elevated in urine samples of breast cancer patients and shown their diagnostic utility [218].
miR-143-3p and miR-30e-5p were found to be elevated in urine samples of pancreatic ductal adenocarcinoma (PDAC) patients and were shown their potential utility as diagnostic biomarkers in the early stage of the disease [219].
Collectively, circulating miRNAs can be found in various bodily fluids and be obtained with minimally invasive or non-invasive methods, and potentially be used as biomarkers for human diseases including cancer.
In addition, miRNAs are relatively stable in those different types of body fluids making them ideal biomarkers for early diagnosis, prognosis, prediction, and monitoring of various types of cancers.
8. Circulating miRNAs as Potential Biomarkers in Pancreatic Cancer
Concerning their utility as potential biomarkers in pancreatic cancer, several studies have reported that the expression of several circulating miRNAs is altered in pancreatic cancer [14,62,66,67,129,130,185,186,187,188,189,220,221,222,223,224].
Table 1 illustrates examples of recent clinical findings on circulating miRNAs as prognostic markers and possible mechanisms of action and survival status in pancreatic cancer that have been reported in the literature [5,14,62,67,129,225,226].

Table 1.
Recent clinical findings on miRNAs as prognostic markers and possible mechanisms of action and survival status in pancreatic cancer. Abbreviations: Akt: Ak strain transforming; Anxa3: Annexin A3; ATRX: Alpha-thalassemia/mental retardation, X-linked; BCL2: B-cell lymphoma 2; CDK6: Cell division protein kinase 6; E-cadherin: Epithelial cadherin; KRAS: Kirsten rat sarcoma virus; FOXO1: Forkhead box protein O1; HIV-1 Tat interactive protein 2 (HTATIP2); HRAS: Harvey rat sarcoma virus; IL-6R: Interleukin 6 receptor; TIP30: JAK: Janus kinase; miR: microRNA; mRNA: messenger RNA; NA: not applicable; NEDD9: Neural precursor cell expressed developmentally down-regulated protein 9; NOTCH: Neurogenic locus notch homolog protein; PD-L1: Programmed death-ligand 1; PDCD4: Programmed cell death protein 4; PI3K: Phosphoinositide 3-kinase; PLEK2: Pleckstrin 2; PTEN: Phosphatase and tensin homolog; RalB: RAS like proto-oncogene B; RKIP: RAF-kinase inhibitor protein; ROCK1: Rho-associated coiled-coil kinases; TP53INP: Tumor protein p53-inducible nuclear protein; TRIM71: Tripartite motif containing 71: WT1: Wilms’ tumor suppressor gene1; ZEB1: Zinc finger E-box-binding homeobox 1; ZIC1: Zinc finger of the cerebellum.
Examples include miR-10b, which was shown to be upregulated in pancreatic cancer and shown to silence TIP30 and is linked to poor survival [227,228].
miR-21 which was shown to be upregulated in pancreatic cancer and silences PTEN, PDCD4, IL-6R, and CDK6 mRNAs is associated with worse survival [229,230,231].
On the other hand, miR-34a which was shown to be upregulated in pancreatic cancer and silences NOTCH, BCL2, and CDK6 mRNAs, is associated with better survival [209,234,235].
Likewise, the miR−200 family, which was shown to be downregulated in pancreatic cancer and inhibits E-cadherin, ZEBR, is associated with better survival [234,246,247].
Similarly, miR-873, which was shown to be downregulated in pancreatic cancer and inhibits PLEK2 mRNA via the pleckstrin-2-dependent PI3K/AKT pathway, is associated with better survival [253].
The let-7 family, which was shown to be downregulated in pancreatic cancer and inhibits KRAS, HRAS, and TRIM71, is associated with poor survival [243,244].
miR−216, which was shown to be downregulated in pancreatic cancer and inhibits ROCK1, is associated with poor survival [248,249].
miR-181a, which was shown to be upregulated in pancreatic cancer and inhibits RKIP mRNA and induces epithelial–mesenchymal transition such as cancer stem cell phenotype, is associated with poor survival [250].
miR-223, which was shown to be upregulated in pancreatic cancer and inhibits ZIC1 mRNA via PI3K/Akt/mTOR signaling pathway, is associated with poor survival [251].
miR-23b-3p, which was shown to be upregulated in pancreatic cancer and inhibits PTEN mRNA via the JAK/PI3K and Akt/NF-kappa B signaling pathways, is associated with poor survival [252].
miR-216a, which was shown to be downregulated in pancreatic cancer and inhibits WT1 mRNA and regulates KRT7 transcription, is associated with poor survival [254].
miR-139, which was shown to be downregulated in pancreatic cancer and inhibits RalB mRNA via the Ral/RAC/PI3K pathway, is associated with poor survival [255].
miR-1252-5p, which was shown to be downregulated in pancreatic cancer and inhibits NEDD9 mRNA. Importanly, miR-1252-5p is regulated by Myb, regulates NEDD9 mRNA inhibition, and is associated with poor survival [256].
miR-125b, which was shown to be downregulated in pancreatic cancer and inhibits NEDD9 mRNA via PI3K/AKT signaling pathway, is associated with poor survival [257].
miR-382, which was shown to be downregulated in pancreatic cancer and inhibits Anxa3 mRNA via PI3K/Akt signaling pathway, is associated with poor survival [258].
miR-519, which was shown to be downregulated in pancreatic cancer and inhibits PD-L1 mRNA, is associated with poor survival [259].
miR−155, which was shown to be upregulated in pancreatic cancer and inhibits TP53INP expression, is associated with poor survival [237,238].
Notably, miR-155-5p was upregulated in tumor tissues and plasma in pancreatic ductal adenocarcinoma, and the expressions in tissues were associated with tumor stage and poor prognosis [224,260,261].
It was also shown that long-term administration of gemcitabine in tumor cells led to the overexpression of miR-155-5p, inducing gemcitabine resistance via anti-apoptotic activity and that the elevated miR-155-5p expression correlated with chemoresistance and poor prognosis for pancreatic ductal adenocarcinoma patients receiving gemcitabine treatment [238].
Likewise, serum miR-373-3p was found to be downregulated in pancreatic cancer, and the miR-373-3p level was negatively correlated with tumor, node, and metastasis (TNM) stage, lymph node metastasis, and distant metastasis [262]. Furthermore, pancreatic cancer patients with reduced serum miR-373 levels exhibited shorter five-year overall survival [262].
Because individual miRNAs have less discriminatory power, a combination of miRNAs, such as miR-16 and miR-196 with CA19-9 as a combination biomarker panel, was shown to yield more robust results, as did a panel of seven miRNAs (i.e., miR-20a, miR-21, miR-24, miR-25, miR-99a, miR-185 and miR-191) which had already been shown to discriminate between PDAC and healthy sera [221]. On the other hand, a three-microRNA combination including miR-106b, miR-126, and miR-486 resulted in a slightly less accurate diagnosis [221].
Another study showed that changes in the levels of miR-25, GDF-15, and CA19-9 made accurate discrimination possible and that the plasma levels of the six miRNAs and MIC-1, CA19-9 were elevated in pancreatic cancer patients compared with those of healthy controls [263]. Among them, miR-20a, miR-21, miR-25, MIC-1, and CA19-9 could distinguish pancreatic cancer patients from those with other GI cancers or biliopancreatic diversion cancers [263]. These findings suggest that a combination of biomarkers has better diagnostic value compared with using a single marker.
Table 2 shows current miRNA human clinical trials as diagnostic biomarkers as of the data cutoff date of 30 June 2023.
Table 2.
Current miRNA human clinical trials as diagnostic biomarkers. Source: Clinicaltrials.gov (accessed on 6 July 2023). Key words: miRNA and pancreatic cancer. Data acquisition: 30 June 2023. Abbreviations: cfDNA: cell-free DNA; MEN1: Multiple endocrine neoplasia type 1; Nab-Paclitaxel: nanoparticle albumin–bound paclitaxel.
Collectively, circulating miRNAs are potentially effective minimally invasive or non-invasive cancer biomarkers for clinical applications including cancer screening at the early stage, subtype classification, and predicting drug sensitivity for treatment strategy selection, as well as screening for chemo- or radio-resistance of tumors.
However, challenges remain concerning the sensitivity, specificity, and applicability of potential circulating miRNA biomarkers and future work will warrant their utility as cancer biomarkers for clinical applications.
9. Challenges Involving miRNAs as Biomarkers in Clinical Applications
Because of the aggressive nature of pancreatic cancer and many others, and the lack of reliable and robust biomarkers for early diagnosis, prognosis, and prediction of disease course, miRNAs may provide an additional potential tool to facilitate diagnosis, prediction, monitoring, and management of various therapeutics used in the treatment of pancreatic cancer and many other cancers and potentially improve low survival rates [129].
As discussed in the literature [264], circulating miRNA levels reflect the physiological or pathological status of a person, thus the analysis of miRNA patterns for various disease states may not only lead to novel approaches for the diagnosis and prognosis of disease or condition but also for the selection of appropriate therapies and as predicter biomarkers for monitoring therapy response.
In addition, changes in miRNA signatures provide additional information about the gene networks and molecular pathways that mediate the pathology and regulate the interindividual variability of medical treatments [265].
Furthermore, miRNAs are highly stable, have a long half-life in biological specimens, and the methods for their analysis are readily available and well-established and do not require any special handling and can be applied to samples currently available.
Similarly, profiling of circulating miRNAs can be done with relatively high sensitivity and high specificity, depending on the material available and method used, and with relatively low cost using readily available standard techniques already employed in clinical laboratories such as Northern blot, microarray, quantitative real-time PCR (qRT-PCR), next-generation RNA sequencing, single molecular sequencing, and in situ hybridization [264,266,267].
As discussed in the literature [225], miRNAs have potential in pancreatic cancer management as biomarkers that may help in treatment selection or response evaluation as well as potential therapeutic targets. Despite extensive research on miRNAs as biomarkers in various cancers, data concerning miRNAs’ role in pancreatic cancer are relatively few.
However, despite their potential clinical applications, several challenges including widespread inconsistencies have been observed among the studies which limit the full application of circulating miRNAs as biomarkers in clinical practice [268]. To address these challenges, more research is needed in the detection of miRNAs with high specificity and sensitivity in specimens where RNA material is limited.
In addition, validation of the potential miRNA biomarkers in preclinical patient-derived in vitro and in vivo models, improved bioinformatics to better understand the various pathways of miRNA dysregulation, and validation studies in larger prospective and randomized clinical trials are needed to establish the clinical relevance of miRNAs as biomarkers in pancreatic cancer.
9.1. Heterogeneity of Methodologies, miRNA Source Specimens, and Lack of Controls
The main limiting factors in the application of miRNAs as biomarkers in clinical practice include the lack of consensus on methodologies and the use of different biological fluids such as serum, plasma, or extracellular vesicles (EVs), and others for miRNA profiling which introduces biological bias by the use of different matrices [204], there are also methodological and technical biases regarding the distinct isolation protocols.
Other preanalytical and analytical aspects such as technical variation involving RNA isolation, the lack of robust internal controls, and the impact of confounding factors such as sex, age, and concurrent therapies may affect the miRNA levels [269].
9.2. Sample Source and Physiological Variations and Low RNA Input
Other challenges involving miRNAs include a low concentration of miRNAs in circulation, sensitivity (a test’s ability to designate an individual with a disease as positive), specificity (a test’s ability to assess the exact component in a mixture), and selectivity (a test’s ability to differentiate the components in a mixture from each other) of the methods used for detecting miRNAs [266], cross-reactivity of miRNAs with different pathologies or disease states and normal physiological states, and stages of a particular disease.
In addition, as discussed in the literature [267], several other factors affect miRNA expression levels, including physiological fluctuations of miRNA expression levels during various cellular processes such as cell division, maturation, differentiation, and development [75,270,271,272,273,274] and age [275,276] or are affected by changes in environmental factors or exposures such as drug treatment and others [277,278].
In addition, miRNAs that are secreted in disease state by specific cells associated with disease, intracellular miRNAs, may also be secreted by other cell types through exosomes and conjugate to RNA-binding proteins, or they may be passively secreted after apoptosis or tissue injury [191].
As for disease pathogenesis, changes in cellular and circulating levels of miRNAs can either be a cause or result of developing pathogenic processes. Also, because miRNA levels have been shown to have a strong correlation with disease progression in various disease types including cancer and many others [21,22,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,53,55,56,57,58,60,61,64,66,279,280,281], several of these aspects must be considered when conducting miRNA measurements as disease biomarkers.
Furthermore, there have been variations in results among studies that often result from using patient tissue specimens which contain heterogeneous cell populations (e.g., the ductal, acinar, and islet cells, along with other inflammatory, fibroblastic components that will form the tumor microenvironment). Due to this intratumoral heterogeneity, the prediction of target genes of miRNAs is one of the major challenges of assigning miRNAs in large-scale applications, as each miRNA regulates more than one target and each target might be regulated by multiple miRNAs. In addition, the type of miRNAs might change during the course and stage of cancer, which further complicates the target prediction; however, this can also be beneficial for assigning a specific miRNA or several miRNAs to a specific stage of cancer. Therefore, more innovative approaches for predicting miRNA targets might be required to validate the predicted targets.
Several recent studies have demonstrated that liquid biopsy-based methods can accurately predict tumor response in pancreatic cancer and that sensitivity and specificity are increased when combined with CA19-9 [5,282,283,284,285,286,287,288]; however, there is not enough evidence to demonstrate that liquid biopsy improves patient outcomes or reduces unnecessary chemotherapy.
9.3. Methodological and Technical Variations in Detecting miRNAs
One of the main challenges in miRNA biomarker discovery is the poor reproducibility across technologies.
Sensitive and highly specific miRNA detection is required not only for studying physiological processes regulated by these RNAs but also for better understanding of relevant pathologies and their development as potential biomarkers. In addition, research on miRNA biogenesis, localization, function, and dysfunction often needs to be studied in a single cell, with single molecule sensitivity, and spatial as well as temporal resolution [267]. Further, depending on the modality of detection, assay sensitivities remain variable and are impacted by clinical stage, tumor burden, location of primary and metastatic tumor sites, and rates of cell turnover [5]. As a result, the detection of miRNAs can be affected by methods, measurement principles, and other technical factors [130].
There are several detection methods used for miRNA profiling, including Northern blot, microarray, qRT-PCR, next-generation RNA sequencing, single molecular sequencing, and in situ hybridization [266,267], and each of these methods has its strengths and limitations [289].
While assays such as Northern blot, qRT-PCR, microarray, next Generation RNA sequencing, and single molecular sequencing all provide some level of information about the miRNAs profiled, the specificity and sensitivity of each assay, as well as the ability to conduct them in high-throughput format and starting material as well as cost are highly variable. For example, Northern blot has high specificity and middle sensitivity and is an established technique but requires a large amount of sample, is tedious, and has very low throughput. qRT-PCR has moderate specificity and high sensitivity and is an established technique and can be automated but is medium throughput and cannot identify new miRNAs. Microarray has low specificity and low to middle sensitivity with low cost, high-throughput, and is fast; however, microarray is semiquantitative and cannot identify new miRNAs.
In situ hybridization has low specificity and low to middle sensitivity with low cost, low throughput, and slow processing of samples and requires a high level of expertise.
Next-generation RNA sequencing has very high specificity and middle sensitivity with middle high-throughput and whole-genome sequencing capability but is expensive and takes time to process samples and requires a high level of expertise.
Single molecular sequencing has moderate specificity and high sensitivity with middle high-throughput and whole-genome sequencing capability and is relatively fast but is very expensive.
Despite the advances in the development of next-generation RNA sequencing platforms aimed to analyze ultra-low quantities of RNA material, many of these approaches are biased and poorly reproducible with other RNA detection methods [290] due to the low RNA input requiring longer amplification cycles hence resulting in the introduction of amplification biases and the presence of overrepresented sequences with low to no clinical relevance [291].
In addition, the identification and quantification of miRNAs are challenging because of their short length, the existence of miRNA isomers, high homology within miRNA families, and O-methyl 3′ modifications [292].
9.4. Variations in Data Analysis and Bioinformatics Tools in the Analysis of miRNAs
Moreover, as discussed recently in the literature [190], several bioinformatic tools, databases, and algorithms have been developed in recent years that are available online to predict regulatory targets and mechanistic functions of each miRNA [101,293].
Importantly, multiple independent miRNA target prediction algorithms such as TargetScan [101] are used concurrently to increase the predictive power of these algorithms and for the prediction of miRNA-binding sites in their protein-coding genes and the biological pathways they regulate.
Furthermore, bioinformatic platforms including IPA/Ingenuity, Kegg, and others are being used to predict biological pathways and disease states that are regulated by miRNAs. Thus, streamlining miRNA research with miRNA bioinformatics platforms is key to facilitating the identification and development of miRNAs as clinically relevant biomarkers.
However, predicted targets and mechanistic functions of candidate miRNA biomarkers by various bioinformatics tools need validation in appropriate in vitro and in vivo preclinical models before testing them in clinical studies. For example, various preclinical models, including various immortalized human cell lines, patient-derived cells, and models such as organoids and in vivo patient-derived xenograft models, have been used to investigate the mechanisms of miRNA candidates.
One challenge is the specificity of the detection method for miRNAs due to their high degree of sequence homology, which is closely related to the type, medication, and prognosis of related diseases [266]. A recent report reviews [266] different methods used in miRNA detection.
Consequently, there is a need for the development of novel miRNA detection and analysis methods, technology platforms, and bioinformatics tools for easy, rapid, accurate, quantitative, robust, reproducible, and cost-effective detection and analysis of miRNAs in different types of specimens with high sensitivity and specificity and this need has attracted some attention from assay developers and biotech researchers [266,294,295,296].
Because each disease is fundamentally the consequence of abnormalities in the entire gene expression network and specific biological pathways, a panel of miRNAs might be a better strategy to understand the disease pathobiology. In many studies, the combination of a panel of miRNAs has been performed to increase the discriminative potential of the biomarker set [37,38,41,42,43,44,190].
Recently, due to its ability to analyze and aid in the interpretation of large datasets, artificial intelligence (AI), including machine learning algorithms, has been adapted for biomarker discovery, and has become a popular alternative statistical approach [264]. The advantages that AI provides are multiple. For example, the AI approach is not solely dependent on datasets involving circulating miRNAs, pharmacological, clinical patient data, and sociodemographic patient data to discriminate disease cases from controls or predict disease risk or outcome but is also capable of combining the value of several genes to produce a miRNA signature with predictive potential [264]. The value of this approach has shown its utility in the evaluation of cardiovascular conditions [297], suggesting that the results produced from these emerging approaches and tools will lead to identification of more robust miRNA biomarkers with stronger discriminative potential.
9.5. Poor Experimental Design in Clinical Studies
To date, several registered human clinical studies in the “clinicaltrials.gov” (accessed on 6 July 2023) database are exploring the utility of miRNAs as biomarkers across human conditions including various types of cancers, diabetes, lupus, coronary heart disease, epilepsy, depressive disorder, stroke, Addison’s disease, influenza, liver disease, and even toxic exposure to agents such as acetaminophen and several phase four trials that monitored select miRNAs as biomarkers for disease progression in patients receiving FDA-approved drugs [190].
In addition, as illustrated in Table 2, there are several ongoing or completed prospective human clinical trials for the assessment of the clinical utility of circulating miRNAs in pancreatic cancer including miRNA-25 as a diagnostic tool in pancreatic cancer (NCT03432624).
However, there are several challenges associated with these studies, including the insufficient sensitivity, specificity, and selectivity of miRNAs, especially in early-stage disease, due to the low abundance of circulating miRNAs, variability of detection methods used to measure miRNAs, and the lack of standardization of miRNA isolation and detection methods. The detection of miRNAs can be influenced by methods and measurement principles and other technical factors [130]. More importantly, there is a lack of comprehensive validation of miRNA signatures for a disease of interest in large prospective randomized controlled human trials, which is key to further validating the sensitivity, specificity, selectivity, and applicability of the candidate disease-specific miRNAs as biomarkers.
Notably, many miRNAs reported in the literature as associated with cancer and other conditions may be the outcome of underpowered studies due to small sample sizes, which often results in poor replication of results between studies and limits the potential utility of miRNAs as biomarkers in clinical applications [298].
Furthermore, due to many confounding factors used as criteria in clinical studies, clinical trials must consider avoiding the hurdle of limited sample size [23].
For example, although case-control studies based on patients with different pathologies and healthy individuals have been used in discovery studies to serve as controls and are useful for molecular phenotyping, this approach often does not sufficiently consider confounding factors such as age, sex, ethnicity, lifestyle, treatment history, history of diseases, concurrent therapies and treatments and comorbidities [264].
In addition, the high cost of analysis of miRNAs in a large number of samples, lack of replication of the results in independent clinical trials, and lack or low use of prospective, adequately powered large, and well-controlled randomized trials are the other factors that limit the translation and clinical applicability of the experimental findings.
Until these issues are addressed and the data published, widespread use of circulating miRNAs as biomarkers for diagnosis, monitoring of treatment response, and detection of recurrence of pancreatic cancer is unsupported.
10. Conclusions and Future Perspectives
Because many miRNAs are abnormally expressed or mutated in most malignant cancers such as pancreatic cancer and others, and since miRNAs could act as an oncogene or a tumor suppressor gene, they have emerged as potential biomarkers as well as therapeutic targets. In particular, circulating miRNAs which can be detected in various body fluids have become promising effective non-invasive cancer biomarkers for clinical application. Toward this goal, in recent years, significant progress has been made in miRNA profiling in liquid biopsy specimens. These non- or minimally invasive platforms thus hold significant promise and are likely to soon improve screening, diagnosis, cancer subtype classification, prognosis, and monitoring of therapy response and surveillance of pancreatic cancer and many other cancers and malignancies. The wide adoption of next-generation molecular profiling technologies such as high-throughput next-generation RNA sequencing including single-cell RNA sequencing is providing researchers with better insights into gene dysregulation in tumorigenesis and the discovery of novel RNA biomarkers including miRNAs and other non-coding-RNAs for cancer screening as well as for developing RNA-targeted or RNA-based cancer therapies.
Moreover, in addition to tissue-specific miRNA profiling, circulating miRNAs are being explored as potential minimally invasive biomarkers for clinical application, including cancer screening in the early stage, subtype classification, and predicting drug sensitivity for treatment strategy selection, as well as screening the chemo- or radio-resistance of tumors. Furthermore, miRNAs may provide important insights into tumor evolution and mechanisms of therapy resistance.
However, there are many challenges concerning the applicability of circulating miRNAs as biomarkers. For example, widespread adoption of miRNA biomarkers in routine clinical practice has been hindered by insufficient sensitivity, specificity, and selectivity of miRNAs, especially in early-stage disease and across different types of diseases. Because of this, a comprehensive validation and standardization of current miRNA profiling and analysis methodologies, as well as confirmation of candidate circulating miRNAs in large prospective randomized controlled trials to further validate the sensitivity, specificity, selectivity, and applicability of potential circulating miRNA biomarkers, are required.
It is also worth mentioning that, due to many confounding factors used as criteria in clinical applications such as age, sex, ethnicity, lifestyle, treatment history, history of disease, and others, it is mandatory to avoid the obstacle of limited sample size.
Given the potential to change clinical practice, miRNA biomarkers, especially those that can be used in early diagnosis of aggressive cancers such as pancreatic cancer, are rapidly improving and are expected to become part of the standard toolkit in pancreatic cancer management.
However, there needs to be significant efforts put in place to accomplish the clinical application of miRNAs as biomarkers, in particular, efforts need to focus on overcoming current methodological, technical, or analytical challenges and harmonization of miRNA isolation and quantification methods alongside the use of standard operating procedures, the development of automated and standardized assays, and the miniaturization of the methods to improve reproducibility between independent studies and more importantly for bench-to-bedside translation of the miRNA biomarkers for clinical applications.
To facilitate these, there is a need for the establishment of specific guidelines and protocols on best practices to augment the quality of the miRNA biomarker studies.
Furthermore, the implementation of miRNAs as biomarkers in clinical research also requires biobanks accessible to researchers to address the low availability of large and independent cohorts and importantly, cost-effectiveness.
Furthermore, a systematic and collaborative strategy that incorporates basic and clinical research and associated investigators, industry partners, and governmental agencies is key to help in translating early miRNA biomarkers into clinical applications.
Collectively, key to the success lies in the establishment of a research ecosystem favoring and supporting collaboration across various stakeholders from basic to clinical research to industry partners and improvement of funding opportunities provided by governmental agencies to facilitate the translation from bench to bedside.
Funding
This research received no external funding.
Institutional Review Board Statement
This article does not contain any studies with human participants or animals performed by any of the authors. Informed consent was not required for the preparation of this review article as it used secondary sources only.
Informed Consent Statement
The authors grant the publisher the sole and exclusive license of the full copyright in the Contribution, which license the Publisher hereby accepts.
Data Availability Statement
The data presented in this study are openly available in https://seer.cancer.gov, https://pubmed.ncbi.nlm.nih.gov/, https://clinicaltrials.gov/, and https://www.genomicseducation.hee.nhs.uk/genotes/knowledge-hub/non-coding-dna, all accessed on 26 July 2023.
Acknowledgments
The author thanks Legorreta Cancer Center at Brown University for providing the intellectual and collaborative environment, as well as the necessary resources, to prepare this review article.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Singhi, A.D.; Koay, E.J.; Chari, S.T.; Maitra, A. Early Detection of Pancreatic Cancer: Opportunities and Challenges. Gastroenterology 2019, 156, 2024–2040. [Google Scholar] [CrossRef] [PubMed]
- Mizrahi, J.D.; Surana, R.; Valle, J.W.; Shroff, R.T. Pancreatic cancer. Lancet 2020, 395, 2008–2020. [Google Scholar] [CrossRef]
- Advancing on pancreatic cancer. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 447. [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA A Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef]
- Raufi, A.G.; May, M.S.; Hadfield, M.J.; Seyhan, A.A.; El-Deiry, W.S. Advances in Liquid Biopsy Technology and Implications for Pancreatic Cancer. Int. J. Mol. Sci. 2023, 24, 4238. [Google Scholar] [CrossRef]
- SEER Cancer Statistics. Available online: https://seer.cancer.gov (accessed on 22 June 2022).
- Conroy, T.; Hammel, P.; Hebbar, M.; Ben Abdelghani, M.; Wei, A.C.; Raoul, J.L.; Chone, L.; Francois, E.; Artru, P.; Biagi, J.J.; et al. FOLFIRINOX or Gemcitabine as Adjuvant Therapy for Pancreatic Cancer. N. Engl. J. Med. 2018, 379, 2395–2406. [Google Scholar] [CrossRef] [PubMed]
- Moletta, L.; Serafini, S.; Valmasoni, M.; Pierobon, E.S.; Ponzoni, A.; Sperti, C. Surgery for Recurrent Pancreatic Cancer: Is It Effective? Cancers 2019, 11, 991. [Google Scholar] [CrossRef] [PubMed]
- Conroy, T.; Desseigne, F.; Ychou, M.; Bouche, O.; Guimbaud, R.; Becouarn, Y.; Adenis, A.; Raoul, J.L.; Gourgou-Bourgade, S.; de la Fouchardiere, C.; et al. FOLFIRINOX versus gemcitabine for metastatic pancreatic cancer. N. Engl. J. Med. 2011, 364, 1817–1825. [Google Scholar] [CrossRef]
- Goldstein, D.; El-Maraghi, R.H.; Hammel, P.; Heinemann, V.; Kunzmann, V.; Sastre, J.; Scheithauer, W.; Siena, S.; Tabernero, J.; Teixeira, L.; et al. nab-Paclitaxel plus gemcitabine for metastatic pancreatic cancer: Long-term survival from a phase III trial. J. Natl. Cancer Inst. 2015, 107, dju413. [Google Scholar] [CrossRef]
- Chhoda, A.; Lu, L.; Clerkin, B.M.; Risch, H.; Farrell, J.J. Current Approaches to Pancreatic Cancer Screening. Am. J. Pathol. 2019, 189, 22–35. [Google Scholar] [CrossRef]
- Luo, G.; Jin, K.; Deng, S.; Cheng, H.; Fan, Z.; Gong, Y.; Qian, Y.; Huang, Q.; Ni, Q.; Liu, C.; et al. Roles of CA19-9 in pancreatic cancer: Biomarker, predictor and promoter. Biochim. Biophys. Acta Rev. Cancer 2021, 1875, 188409. [Google Scholar] [PubMed]
- Ni, X.G.; Bai, X.F.; Mao, Y.L.; Shao, Y.F.; Wu, J.X.; Shan, Y.; Wang, C.F.; Wang, J.; Tian, Y.T.; Liu, Q.; et al. The clinical value of serum CEA, CA19-9, and CA242 in the diagnosis and prognosis of pancreatic cancer. Eur. J. Surg. Oncol. 2005, 31, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Smolarz, B.; Durczyński, A.; Romanowicz, H.; Hogendorf, P. The Role of microRNA in Pancreatic Cancer. Biomedicines 2021, 9, 1322. [Google Scholar] [CrossRef]
- Macconaill, L.E.; Garraway, L.A. Clinical implications of the cancer genome. J. Clin. Oncol. 2010, 28, 5219–5228. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Rands, C.M.; Meader, S.; Ponting, C.P.; Lunter, G. 8.2% of the Human genome is constrained: Variation in rates of turnover across functional element classes in the human lineage. PLoS Genet. 2014, 10, e1004525. [Google Scholar] [CrossRef]
- Dunham, I.; Kundaje, A.; Aldred, S.F.; Collins, P.J.; Davis, C.A.; Doyle, F.; Epstein, C.B.; Frietze, S.; Harrow, J.; Kaul, R.; et al. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar]
- Halldorsson, B.V.; Eggertsson, H.P.; Moore, K.H.S.; Hauswedell, H.; Eiriksson, O.; Ulfarsson, M.O.; Palsson, G.; Hardarson, M.T.; Oddsson, A.; Jensson, B.O.; et al. The sequences of 150,119 genomes in the UK Biobank. Nature 2022, 607, 732–740. [Google Scholar]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar]
- Li, Y.; Kowdley, K.V. MicroRNAs in Common Human Diseases. Genom. Proteom. Bioinform. 2012, 10, 246–253. [Google Scholar] [CrossRef]
- De Guire, V.; Robitaille, R.; Tetreault, N.; Guerin, R.; Menard, C.; Bambace, N.; Sapieha, P. Circulating miRNAs as sensitive and specific biomarkers for the diagnosis and monitoring of human diseases: Promises and challenges. Clin. Biochem. 2013, 46, 846–860. [Google Scholar]
- Condrat, C.E.; Thompson, D.C.; Barbu, M.G.; Bugnar, O.L.; Boboc, A.; Cretoiu, D.; Suciu, N.; Cretoiu, S.M.; Voinea, S.C. miRNAs as Biomarkers in Disease: Latest Findings Regarding Their Role in Diagnosis and Prognosis. Cells 2020, 9, 276. [Google Scholar]
- Gareev, I.; de Jesus Encarnacion Ramirez, M.; Goncharov, E.; Ivliev, D.; Shumadalova, A.; Ilyasova, T.; Wang, C. MiRNAs and lncRNAs in the regulation of innate immune signaling. Noncoding RNA Res. 2023, 8, 534–541. [Google Scholar] [PubMed]
- Pauley, K.M.; Cha, S.; Chan, E.K. MicroRNA in autoimmunity and autoimmune diseases. J. Autoimmun. 2009, 32, 189–194. [Google Scholar] [PubMed]
- Sullivan, C.S.; Ganem, D. MicroRNAs and viral infection. Mol. Cell 2005, 20, 3–7. [Google Scholar]
- Skalsky, R.L.; Cullen, B.R. Viruses, microRNAs, and host interactions. Annu. Rev. Microbiol. 2010, 64, 123–141. [Google Scholar]
- Barbu, M.G.; Condrat, C.E.; Thompson, D.C.; Bugnar, O.L.; Cretoiu, D.; Toader, O.D.; Suciu, N.; Voinea, S.C. MicroRNA Involvement in Signaling Pathways During Viral Infection. Front. Cell Dev. Biol. 2020, 8, 143. [Google Scholar] [PubMed]
- Abu-Izneid, T.; AlHajri, N.; Ibrahim, A.M.; Javed, M.N.; Salem, K.M.; Pottoo, F.H.; Kamal, M.A. Micro-RNAs in the regulation of immune response against SARS CoV-2 and other viral infections. J. Adv. Res. 2021, 30, 133–145. [Google Scholar]
- Elfimova, N.; Schlattjan, M.; Sowa, J.P.; Dienes, H.P.; Canbay, A.; Odenthal, M. Circulating microRNAs: Promising candidates serving as novel biomarkers of acute hepatitis. Front. Physiol. 2012, 3, 476. [Google Scholar] [PubMed]
- Li, Y.J.; Xu, M.; Gao, Z.H.; Wang, Y.Q.; Yue, Z.; Zhang, Y.X.; Li, X.X.; Zhang, C.; Xie, S.Y.; Wang, P.Y. Alterations of serum levels of BDNF-related miRNAs in patients with depression. PLoS ONE 2013, 8, e63648. [Google Scholar]
- Scott, K.A.; Hoban, A.E.; Clarke, G.; Moloney, G.M.; Dinan, T.G.; Cryan, J.F. Thinking small: Towards microRNA-based therapeutics for anxiety disorders. Expert. Opin. Investig. Drugs 2015, 24, 529–542. [Google Scholar]
- Liu, W.; Liu, C.; Zhu, J.; Shu, P.; Yin, B.; Gong, Y.; Qiang, B.; Yuan, J.; Peng, X. MicroRNA-16 targets amyloid precursor protein to potentially modulate Alzheimer’s-associated pathogenesis in SAMP8 mice. Neurobiol. Aging 2012, 33, 522–534. [Google Scholar]
- Weir, D.W.; Sturrock, A.; Leavitt, B.R. Development of biomarkers for Huntington’s disease. Lancet Neurol. 2011, 10, 573–590. [Google Scholar]
- Wang, R.; Li, N.; Zhang, Y.; Ran, Y.; Pu, J. Circulating microRNAs are promising novel biomarkers of acute myocardial infarction. Intern. Med. 2011, 50, 1789–1795. [Google Scholar]
- Recchioni, R.; Marcheselli, F.; Olivieri, F.; Ricci, S.; Procopio, A.D.; Antonicelli, R. Conventional and novel diagnostic biomarkers of acute myocardial infarction: A promising role for circulating microRNAs. Biomarkers 2013, 18, 547–558. [Google Scholar]
- Nunez Lopez, Y.O.; Coen, P.M.; Goodpaster, B.H.; Seyhan, A.A. Gastric bypass surgery with exercise alters plasma microRNAs that predict improvements in cardiometabolic risk. Int. J. Obes. 2017, 41, 1121–1130. [Google Scholar]
- Nunez Lopez, Y.O.; Garufi, G.; Pasarica, M.; Seyhan, A.A. Elevated and Correlated Expressions of miR-24, miR-30d, miR-146a, and SFRP-4 in Human Abdominal Adipose Tissue Play a Role in Adiposity and Insulin Resistance. Int. J. Endocrinol. 2018, 2018, 7351902. [Google Scholar] [PubMed]
- Wang, Q.; Wang, Y.; Minto, A.W.; Wang, J.; Shi, Q.; Li, X.; Quigg, R.J. MicroRNA-377 is up-regulated and can lead to increased fibronectin production in diabetic nephropathy. FASEB J. 2008, 22, 4126–4135. [Google Scholar] [PubMed]
- Gambardella, S.; Rinaldi, F.; Lepore, S.M.; Viola, A.; Loro, E.; Angelini, C.; Vergani, L.; Novelli, G.; Botta, A. Overexpression of microRNA-206 in the skeletal muscle from myotonic dystrophy type 1 patients. J. Transl. Med. 2010, 8, 48. [Google Scholar] [PubMed]
- Seyhan, A.A. microRNAs with different functions and roles in disease development and as potential biomarkers of diabetes: Progress and challenges. Mol. Biosyst. 2015, 11, 1217–1234. [Google Scholar] [PubMed]
- Seyhan, A.A.; Nunez Lopez, Y.O.; Xie, H.; Yi, F.; Mathews, C.; Pasarica, M.; Pratley, R.E. Pancreas-enriched miRNAs are altered in the circulation of subjects with diabetes: A pilot cross-sectional study. Sci. Rep. 2016, 6, 31479. [Google Scholar] [CrossRef]
- Nunez Lopez, Y.O.; Garufi, G.; Seyhan, A.A. Altered levels of circulating cytokines and microRNAs in lean and obese individuals with prediabetes and type 2 diabetes. Mol. Biosyst. 2016, 13, 106–121. [Google Scholar] [CrossRef] [PubMed]
- Nunez Lopez, Y.O.; Pittas, A.G.; Pratley, R.E.; Seyhan, A.A. Circulating levels of miR-7, miR-152 and miR-192 respond to vitamin D supplementation in adults with prediabetes and correlate with improvements in glycemic control. J. Nutr. Biochem. 2017, 49, 117–122. [Google Scholar] [CrossRef]
- Kluiver, J.; Poppema, S.; de Jong, D.; Blokzijl, T.; Harms, G.; Jacobs, S.; Kroesen, B.J.; van den Berg, A. BIC and miR-155 are highly expressed in Hodgkin, primary mediastinal and diffuse large B cell lymphomas. J. Pathol. 2005, 207, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D.; Sweet-Cordero, A.; Ebert, B.L.; Mak, R.H.; Ferrando, A.A.; et al. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef]
- Ventura, A.; Jacks, T. MicroRNAs and cancer: Short RNAs go a long way. Cell 2009, 136, 586–591. [Google Scholar] [CrossRef]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Lawler, S.; Chiocca, E.A. Emerging functions of microRNAs in glioblastoma. J. Neurooncol. 2009, 92, 297–306. [Google Scholar] [CrossRef]
- Suzuki, H.; Maruyama, R.; Yamamoto, E.; Kai, M. Epigenetic alteration and microRNA dysregulation in cancer. Front. Genet. 2013, 4, 258. [Google Scholar] [CrossRef]
- Di Leva, G.; Garofalo, M.; Croce, C.M. MicroRNAs in cancer. Annu. Rev. Pathol. 2014, 9, 287–314. [Google Scholar] [CrossRef]
- Hayes, J.; Peruzzi, P.P.; Lawler, S. MicroRNAs in cancer: Biomarkers, functions and therapy. Trends Mol. Med. 2014, 20, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Graveel, C.R.; Calderone, H.M.; Westerhuis, J.J.; Winn, M.E.; Sempere, L.F. Critical analysis of the potential for microRNA biomarkers in breast cancer management. Breast Cancer 2015, 7, 59–79. [Google Scholar] [PubMed]
- Lin, S.; Gregory, R.I. MicroRNA biogenesis pathways in cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [PubMed]
- Lulla, A.R.; Slifker, M.J.; Zhou, Y.; Lev, A.; Einarson, M.B.; Dicker, D.T.; El-Deiry, W.S. miR-6883 Family miRNAs Target CDK4/6 to Induce G(1) Phase Cell-Cycle Arrest in Colon Cancer Cells. Cancer Res. 2017, 77, 6902–6913. [Google Scholar] [CrossRef]
- Biswas, S. MicroRNAs as Therapeutic Agents: The Future of the Battle against Cancer. Curr. Top. Med. Chem. 2018, 18, 2544–2554. [Google Scholar] [CrossRef]
- Hu, W.; Tan, C.; He, Y.; Zhang, G.; Xu, Y.; Tang, J. Functional miRNAs in breast cancer drug resistance. Oncotargets Ther. 2018, 11, 1529–1541. [Google Scholar] [CrossRef]
- Plotnikova, O.; Baranova, A.; Skoblov, M. Comprehensive Analysis of Human microRNA-mRNA Interactome. Front. Genet. 2019, 10, 933. [Google Scholar] [CrossRef]
- Ali Syeda, Z.; Langden, S.S.S.; Munkhzul, C.; Lee, M.; Song, S.J. Regulatory Mechanism of MicroRNA Expression in Cancer. Int. J. Mol. Sci. 2020, 21, 1723. [Google Scholar] [CrossRef]
- Annese, T.; Tamma, R.; De Giorgis, M.; Ribatti, D. microRNAs Biogenesis, Functions and Role in Tumor Angiogenesis. Front. Oncol. 2020, 10, 581007. [Google Scholar] [CrossRef]
- Fathi, M.; Ghafouri-Fard, S.; Abak, A.; Taheri, M. Emerging roles of miRNAs in the development of pancreatic cancer. Biomed. Pharmacother. 2021, 141, 111914. [Google Scholar] [CrossRef] [PubMed]
- Galka-Marciniak, P.; Urbanek-Trzeciak, M.O.; Nawrocka, P.M.; Kozlowski, P. A pan-cancer atlas of somatic mutations in miRNA biogenesis genes. Nucleic Acids Res. 2021, 49, 601–620. [Google Scholar] [CrossRef] [PubMed]
- Pajares, M.J.; Alemany-Cosme, E.; Goni, S.; Bandres, E.; Palanca-Ballester, C.; Sandoval, J. Epigenetic Regulation of microRNAs in Cancer: Shortening the Distance from Bench to Bedside. Int. J. Mol. Sci. 2021, 22, 7350. [Google Scholar] [CrossRef]
- Inoue, J.; Inazawa, J. Cancer-associated miRNAs and their therapeutic potential. J. Hum. Genet. 2021, 66, 937–945. [Google Scholar] [CrossRef]
- Smolarz, B.; Durczyński, A.; Romanowicz, H.; Szyłło, K.; Hogendorf, P. miRNAs in Cancer (Review of Literature). Int. J. Mol. Sci. 2022, 23, 2805. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Meng, Q.; Qian, J.; Li, M.; Gu, C.; Yang, Y. Review: RNA-based diagnostic markers discovery and therapeutic targets development in cancer. Pharmacol. Ther. 2022, 234, 108123. [Google Scholar] [CrossRef] [PubMed]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—microRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef]
- Ding, J.; Cao, Y.; Qi, C.; Zong, Z. Dysregulated microRNAs participate in the crosstalk between colorectal cancer and atrial fibrillation. Hum. Cell 2023, 36, 1336–1342. [Google Scholar] [CrossRef]
- Ricarte-Filho, J.C.; Casado-Medrano, V.; Reichenberger, E.; Spangler, Z.; Scheerer, M.; Isaza, A.; Baran, J.; Patel, T.; MacFarland, S.P.; Brodeur, G.M.; et al. DICER1 RNase IIIb domain mutations trigger widespread miRNA dysregulation and MAPK activation in pediatric thyroid cancer. Front. Endocrinol. 2023, 14, 1083382. [Google Scholar] [CrossRef]
- Menon, A.; Abd-Aziz, N.; Khalid, K.; Poh, C.L.; Naidu, R. miRNA: A Promising Therapeutic Target in Cancer. Int. J. Mol. Sci. 2022, 23, 11502. [Google Scholar] [CrossRef]
- Hata, A.; Lieberman, J. Dysregulation of microRNA biogenesis and gene silencing in cancer. Sci. Signal. 2015, 8, re3. [Google Scholar] [CrossRef]
- Hata, A.; Kashima, R. Dysregulation of microRNA biogenesis machinery in cancer. Crit. Rev. Biochem. Mol. Biol. 2016, 51, 121–134. [Google Scholar] [CrossRef]
- Bortoletto, A.S.; Parchem, R.J. KRAS Hijacks the miRNA Regulatory Pathway in Cancer. Cancer Res. 2023, 83, 1563–1572. [Google Scholar] [CrossRef]
- Wienholds, E.; Kloosterman, W.P.; Miska, E.; Alvarez-Saavedra, E.; Berezikov, E.; de Bruijn, E.; Horvitz, H.R.; Kauppinen, S.; Plasterk, R.H. MicroRNA expression in zebrafish embryonic development. Science 2005, 309, 310–311. [Google Scholar] [CrossRef]
- Aboobaker, A.A.; Tomancak, P.; Patel, N.; Rubin, G.M.; Lai, E.C. Drosophila microRNAs exhibit diverse spatial expression patterns during embryonic development. Proc. Natl. Acad. Sci. USA 2005, 102, 18017–18022. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.C.; Harland, R.M. Expression of microRNAs during embryonic development of Xenopus tropicalis. Gene Expr. Patterns 2008, 8, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Li, L.J.; Leng, R.X.; Fan, Y.G.; Pan, H.F.; Ye, D.Q. Translation of noncoding RNAs: Focus on lncRNAs, pri-miRNAs, and circRNAs. Exp. Cell Res. 2017, 361, 1–8. [Google Scholar] [CrossRef]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef]
- Pasquinelli, A.E.; Reinhart, B.J.; Slack, F.; Martindale, M.Q.; Kuroda, M.I.; Maller, B.; Hayward, D.C.; Ball, E.E.; Degnan, B.; Müller, P.; et al. Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature 2000, 408, 86–89. [Google Scholar] [CrossRef]
- Li, S.C.; Chan, W.C.; Hu, L.Y.; Lai, C.H.; Hsu, C.N.; Lin, W.C. Identification of homologous microRNAs in 56 animal genomes. Genomics 2010, 96, 1–9. [Google Scholar] [CrossRef]
- Friedländer, M.R.; Lizano, E.; Houben, A.J.S.; Bezdan, D.; Báñez-Coronel, M.; Kudla, G.; Mateu-Huertas, E.; Kagerbauer, B.; González, J.; Chen, K.C.; et al. Evidence for the biogenesis of more than 1000 novel human microRNAs. Genome Biol. 2014, 15, R57. [Google Scholar] [CrossRef]
- Griffiths-Jones, S.; Grocock, R.J.; van Dongen, S.; Bateman, A.; Enright, A.J. miRBase: MicroRNA sequences, targets and gene nomenclature. Nucleic Acids Res. 2006, 34, D140–D144. [Google Scholar] [CrossRef]
- Khraiwesh, B.; Arif, M.A.; Seumel, G.I.; Ossowski, S.; Weigel, D.; Reski, R.; Frank, W. Transcriptional control of gene expression by microRNAs. Cell 2010, 140, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Samad, A.F.A.; Sajad, M.; Nazaruddin, N.; Fauzi, I.A.; Murad, A.M.A.; Zainal, Z.; Ismail, I. MicroRNA and Transcription Factor: Key Players in Plant Regulatory Network. Front. Plant Sci. 2017, 8, 565. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Cui, Q.; Wang, J.; Zhou, Y. TransmiR v2.0: An updated transcription factor-microRNA regulation database. Nucleic Acids Res. 2018, 47, D253–D258. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.Y.; Meister, G. microRNA-guided posttranscriptional gene regulation. Biol. Chem. 2005, 386, 1205–1218. [Google Scholar] [CrossRef]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Pu, M.; Chen, J.; Tao, Z.; Miao, L.; Qi, X.; Wang, Y.; Ren, J. Regulatory network of miRNA on its target: Coordination between transcriptional and post-transcriptional regulation of gene expression. Cell. Mol. Life Sci. 2019, 76, 441–451. [Google Scholar] [CrossRef]
- van Rooij, E. The art of microRNA research. Circ. Res. 2011, 108, 219–234. [Google Scholar] [CrossRef]
- Guo, Z.; Maki, M.; Ding, R.; Yang, Y.; Zhang, B.; Xiong, L. Genome-wide survey of tissue-specific microRNA and transcription factor regulatory networks in 12 tissues. Sci. Rep. 2014, 4, 5150. [Google Scholar] [CrossRef] [PubMed]
- de Rie, D.; Abugessaisa, I.; Alam, T.; Arner, E.; Arner, P.; Ashoor, H.; Åström, G.; Babina, M.; Bertin, N.; Burroughs, A.M.; et al. An integrated expression atlas of miRNAs and their promoters in human and mouse. Nat. Biotechnol. 2017, 35, 872–878. [Google Scholar] [CrossRef] [PubMed]
- Alles, J.; Fehlmann, T.; Fischer, U.; Backes, C.; Galata, V.; Minet, M.; Hart, M.; Abu-Halima, M.; Grässer, F.A.; Lenhof, H.P.; et al. An estimate of the total number of true human miRNAs. Nucleic Acids Res. 2019, 47, 3353–3364. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Ludwig, N.; Leidinger, P.; Becker, K.; Backes, C.; Fehlmann, T.; Pallasch, C.; Rheinheimer, S.; Meder, B.; Stähler, C.; Meese, E.; et al. Distribution of miRNA expression across human tissues. Nucleic Acids Res. 2016, 44, 3865–3877. [Google Scholar] [CrossRef]
- Beck, J.D.; Reidenbach, D.; Salomon, N.; Sahin, U.; Türeci, Ö.; Vormehr, M.; Kranz, L.M. mRNA therapeutics in cancer immunotherapy. Mol. Cancer 2021, 20, 69. [Google Scholar] [CrossRef] [PubMed]
- O’Carroll, D.; Schaefer, A. General principals of miRNA biogenesis and regulation in the brain. Neuropsychopharmacology 2013, 38, 39–54. [Google Scholar] [CrossRef]
- Rodriguez, A.; Griffiths-Jones, S.; Ashurst, J.L.; Bradley, A. Identification of mammalian microRNA host genes and transcription units. Genome Res. 2004, 14, 1902–1910. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef]
- Iwasaki, S.; Kobayashi, M.; Yoda, M.; Sakaguchi, Y.; Katsuma, S.; Suzuki, T.; Tomari, Y. Hsc70/Hsp90 chaperone machinery mediates ATP-dependent RISC loading of small RNA duplexes. Mol. Cell 2010, 39, 292–299. [Google Scholar] [CrossRef]
- Shukla, G.C.; Singh, J.; Barik, S. MicroRNAs: Processing, Maturation, Target Recognition and Regulatory Functions. Mol. Cell Pharmacol. 2011, 3, 83–92. [Google Scholar]
- Akgul, B.; Erdogan, I. Intracytoplasmic Re-localization of miRISC Complexes. Front. Genet. 2018, 9, 403. [Google Scholar] [CrossRef] [PubMed]
- Selbach, M.; Schwanhäusser, B.; Thierfelder, N.; Fang, Z.; Khanin, R.; Rajewsky, N. Widespread changes in protein synthesis induced by microRNAs. Nature 2008, 455, 58–63. [Google Scholar] [CrossRef]
- Uhlmann, S.; Mannsperger, H.; Zhang, J.D.; Horvat, E.; Schmidt, C.; Küblbeck, M.; Henjes, F.; Ward, A.; Tschulena, U.; Zweig, K.; et al. Global microRNA level regulation of EGFR-driven cell-cycle protein network in breast cancer. Mol. Syst. Biol. 2012, 8, 570. [Google Scholar] [CrossRef]
- Grimson, A.; Farh, K.K.; Johnston, W.K.; Garrett-Engele, P.; Lim, L.P.; Bartel, D.P. MicroRNA targeting specificity in mammals: Determinants beyond seed pairing. Mol. Cell 2007, 27, 91–105. [Google Scholar] [CrossRef]
- Saetrom, P.; Heale, B.S.; Snøve, O., Jr.; Aagaard, L.; Alluin, J.; Rossi, J.J. Distance constraints between microRNA target sites dictate efficacy and cooperativity. Nucleic Acids Res. 2007, 35, 2333–2342. [Google Scholar] [CrossRef] [PubMed]
- Desterro, J.; Bak-Gordon, P.; Carmo-Fonseca, M. Targeting mRNA processing as an anticancer strategy. Nat. Rev. Drug Discov. 2020, 19, 112–129. [Google Scholar] [CrossRef]
- Vogelstein, B.; Papadopoulos, N.; Velculescu, V.E.; Zhou, S.; Diaz, L.A., Jr.; Kinzler, K.W. Cancer genome landscapes. Science 2013, 339, 1546–1558. [Google Scholar] [CrossRef]
- Choudhury, Y.; Tay, F.C.; Lam, D.H.; Sandanaraj, E.; Tang, C.; Ang, B.T.; Wang, S. Attenuated adenosine-to-inosine editing of microRNA-376a* promotes invasiveness of glioblastoma cells. J. Clin. Investig. 2012, 122, 4059–4076. [Google Scholar] [CrossRef] [PubMed]
- Stahlhut, C.; Slack, F.J. MicroRNAs and the cancer phenotype: Profiling, signatures and clinical implications. Genome Med. 2013, 5, 111. [Google Scholar] [CrossRef] [PubMed]
- Dvinge, H.; Git, A.; Gräf, S.; Salmon-Divon, M.; Curtis, C.; Sottoriva, A.; Zhao, Y.; Hirst, M.; Armisen, J.; Miska, E.A.; et al. The shaping and functional consequences of the microRNA landscape in breast cancer. Nature 2013, 497, 378–382. [Google Scholar] [CrossRef]
- Kim, T.M.; Huang, W.; Park, R.; Park, P.J.; Johnson, M.D. A developmental taxonomy of glioblastoma defined and maintained by MicroRNAs. Cancer Res. 2011, 71, 3387–3399. [Google Scholar] [CrossRef] [PubMed]
- Manterola, L.; Guruceaga, E.; Gállego Pérez-Larraya, J.; González-Huarriz, M.; Jauregui, P.; Tejada, S.; Diez-Valle, R.; Segura, V.; Samprón, N.; Barrena, C.; et al. A small noncoding RNA signature found in exosomes of GBM patient serum as a diagnostic tool. Neuro Oncol. 2014, 16, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Kasinski, A.L.; Slack, F.J. MicroRNAs en route to the clinic: Progress in validating and targeting microRNAs for cancer therapy. Nat. Rev. Cancer 2011, 11, 849–864. [Google Scholar] [CrossRef]
- Shankar, S.; Tien, J.C.-Y.; Siebenaler, R.F.; Chugh, S.; Dommeti, V.L.; Zelenka-Wang, S.; Wang, X.-M.; Apel, I.J.; Waninger, J.; Eyunni, S.; et al. An essential role for Argonaute 2 in EGFR-KRAS signaling in pancreatic cancer development. Nat. Commun. 2020, 11, 2817. [Google Scholar] [CrossRef]
- Eder, M.; Scherr, M. MicroRNA and lung cancer. N. Engl. J. Med. 2005, 352, 2446–2448. [Google Scholar] [CrossRef]
- Gonzales-Aloy, E.; Connerty, P.; Salik, B.; Liu, B.; Woo, A.J.; Haber, M.; Norris, M.D.; Wang, J.; Wang, J.Y. miR-101 suppresses the development of MLL-rearranged acute myeloid leukemia. Haematologica 2019, 104, e296–e299. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, X.; Cheng, R.; Yang, F.; Yu, M.; Wang, C.; Cui, S.; Hong, Y.; Liang, H.; Liu, M.; et al. The Jun/miR-22/HuR regulatory axis contributes to tumourigenesis in colorectal cancer. Mol. Cancer 2018, 17, 11. [Google Scholar] [CrossRef]
- Zhu, S.; Cheng, X.; Wang, R.; Tan, Y.; Ge, M.; Li, D.; Xu, Q.; Sun, Y.; Zhao, C.; Chen, S.; et al. Restoration of microRNA function impairs MYC-dependent maintenance of MLL leukemia. Leukemia 2020, 34, 2484–2488. [Google Scholar] [CrossRef]
- Fu, Z.; Wang, L.; Li, S.; Chen, F.; Au-Yeung, K.K.; Shi, C. MicroRNA as an Important Target for Anticancer Drug Development. Front. Pharmacol. 2021, 12, 736323. [Google Scholar] [CrossRef]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OncomiR or Tumor Suppressor? The Duplicity of MicroRNAs in Cancer. Cancer Res. 2016, 76, 3666–3670. [Google Scholar] [CrossRef] [PubMed]
- Gambari, R.; Brognara, E.; Spandidos, D.A.; Fabbri, E. Targeting oncomiRNAs and mimicking tumor suppressor miRNAs: Νew trends in the development of miRNA therapeutic strategies in oncology (Review). Int. J. Oncol. 2016, 49, 5–32. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.C.S. OncomiRs: The discovery and progress of microRNAs in cancers. Mol. Cancer 2007, 6, 60. [Google Scholar] [CrossRef] [PubMed]
- Frixa, T.; Donzelli, S.; Blandino, G. Oncogenic MicroRNAs: Key Players in Malignant Transformation. Cancers 2015, 7, 2466–2485. [Google Scholar] [CrossRef]
- Zhou, K.; Liu, M.; Cao, Y. New Insight into microRNA Functions in Cancer: Oncogene-microRNA-Tumor Suppressor Gene Network. Front. Mol. Biosci. 2017, 4, 46. [Google Scholar] [CrossRef]
- Shenouda, S.K.; Alahari, S.K. MicroRNA function in cancer: Oncogene or a tumor suppressor? Cancer Metastasis. Rev. 2009, 28, 369–378. [Google Scholar] [CrossRef]
- Daoud, A.Z.; Mulholland, E.J.; Cole, G.; McCarthy, H.O. MicroRNAs in Pancreatic Cancer: Biomarkers, prognostic, and therapeutic modulators. BMC Cancer 2019, 19, 1130. [Google Scholar] [CrossRef]
- Wang, H.; Peng, R.; Wang, J.; Qin, Z.; Xue, L. Circulating microRNAs as potential cancer biomarkers: The advantage and disadvantage. Clin. Epigenetics 2018, 10, 59. [Google Scholar] [CrossRef]
- Landgraf, P.; Rusu, M.; Sheridan, R.; Sewer, A.; Iovino, N.; Aravin, A.; Pfeffer, S.; Rice, A.; Kamphorst, A.O.; Landthaler, M.; et al. A mammalian microRNA expression atlas based on small RNA library sequencing. Cell 2007, 129, 1401–1414. [Google Scholar] [CrossRef]
- Vardapour, R.; Kehl, T.; Kneitz, S.; Ludwig, N.; Meese, E.; Lenhof, H.P.; Gessler, M. The DGCR8 E518K mutation found in Wilms tumors leads to a partial miRNA processing defect that alters gene expression patterns and biological processes. Carcinogenesis 2022, 43, 82–93. [Google Scholar] [CrossRef]
- Rivera, B.; Nadaf, J.; Fahiminiya, S.; Apellaniz-Ruiz, M.; Saskin, A.; Chong, A.S.; Sharma, S.; Wagener, R.; Revil, T.; Condello, V.; et al. DGCR8 microprocessor defect characterizes familial multinodular goiter with schwannomatosis. J. Clin. Investig. 2020, 130, 1479–1490. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, L.; Canberk, S.; Macedo, S.; Soares, P.; Vinagre, J. DGCR8 Microprocessor Subunit Mutation and Expression Deregulation in Thyroid Lesions. Int. J. Mol. Sci. 2022, 23, 14812. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Lv, Z.; Ding, H.; Fang, X.; Sun, M. Association of miRNA biosynthesis genes DROSHA and DGCR8 polymorphisms with cancer susceptibility: A systematic review and meta-analysis. Biosci. Rep. 2018, 38, BSR20180072. [Google Scholar] [CrossRef] [PubMed]
- Li, B.K.; Vasiljevic, A.; Dufour, C.; Yao, F.; Ho, B.L.B.; Lu, M.; Hwang, E.I.; Gururangan, S.; Hansford, J.R.; Fouladi, M.; et al. Pineoblastoma segregates into molecular sub-groups with distinct clinico-pathologic features: A Rare Brain Tumor Consortium registry study. Acta Neuropathol. 2020, 139, 223–241. [Google Scholar] [CrossRef]
- Rotunno, M.; Zhao, Y.; Bergen, A.W.; Koshiol, J.; Burdette, L.; Rubagotti, M.; Linnoila, R.I.; Marincola, F.M.; Bertazzi, P.A.; Pesatori, A.C.; et al. Inherited polymorphisms in the RNA-mediated interference machinery affect microRNA expression and lung cancer survival. Br. J. Cancer 2010, 103, 1870–1874. [Google Scholar] [CrossRef]
- Permuth-Wey, J.; Kim, D.; Tsai, Y.Y.; Lin, H.Y.; Chen, Y.A.; Barnholtz-Sloan, J.; Birrer, M.J.; Bloom, G.; Chanock, S.J.; Chen, Z.; et al. LIN28B polymorphisms influence susceptibility to epithelial ovarian cancer. Cancer Res. 2011, 71, 3896–3903. [Google Scholar] [CrossRef]
- Felicio, P.S.; Grasel, R.S.; Campacci, N.; de Paula, A.E.; Galvão, H.C.R.; Torrezan, G.T.; Sabato, C.S.; Fernandes, G.C.; Souza, C.P.; Michelli, R.D.; et al. Whole-exome sequencing of non-BRCA1/BRCA2 mutation carrier cases at high-risk for hereditary breast/ovarian cancer. Hum. Mutat. 2021, 42, 290–299. [Google Scholar] [CrossRef]
- Liang, D.; Meyer, L.; Chang, D.W.; Lin, J.; Pu, X.; Ye, Y.; Gu, J.; Wu, X.; Lu, K. Genetic variants in MicroRNA biosynthesis pathways and binding sites modify ovarian cancer risk, survival, and treatment response. Cancer Res. 2010, 70, 9765–9776. [Google Scholar] [CrossRef]
- Mohammadpour-Gharehbagh, A.; Heidari, Z.; Eskandari, M.; Aryan, A.; Salimi, S. Association between Genetic Polymorphisms in microRNA Machinery Genes and Risk of Papillary Thyroid Carcinoma. Pathol. Oncol. Res. 2020, 26, 1235–1241. [Google Scholar] [CrossRef]
- Guénard, F.; Labrie, Y.; Ouellette, G.; Beauparlant, C.J.; Durocher, F. Genetic sequence variations of BRCA1-interacting genes AURKA, BAP1, BARD1 and DHX9 in French Canadian families with high risk of breast cancer. J. Hum. Genet. 2009, 54, 152–161. [Google Scholar] [CrossRef]
- Melo, S.A.; Moutinho, C.; Ropero, S.; Calin, G.A.; Rossi, S.; Spizzo, R.; Fernandez, A.F.; Davalos, V.; Villanueva, A.; Montoya, G.; et al. A genetic defect in exportin-5 traps precursor microRNAs in the nucleus of cancer cells. Cancer Cell 2010, 18, 303–315. [Google Scholar] [CrossRef]
- Liao, Y.; Liao, Y.; Li, J.; Liu, L.; Li, J.; Wan, Y.; Peng, L. Genetic variants in miRNA machinery genes associated with clinicopathological characteristics and outcomes of gastric cancer patients. Int. J. Biol. Markers 2018, 33, 301–307. [Google Scholar] [CrossRef]
- Palculict, T.B.; Ruteshouser, E.C.; Fan, Y.; Wang, W.; Strong, L.; Huff, V. Identification of germline DICER1 mutations and loss of heterozygosity in familial Wilms tumour. J. Med. Genet. 2016, 53, 385–388. [Google Scholar] [CrossRef] [PubMed]
- Schultz, K.A.; Pacheco, M.C.; Yang, J.; Williams, G.M.; Messinger, Y.; Hill, D.A.; Dehner, L.P.; Priest, J.R. Ovarian sex cord-stromal tumors, pleuropulmonary blastoma and DICER1 mutations: A report from the International Pleuropulmonary Blastoma Registry. Gynecol. Oncol. 2011, 122, 246–250. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.K.; Sabbaghian, N.; Xu, B.; Addidou-Kalucki, S.; Bernard, C.; Zou, D.; Reeve, A.E.; Eccles, M.R.; Cole, C.; Choong, C.S.; et al. Biallelic DICER1 mutations occur in Wilms tumours. J. Pathol. 2013, 230, 154–164. [Google Scholar] [CrossRef]
- Caruso, S.; Calderaro, J.; Letouzé, E.; Nault, J.C.; Couchy, G.; Boulai, A.; Luciani, A.; Zafrani, E.S.; Bioulac-Sage, P.; Seror, O.; et al. Germline and somatic DICER1 mutations in familial and sporadic liver tumors. J. Hepatol. 2017, 66, 734–742. [Google Scholar] [CrossRef]
- Heravi-Moussavi, A.; Anglesio, M.S.; Cheng, S.W.; Senz, J.; Yang, W.; Prentice, L.; Fejes, A.P.; Chow, C.; Tone, A.; Kalloger, S.E.; et al. Recurrent somatic DICER1 mutations in nonepithelial ovarian cancers. N. Engl. J. Med. 2012, 366, 234–242. [Google Scholar] [CrossRef]
- Witkowski, L.; Mattina, J.; Schönberger, S.; Murray, M.J.; Choong, C.S.; Huntsman, D.G.; Reis-Filho, J.S.; McCluggage, W.G.; Nicholson, J.C.; Coleman, N.; et al. DICER1 hotspot mutations in non-epithelial gonadal tumours. Br. J. Cancer 2013, 109, 2744–2750. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, D.; Li, Y.; Bian, L.; Ma, T.; Xie, M. DICER1 mutations in a patient with an ovarian Sertoli-Leydig tumor, well-differentiated fetal adenocarcinoma of the lung, and familial multinodular goiter. Eur. J. Med. Genet. 2014, 57, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Brenneman, M.; Field, A.; Yang, J.; Williams, G.; Doros, L.; Rossi, C.; Schultz, K.A.; Rosenberg, A.; Ivanovich, J.; Turner, J.; et al. Temporal order of RNase IIIb and loss-of-function mutations during development determines phenotype in pleuropulmonary blastoma/DICER1 syndrome: A unique variant of the two-hit tumor suppression model. F1000Research 2015, 4, 214. [Google Scholar] [CrossRef]
- Garre, P.; Pérez-Segura, P.; Díaz-Rubio, E.; Caldés, T.; de la Hoya, M. Reassessing the TARBP2 mutation rate in hereditary nonpolyposis colorectal cancer. Nat. Genet. 2010, 42, 817–818. [Google Scholar] [CrossRef]
- Torrezan, G.T.; Ferreira, E.N.; Nakahata, A.M.; Barros, B.D.; Castro, M.T.; Correa, B.R.; Krepischi, A.C.; Olivieri, E.H.; Cunha, I.W.; Tabori, U.; et al. Recurrent somatic mutation in DROSHA induces microRNA profile changes in Wilms tumour. Nat. Commun. 2014, 5, 4039. [Google Scholar] [CrossRef]
- Caramuta, S.; Lee, L.; Ozata, D.M.; Akçakaya, P.; Xie, H.; Höög, A.; Zedenius, J.; Bäckdahl, M.; Larsson, C.; Lui, W.O. Clinical and functional impact of TARBP2 over-expression in adrenocortical carcinoma. Endocr. Relat. Cancer 2013, 20, 551–564. [Google Scholar] [CrossRef]
- Chang, S.S.; Smith, I.; Glazer, C.; Hennessey, P.; Califano, J.A. EIF2C is overexpressed and amplified in head and neck squamous cell carcinoma. ORL J. Otorhinolaryngol. Relat. Spec. 2010, 72, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chen, L.; Barlogie, B.; Stephens, O.; Wu, X.; Williams, D.R.; Cartron, M.A.; van Rhee, F.; Nair, B.; Waheed, S.; et al. High-risk myeloma is associated with global elevation of miRNAs and overexpression of EIF2C2/AGO2. Proc. Natl. Acad. Sci. USA 2010, 107, 7904–7909. [Google Scholar] [CrossRef] [PubMed]
- Casey, M.C.; Prakash, A.; Holian, E.; McGuire, A.; Kalinina, O.; Shalaby, A.; Curran, C.; Webber, M.; Callagy, G.; Bourke, E.; et al. Quantifying Argonaute 2 (Ago2) expression to stratify breast cancer. BMC Cancer 2019, 19, 712. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Huang, J.; Zhou, Y.; Zhou, Y.; Yu, L.; Li, H.; Hou, L.; Zhu, L.; Ge, D.; Zeng, Y.; et al. Germline and somatic variations influence the somatic mutational signatures of esophageal squamous cell carcinomas in a Chinese population. BMC Genom. 2018, 19, 538. [Google Scholar] [CrossRef]
- Kim, M.S.; Oh, J.E.; Kim, Y.R.; Park, S.W.; Kang, M.R.; Kim, S.S.; Ahn, C.H.; Yoo, N.J.; Lee, S.H. Somatic mutations and losses of expression of microRNA regulation-related genes AGO2 and TNRC6A in gastric and colorectal cancers. J. Pathol. 2010, 221, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Szafranska, A.E.; Davison, T.S.; John, J.; Cannon, T.; Sipos, B.; Maghnouj, A.; Labourier, E.; Hahn, S.A. MicroRNA expression alterations are linked to tumorigenesis and non-neoplastic processes in pancreatic ductal adenocarcinoma. Oncogene 2007, 26, 4442–4452. [Google Scholar] [CrossRef]
- Jiao, L.R.; Frampton, A.E.; Jacob, J.; Pellegrino, L.; Krell, J.; Giamas, G.; Tsim, N.; Vlavianos, P.; Cohen, P.; Ahmad, R.; et al. MicroRNAs targeting oncogenes are down-regulated in pancreatic malignant transformation from benign tumors. PLoS ONE 2012, 7, e32068. [Google Scholar] [CrossRef] [PubMed]
- Schetter, A.J.; Okayama, H.; Harris, C.C. The role of microRNAs in colorectal cancer. Cancer J. 2012, 18, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.L.; Tsai, Y.M.; Lien, C.T.; Kuo, P.L.; Hung, A.J. The Roles of MicroRNA in Lung Cancer. Int. J. Mol. Sci. 2019, 20, 1611. [Google Scholar] [CrossRef]
- Le Rolle, A.F.; Chiu, T.K.; Zeng, Z.; Shia, J.; Weiser, M.R.; Paty, P.B.; Chiu, V.K. Oncogenic KRAS activates an embryonic stem cell-like program in human colon cancer initiation. Oncotarget 2016, 7, 2159–2174. [Google Scholar] [CrossRef]
- Waters, A.M.; Der, C.J. KRAS: The Critical Driver and Therapeutic Target for Pancreatic Cancer. Cold Spring Harb. Perspect. Med. 2018, 8, a031435. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Middleton, J.; Jeon, Y.J.; Magee, P.; Veneziano, D.; Laganà, A.; Leong, H.S.; Sahoo, S.; Fassan, M.; Booton, R.; et al. KRAS induces lung tumorigenesis through microRNAs modulation. Cell Death Dis. 2018, 9, 219. [Google Scholar] [CrossRef]
- Buscail, L.; Bournet, B.; Cordelier, P. Role of oncogenic KRAS in the diagnosis, prognosis and treatment of pancreatic cancer. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 153–168. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Haigis, K.M. KRAS Alleles: The Devil Is in the Detail. Trends Cancer 2017, 3, 686–697. [Google Scholar] [CrossRef]
- Wang, M.T.; Fer, N.; Galeas, J.; Collisson, E.A.; Kim, S.E.; Sharib, J.; McCormick, F. Blockade of leukemia inhibitory factor as a therapeutic approach to KRAS driven pancreatic cancer. Nat. Commun. 2019, 10, 3055. [Google Scholar] [CrossRef]
- Drosten, M.; Barbacid, M. Targeting the MAPK Pathway in KRAS-Driven Tumors. Cancer Cell 2020, 37, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Shui, B.; La Rocca, G.; Ventura, A.; Haigis, K.M. Interplay between K-RAS and miRNAs. Trends Cancer 2022, 8, 384–396. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, A.J.; Hoshino, D.; Hong, N.H.; Cha, D.J.; Franklin, J.L.; Coffey, R.J.; Patton, J.G.; Weaver, A.M. KRAS-MEK Signaling Controls Ago2 Sorting into Exosomes. Cell Rep. 2016, 15, 978–987. [Google Scholar] [CrossRef]
- Eulalio, A.; Behm-Ansmant, I.; Schweizer, D.; Izaurralde, E. P-body formation is a consequence, not the cause, of RNA-mediated gene silencing. Mol. Cell Biol. 2007, 27, 3970–3981. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Schmich, F.; Srivatsa, S.; Weidner, J.; Beerenwinkel, N.; Spang, A. Context-dependent deposition and regulation of mRNAs in P-bodies. Elife 2018, 7, e29815. [Google Scholar] [CrossRef]
- Zhang, B.; Herman, P.K. It is all about the process(ing): P-body granules and the regulation of signal transduction. Curr. Genet. 2020, 66, 73–77. [Google Scholar] [CrossRef]
- Luo, Y.; Na, Z.; Slavoff, S.A. P-Bodies: Composition, Properties, and Functions. Biochemistry 2018, 57, 2424–2431. [Google Scholar] [CrossRef]
- Nakajima, E.C.; Drezner, N.; Li, X.; Mishra-Kalyani, P.S.; Liu, Y.; Zhao, H.; Bi, Y.; Liu, J.; Rahman, A.; Wearne, E.; et al. FDA Approval Summary: Sotorasib for KRAS G12C-Mutated Metastatic NSCLC. Clin. Cancer Res. 2022, 28, 1482–1486. [Google Scholar] [CrossRef]
- Dhillon, S. Adagrasib: First Approval. Drugs 2023, 83, 275–285. [Google Scholar] [CrossRef]
- Huang, L.; Guo, Z.; Wang, F.; Fu, L. KRAS mutation: From undruggable to druggable in cancer. Signal. Transduct. Target. Ther. 2021, 6, 386. [Google Scholar] [CrossRef]
- Goonetilleke, K.S.; Siriwardena, A.K. Systematic review of carbohydrate antigen (CA 19-9) as a biochemical marker in the diagnosis of pancreatic cancer. Eur. J. Surg. Oncol. 2007, 33, 266–270. [Google Scholar] [CrossRef]
- Kabiraj, L.; Kundu, A. Potential role of microRNAs in pancreatic cancer manifestation: A review. J. Egypt. Natl. Cancer Inst. 2022, 34, 26. [Google Scholar] [CrossRef]
- Qi, Z.H.; Xu, H.X.; Zhang, S.R.; Xu, J.Z.; Li, S.; Gao, H.L.; Jin, W.; Wang, W.Q.; Wu, C.T.; Ni, Q.X.; et al. The Significance of Liquid Biopsy in Pancreatic Cancer. J. Cancer 2018, 9, 3417–3426. [Google Scholar] [CrossRef] [PubMed]
- Ganepola, G.A.; Rutledge, J.R.; Suman, P.; Yiengpruksawan, A.; Chang, D.H. Novel blood-based microRNA biomarker panel for early diagnosis of pancreatic cancer. World J. Gastrointest. Oncol. 2014, 6, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Schultz, N.A.; Dehlendorff, C.; Jensen, B.V.; Bjerregaard, J.K.; Nielsen, K.R.; Bojesen, S.E.; Calatayud, D.; Nielsen, S.E.; Yilmaz, M.; Holländer, N.H.; et al. MicroRNA biomarkers in whole blood for detection of pancreatic cancer. JAMA 2014, 311, 392–404. [Google Scholar] [CrossRef]
- Sadri, F.; Hosseini, S.F.; Aghayei, A.; Fereidouni, M.; Rezaei, Z. The Tumor Suppressor Roles and Mechanisms of MiR-491 in Human Cancers. DNA Cell Biol. 2022, 41, 810–823. [Google Scholar] [CrossRef]
- Zhao, G.; Zhang, J.G.; Shi, Y.; Qin, Q.; Liu, Y.; Wang, B.; Tian, K.; Deng, S.C.; Li, X.; Zhu, S.; et al. MiR-130b is a prognostic marker and inhibits cell proliferation and invasion in pancreatic cancer through targeting STAT3. PLoS ONE 2013, 8, e73803. [Google Scholar]
- Mardin, W.A.; Mees, S.T. MicroRNAs: Novel diagnostic and therapeutic tools for pancreatic ductal adenocarcinoma? Ann. Surg. Oncol. 2009, 16, 3183–3189. [Google Scholar] [CrossRef]
- Hanna, J.; Hossain, G.S.; Kocerha, J. The Potential for microRNA Therapeutics and Clinical Research. Front. Genet. 2019, 10, 478. [Google Scholar] [CrossRef]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The microRNA spectrum in 12 body fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef]
- Flatmark, K.; Hoye, E.; Fromm, B. microRNAs as cancer biomarkers. Scand. J. Clin. Lab. Investig. Suppl. 2016, 245, S80–S83. [Google Scholar] [CrossRef]
- Kumar, S.; Reddy, P.H. Are circulating microRNAs peripheral biomarkers for Alzheimer’s disease? Biochim. Biophys. Acta 2016, 1862, 1617–1627. [Google Scholar] [CrossRef] [PubMed]
- Sohel, M.M.H. Circulating microRNAs as biomarkers in cancer diagnosis. Life Sci. 2020, 248, 117473. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Tandon, M.; Alevizos, I.; Illei, G.G. The majority of microRNAs detectable in serum and saliva is concentrated in exosomes. PLoS ONE 2012, 7, e30679. [Google Scholar] [CrossRef] [PubMed]
- Iftikhar, H.; Carney, G.E. Evidence and potential in vivo functions for biofluid miRNAs: From expression profiling to functional testing: Potential roles of extracellular miRNAs as indicators of physiological change and as agents of intercellular information exchange. Bioessays 2016, 38, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Turchinovich, A.; Weiz, L.; Langheinz, A.; Burwinkel, B. Characterization of extracellular circulating microRNA. Nucleic Acids Res. 2011, 39, 7223–7233. [Google Scholar] [CrossRef]
- Makarova, J.A.; Shkurnikov, M.U.; Wicklein, D.; Lange, T.; Samatov, T.R.; Turchinovich, A.A.; Tonevitsky, A.G. Intracellular and extracellular microRNA: An update on localization and biological role. Prog. Histochem. Cytochem. 2016, 51, 33–49. [Google Scholar] [CrossRef]
- Kanlikilicer, P.; Rashed, M.H.; Bayraktar, R.; Mitra, R.; Ivan, C.; Aslan, B.; Zhang, X.; Filant, J.; Silva, A.M.; Rodriguez-Aguayo, C.; et al. Ubiquitous Release of Exosomal Tumor Suppressor miR-6126 from Ovarian Cancer Cells. Cancer Res. 2016, 76, 7194–7207. [Google Scholar] [CrossRef]
- Bayraktar, R.; Van Roosbroeck, K.; Calin, G.A. Cell-to-cell communication: MicroRNAs as hormones. Mol. Oncol. 2017, 11, 1673–1686. [Google Scholar] [CrossRef]
- Rashed, M.H.; Kanlikilicer, P.; Rodriguez-Aguayo, C.; Pichler, M.; Bayraktar, R.; Bayraktar, E.; Ivan, C.; Filant, J.; Silva, A.; Aslan, B.; et al. Exosomal miR-940 maintains SRC-mediated oncogenic activity in cancer cells: A possible role for exosomal disposal of tumor suppressor miRNAs. Oncotarget 2017, 8, 20145–20164. [Google Scholar] [CrossRef]
- Paškevičiūtė, M.; Petrikaitė, V. Overcoming transporter-mediated multidrug resistance in cancer: Failures and achievements of the last decades. Drug Deliv. Transl. Res. 2019, 9, 379–393. [Google Scholar] [CrossRef] [PubMed]
- Buschmann, D.; Kirchner, B.; Hermann, S.; Märte, M.; Wurmser, C.; Brandes, F.; Kotschote, S.; Bonin, M.; Steinlein, O.K.; Pfaffl, M.W.; et al. Evaluation of serum extracellular vesicle isolation methods for profiling miRNAs by next-generation sequencing. J. Extracell. Vesicles 2018, 7, 1481321. [Google Scholar] [CrossRef] [PubMed]
- Cheng, N.; Du, D.; Wang, X.; Liu, D.; Xu, W.; Luo, Y.; Lin, Y. Recent Advances in Biosensors for Detecting Cancer-Derived Exosomes. Trends Biotechnol. 2019, 37, 1236–1254. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.J.; Wei, R.; Li, F.; Liao, S.Y.; Tse, H.F. Mesenchymal stromal cell-derived exosomes in cardiac regeneration and repair. Stem Cell Rep. 2021, 16, 1662–1673. [Google Scholar] [CrossRef] [PubMed]
- Ying, W.; Gao, H.; Dos Reis, F.C.G.; Bandyopadhyay, G.; Ofrecio, J.M.; Luo, Z.; Ji, Y.; Jin, Z.; Ly, C.; Olefsky, J.M. MiR-690, an exosomal-derived miRNA from M2-polarized macrophages, improves insulin sensitivity in obese mice. Cell Metab. 2021, 33, 781–790.e5. [Google Scholar] [CrossRef]
- Thomou, T.; Mori, M.A.; Dreyfuss, J.M.; Konishi, M.; Sakaguchi, M.; Wolfrum, C.; Rao, T.N.; Winnay, J.N.; Garcia-Martin, R.; Grinspoon, S.K.; et al. Adipose-derived circulating miRNAs regulate gene expression in other tissues. Nature 2017, 542, 450–455. [Google Scholar] [CrossRef]
- Budakoti, M.; Panwar, A.S.; Molpa, D.; Singh, R.K.; Büsselberg, D.; Mishra, A.P.; Coutinho, H.D.M.; Nigam, M. Micro-RNA: The darkhorse of cancer. Cell. Signal. 2021, 83, 109995. [Google Scholar] [CrossRef]
- Kong, Y.W.; Ferland-McCollough, D.; Jackson, T.J.; Bushell, M. microRNAs in cancer management. Lancet Oncol. 2012, 13, e249–e258. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, S.; Marzolf, B.; Troisch, P.; Brightman, A.; Hu, Z.; Hood, L.E.; Galas, D.J. Circulating microRNAs, potential biomarkers for drug-induced liver injury. Proc. Natl. Acad. Sci. USA 2009, 106, 4402–4407. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Cui, Q. The relationship of human tissue microRNAs with those from body fluids. Sci. Rep. 2020, 10, 5644. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Ping, J.; Wen, D. MicroRNA-186 regulates the invasion and metastasis of bladder cancer via vascular endothelial growth factor C. Exp. Ther. Med. 2017, 14, 3253–3258. [Google Scholar] [CrossRef]
- Geva, G.A.; Gielchinsky, I.; Aviv, N.; Max, K.E.A.; Gofrit, O.N.; Gur-Wahnon, D.; Ben-Dov, I.Z. Urine cell-free microRNA as biomarkers for transitional cell carcinoma. BMC Res. Notes 2017, 10, 641. [Google Scholar] [CrossRef] [PubMed]
- Fedorko, M.; Juracek, J.; Stanik, M.; Svoboda, M.; Poprach, A.; Buchler, T.; Pacik, D.; Dolezel, J.; Slaby, O. Detection of let-7 miRNAs in urine supernatant as potential diagnostic approach in non-metastatic clear-cell renal cell carcinoma. Biochem. Med. 2017, 27, 411–417. [Google Scholar] [CrossRef]
- Butz, H.; Nofech-Mozes, R.; Ding, Q.; Khella, H.W.Z.; Szabó, P.M.; Jewett, M.; Finelli, A.; Lee, J.; Ordon, M.; Stewart, R.; et al. Exosomal MicroRNAs Are Diagnostic Biomarkers and Can Mediate Cell-Cell Communication in Renal Cell Carcinoma. Eur. Urol. Focus. 2016, 2, 210–218. [Google Scholar] [CrossRef]
- Erbes, T.; Hirschfeld, M.; Rücker, G.; Jaeger, M.; Boas, J.; Iborra, S.; Mayer, S.; Gitsch, G.; Stickeler, E. Feasibility of urinary microRNA detection in breast cancer patients and its potential as an innovative non-invasive biomarker. BMC Cancer 2015, 15, 193. [Google Scholar] [CrossRef]
- Debernardi, S.; Massat, N.J.; Radon, T.P.; Sangaralingam, A.; Banissi, A.; Ennis, D.P.; Dowe, T.; Chelala, C.; Pereira, S.P.; Kocher, H.M.; et al. Noninvasive urinary miRNA biomarkers for early detection of pancreatic adenocarcinoma. Am. J. Cancer Res. 2015, 5, 3455–3466. [Google Scholar]
- Shams, R.; Saberi, S.; Zali, M.; Sadeghi, A.; Ghafouri-Fard, S.; Aghdaei, H.A. Identification of potential microRNA panels for pancreatic cancer diagnosis using microarray datasets and bioinformatics methods. Sci. Rep. 2020, 10, 7559. [Google Scholar] [CrossRef]
- Al-Shaheri, F.N.; Alhamdani, M.S.S.; Bauer, A.S.; Giese, N.; Buchler, M.W.; Hackert, T.; Hoheisel, J.D. Blood biomarkers for differential diagnosis and early detection of pancreatic cancer. Cancer Treat. Rev. 2021, 96, 102193. [Google Scholar] [CrossRef]
- Marin, A.M.; Mattar, S.B.; Amatuzzi, R.F.; Chammas, R.; Uno, M.; Zanette, D.L.; Aoki, M.N. Plasma Exosome-Derived microRNAs as Potential Diagnostic and Prognostic Biomarkers in Brazilian Pancreatic Cancer Patients. Biomolecules 2022, 12, 769. [Google Scholar] [CrossRef] [PubMed]
- Tiriac, H.; Belleau, P.; Engle, D.D.; Plenker, D.; Deschênes, A.; Somerville, T.D.D.; Froeling, F.E.M.; Burkhart, R.A.; Denroche, R.E.; Jang, G.H.; et al. Organoid Profiling Identifies Common Responders to Chemotherapy in Pancreatic Cancer. Cancer Discov. 2018, 8, 1112–1129. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, J.; Chang, P.; LeBlanc, A.; Li, D.; Abbruzzesse, J.L.; Frazier, M.L.; Killary, A.M.; Sen, S. MicroRNAs in plasma of pancreatic ductal adenocarcinoma patients as novel blood-based biomarkers of disease. Cancer Prev. Res. 2009, 2, 807–813. [Google Scholar] [CrossRef]
- Havasi, A.; Sur, D.; Cainap, S.S.; Lungulescu, C.V.; Gavrilas, L.I.; Cainap, C.; Vlad, C.; Balacescu, O. Current and New Challenges in the Management of Pancreatic Neuroendocrine Tumors: The Role of miRNA-Based Approaches as New Reliable Biomarkers. Int. J. Mol. Sci. 2022, 23, 1109. [Google Scholar] [CrossRef]
- Yang, J.; Xu, R.; Wang, C.; Qiu, J.; Ren, B.; You, L. Early screening and diagnosis strategies of pancreatic cancer: A comprehensive review. Cancer Commun. 2021, 41, 1257–1274. [Google Scholar] [CrossRef]
- Ouyang, H.; Gore, J.; Deitz, S.; Korc, M. microRNA-10b enhances pancreatic cancer cell invasion by suppressing TIP30 expression and promoting EGF and TGF-β actions. Oncogene 2014, 33, 4664–4674. [Google Scholar] [CrossRef]
- Preis, M.; Gardner, T.B.; Gordon, S.R.; Pipas, J.M.; Mackenzie, T.A.; Klein, E.E.; Longnecker, D.S.; Gutmann, E.J.; Sempere, L.F.; Korc, M. MicroRNA-10b expression correlates with response to neoadjuvant therapy and survival in pancreatic ductal adenocarcinoma. Clin. Cancer Res. 2011, 17, 5812–5821. [Google Scholar] [CrossRef]
- Giovannetti, E.; Funel, N.; Peters, G.J.; Del Chiaro, M.; Erozenci, L.A.; Vasile, E.; Leon, L.G.; Pollina, L.E.; Groen, A.; Falcone, A.; et al. MicroRNA-21 in pancreatic cancer: Correlation with clinical outcome and pharmacologic aspects underlying its role in the modulation of gemcitabine activity. Cancer Res. 2010, 70, 4528–4538. [Google Scholar] [CrossRef]
- Asangani, I.A.; Rasheed, S.A.; Nikolova, D.A.; Leupold, J.H.; Colburn, N.H.; Post, S.; Allgayer, H. MicroRNA-21 (miR-21) post-transcriptionally downregulates tumor suppressor Pdcd4 and stimulates invasion, intravasation and metastasis in colorectal cancer. Oncogene 2008, 27, 2128–2136. [Google Scholar] [CrossRef]
- Guo, S.; Fesler, A.; Wang, H.; Ju, J. microRNA based prognostic biomarkers in pancreatic Cancer. Biomark. Res. 2018, 6, 18. [Google Scholar] [CrossRef]
- Drakaki, A.; Iliopoulos, D. MicroRNA-gene signaling pathways in pancreatic cancer. Biomed. J. 2013, 36, 200–208. [Google Scholar] [PubMed]
- Ohuchida, K.; Mizumoto, K.; Kayashima, T.; Fujita, H.; Moriyama, T.; Ohtsuka, T.; Ueda, J.; Nagai, E.; Hashizume, M.; Tanaka, M. MicroRNA expression as a predictive marker for gemcitabine response after surgical resection of pancreatic cancer. Ann. Surg. Oncol. 2011, 18, 2381–2387. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; VandenBoom, T.G., 2nd; Kong, D.; Wang, Z.; Ali, S.; Philip, P.A.; Sarkar, F.H. Up-regulation of miR-200 and let-7 by natural agents leads to the reversal of epithelial-to-mesenchymal transition in gemcitabine-resistant pancreatic cancer cells. Cancer Res. 2009, 69, 6704–6712. [Google Scholar] [CrossRef] [PubMed]
- Paik, W.H.; Song, B.J.; Kim, H.W.; Kim, H.R.; Hwang, J.H. MicroRNA-200c as a Prognostic Biomarker for Pancreatic Cancer. Korean J. Gastroenterol. 2015, 66, 215–220. [Google Scholar] [CrossRef]
- Yang, X.L.; Ma, Y.S.; Liu, Y.S.; Jiang, X.H.; Ding, H.; Shi, Y.; Jia, C.Y.; Lu, G.X.; Zhang, D.D.; Wang, H.M.; et al. microRNA-873 inhibits self-renewal and proliferation of pancreatic cancer stem cells through pleckstrin-2-dependent PI3K/AKT pathway. Cell Signal. 2021, 84, 110025. [Google Scholar] [CrossRef] [PubMed]
- Schultz, N.A.; Andersen, K.K.; Roslind, A.; Willenbrock, H.; Wøjdemann, M.; Johansen, J.S. Prognostic microRNAs in cancer tissue from patients operated for pancreatic cancer--five microRNAs in a prognostic index. World J. Surg. 2012, 36, 2699–2707. [Google Scholar] [CrossRef]
- Pai, P.; Rachagani, S.; Are, C.; Batra, S.K. Prospects of miRNA-based therapy for pancreatic cancer. Curr. Drug Targets 2013, 14, 1101–1109. [Google Scholar] [CrossRef]
- Liu, Y.A.; Zhang, Y.; Zheng, Z.; Li, K.; Wu, X.H.; Du, Q.G.; Ye, X.; Wang, L.; Zhu, L. MicroRNA-216b reduces growth, migration and invasion of pancreatic ductal adenocarcinoma cells by directly targeting ρ-associated coiled-coil containing protein kinase 1. Oncol. Lett. 2018, 15, 6745–6751. [Google Scholar] [CrossRef]
- Yi, J.; Li, S.; Wang, C.; Cao, N.; Qu, H.; Cheng, C.; Wang, Z.; Wang, L.; Zhou, L. Potential applications of polyphenols on main ncRNAs regulations as novel therapeutic strategy for cancer. Biomed. Pharmacother. 2019, 113, 108703. [Google Scholar] [CrossRef]
- Kang, H.; Ma, D.; Zhang, J.; Zhao, J.; Yang, M. MicroRNA-18a induces epithelial-mesenchymal transition like cancer stem cell phenotype via regulating RKIP pathway in pancreatic cancer. Ann. Transl. Med. 2020, 8, 433. [Google Scholar] [CrossRef]
- Zhu, J.; Lv, J.; Chen, J.; Zhang, X.; Ji, Y. Down-regulated microRNA-223 or elevated ZIC1 inhibits the development of pancreatic cancer via inhibiting PI3K/Akt/mTOR signaling pathway activation. Cell Cycle 2020, 19, 2851–2865. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, D.; Zhang, G.; Wu, X.; Zhou, L.; Lin, Y.; Ding, J.; An, F.; Zhan, Q. MicroRNA-23b-3p promotes pancreatic cancer cell tumorigenesis and metastasis via the JAK/PI3K and Akt/NF-kappaB signaling pathways. Oncol. Lett. 2020, 20, 160. [Google Scholar]
- Wang, W.; Wang, J.; Yang, C.; Wang, J. MicroRNA-216a targets WT1 expression and regulates KRT7 transcription to mediate the progression of pancreatic cancer-A transcriptome analysis. IUBMB Life 2021, 73, 866–882. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zheng, Y.; Chen, Q.; Dai, Y.; Li, T. MicroRNA-139 inhibits pancreatic-cancer carcinogenesis by suppressing RalB via the Ral/RAC/PI3K pathway. Arch. Biochem. Biophys. 2021, 704, 108719. [Google Scholar] [CrossRef]
- Xue, Y.; Wu, T.; Sheng, Y.; Zhong, Y.; Hu, B.; Bao, C. MicroRNA-1252-5p, regulated by Myb, inhibits invasion and epithelial-mesenchymal transition of pancreatic cancer cells by targeting NEDD9. Aging 2021, 13, 18924–18945. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Zhong, Y.; Wu, T.; Sheng, Y.; Dai, Y.; Xu, L.; Bao, C. Anti-Proliferative and Apoptosis-Promoting Effect of microRNA-125b on Pancreatic Cancer by Targeting NEDD9 via PI3K/AKT Signaling. Cancer Manag. Res. 2020, 12, 7363–7373. [Google Scholar] [CrossRef]
- Wan, X.; Guo, D.; Zhu, Q.; Qu, R. microRNA-382 suppresses the progression of pancreatic cancer through the PI3K/Akt signaling pathway by inhibition of Anxa3. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 319, G309–G322. [Google Scholar] [CrossRef]
- Nong, K.; Zhang, D.; Chen, C.; Yang, Y.; Yang, Y.; Liu, S.; Cai, H. MicroRNA-519 inhibits hypoxia-induced tumorigenesis of pancreatic cancer by regulating immune checkpoint PD-L1. Oncol. Lett. 2020, 19, 1427–1433. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Zubair, H.; Srivastava, S.K.; Singh, S.; Singh, A.P. Insights into the Role of microRNAs in Pancreatic Cancer Pathogenesis: Potential for Diagnosis, Prognosis, and Therapy. Adv. Exp. Med. Biol. 2015, 889, 71–87. [Google Scholar]
- Mikamori, M.; Yamada, D.; Eguchi, H.; Hasegawa, S.; Kishimoto, T.; Tomimaru, Y.; Asaoka, T.; Noda, T.; Wada, H.; Kawamoto, K.; et al. MicroRNA-155 Controls Exosome Synthesis and Promotes Gemcitabine Resistance in Pancreatic Ductal Adenocarcinoma. Sci. Rep. 2017, 7, 42339. [Google Scholar] [CrossRef]
- Greither, T.; Grochola, L.F.; Udelnow, A.; Lautenschlager, C.; Wurl, P.; Taubert, H. Elevated expression of microRNAs 155, 203, 210 and 222 in pancreatic tumors is associated with poorer survival. Int. J. Cancer 2010, 126, 73–80. [Google Scholar] [CrossRef]
- Papaconstantinou, I.G.; Manta, A.; Gazouli, M.; Lyberopoulou, A.; Lykoudis, P.M.; Polymeneas, G.; Voros, D. Expression of microRNAs in patients with pancreatic cancer and its prognostic significance. Pancreas 2013, 42, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Chen, H.; Wang, L.; Wang, F.; Wang, P.; Ning, Z.; Li, Y.; Liu, L.; Chen, Z.; Meng, Z. Low serum miR-373 predicts poor prognosis in patients with pancreatic cancer. Cancer Biomark. 2017, 20, 95–100. [Google Scholar] [CrossRef]
- Yuan, W.; Tang, W.; Xie, Y.; Wang, S.; Chen, Y.; Qi, J.; Qiao, Y.; Ma, J. New combined microRNA and protein plasmatic biomarker panel for pancreatic cancer. Oncotarget 2016, 7, 80033–80045. [Google Scholar] [CrossRef]
- de Gonzalo-Calvo, D.; Pérez-Boza, J.; Curado, J.; Devaux, Y. Challenges of microRNA-based biomarkers in clinical application for cardiovascular diseases. Clin. Transl. Med. 2022, 12, e585. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Ma, K.; Hao, W.; Jiao, Y.; Li, P.; Chen, J.; Xu, C.; Xu, F.J.; Lau, W.B.; Du, J.; et al. mir15a/mir16-1 cluster and its novel targeting molecules negatively regulate cardiac hypertrophy. Clin. Transl. Med. 2020, 10, e242. [Google Scholar] [CrossRef]
- Ouyang, T.; Liu, Z.; Han, Z.; Ge, Q. MicroRNA Detection Specificity: Recent Advances and Future Perspective. Anal. Chem. 2019, 91, 3179–3186. [Google Scholar] [CrossRef]
- Urbanek, M.O.; Nawrocka, A.U.; Krzyzosiak, W.J. Small RNA Detection by in Situ Hybridization Methods. Int. J. Mol. Sci. 2015, 16, 13259–13286. [Google Scholar] [CrossRef]
- Tiberio, P.; Callari, M.; Angeloni, V.; Daidone, M.G.; Appierto, V. Challenges in Using Circulating miRNAs as Cancer Biomarkers. BioMed. Res. Int. 2015, 2015, 731479. [Google Scholar] [CrossRef]
- Godoy, P.M.; Bhakta, N.R.; Barczak, A.J.; Cakmak, H.; Fisher, S.; MacKenzie, T.C.; Patel, T.; Price, R.W.; Smith, J.F.; Woodruff, P.G.; et al. Large Differences in Small RNA Composition Between Human Biofluids. Cell Rep. 2018, 25, 1346–1358. [Google Scholar] [CrossRef] [PubMed]
- Politz, J.C.; Zhang, F.; Pederson, T. MicroRNA-206 colocalizes with ribosome-rich regions in both the nucleolus and cytoplasm of rat myogenic cells. Proc. Natl. Acad. Sci. USA 2006, 103, 18957–18962. [Google Scholar] [CrossRef]
- Sweetman, D.; Rathjen, T.; Jefferson, M.; Wheeler, G.; Smith, T.G.; Wheeler, G.N.; Münsterberg, A.; Dalmay, T. FGF-4 signaling is involved in mir-206 expression in developing somites of chicken embryos. Dev. Dyn. 2006, 235, 2185–2191. [Google Scholar] [CrossRef]
- Tuddenham, L.; Wheeler, G.; Ntounia-Fousara, S.; Waters, J.; Hajihosseini, M.K.; Clark, I.; Dalmay, T. The cartilage specific microRNA-140 targets histone deacetylase 4 in mouse cells. FEBS Lett. 2006, 580, 4214–4217. [Google Scholar] [CrossRef] [PubMed]
- Beyret, E.; Lin, H. Pinpointing the expression of piRNAs and function of the PIWI protein subfamily during spermatogenesis in the mouse. Dev. Biol. 2011, 355, 215–226. [Google Scholar] [CrossRef]
- Song, R.; Ro, S.; Michaels, J.D.; Park, C.; McCarrey, J.R.; Yan, W. Many X-linked microRNAs escape meiotic sex chromosome inactivation. Nat. Genet. 2009, 41, 488–493. [Google Scholar] [CrossRef]
- Huan, T.; Chen, G.; Liu, C.; Bhattacharya, A.; Rong, J.; Chen, B.H.; Seshadri, S.; Tanriverdi, K.; Freedman, J.E.; Larson, M.G.; et al. Age-associated microRNA expression in human peripheral blood is associated with all-cause mortality and age-related traits. Aging Cell 2018, 17, e12687. [Google Scholar] [CrossRef] [PubMed]
- Ong, J.; Woldhuis, R.R.; Boudewijn, I.M.; van den Berg, A.; Kluiver, J.; Kok, K.; Terpstra, M.M.; Guryev, V.; de Vries, M.; Vermeulen, C.J.; et al. Age-related gene and miRNA expression changes in airways of healthy individuals. Sci. Rep. 2019, 9, 3765. [Google Scholar] [CrossRef]
- Chandrasekar, V.; Dreyer, J.L. microRNAs miR-124, let-7d and miR-181a regulate cocaine-induced plasticity. Mol. Cell Neurosci. 2009, 42, 350–362. [Google Scholar] [CrossRef]
- Hollander, J.A.; Im, H.I.; Amelio, A.L.; Kocerha, J.; Bali, P.; Lu, Q.; Willoughby, D.; Wahlestedt, C.; Conkright, M.D.; Kenny, P.J. Striatal microRNA controls cocaine intake through CREB signalling. Nature 2010, 466, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, A.D.; Yelamanchili, S.V.; Marcondes, M.C.; Fox, H.S. Up-regulation of microRNA-142 in simian immunodeficiency virus encephalitis leads to repression of sirtuin1. FASEB J. 2013, 27, 3720–3729. [Google Scholar] [CrossRef]
- Kong, W.; Yang, H.; He, L.; Zhao, J.J.; Coppola, D.; Dalton, W.S.; Cheng, J.Q. MicroRNA-155 is regulated by the transforming growth factor beta/Smad pathway and contributes to epithelial cell plasticity by targeting RhoA. Mol. Cell Biol. 2008, 28, 6773–6784. [Google Scholar] [CrossRef]
- Kocerha, J.; Kouri, N.; Baker, M.; Finch, N.; DeJesus-Hernandez, M.; Gonzalez, J.; Chidamparam, K.; Josephs, K.A.; Boeve, B.F.; Graff-Radford, N.R.; et al. Altered microRNA expression in frontotemporal lobar degeneration with TDP-43 pathology caused by progranulin mutations. BMC Genom. 2011, 12, 527. [Google Scholar] [CrossRef]
- Huang, C.J.; Huang, W.Y.; Chen, C.Y.; Chao, Y.J.; Chiang, N.J.; Shan, Y.S. Cancer-cell-derived cell-free DNA can predict distant metastasis earlier in pancreatic cancer: A prospective cohort study. Ther. Adv. Med. Oncol. 2022, 14, 17588359221106558. [Google Scholar] [CrossRef] [PubMed]
- Pietri, E.; Balsano, R.; Coriano, M.; Gelsomino, F.; Leonardi, F.; Bui, S.; Gnetti, L.; Valle, R.D.; Garajová, I. The implication of liquid biopsies to predict chemoresistance in pancreatic cancer. Cancer Drug Resist. 2021, 4, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Sausen, M.; Phallen, J.; Adleff, V.; Jones, S.; Leary, R.J.; Barrett, M.T.; Anagnostou, V.; Parpart-Li, S.; Murphy, D.; Kay Li, Q.; et al. Clinical implications of genomic alterations in the tumour and circulation of pancreatic cancer patients. Nat. Commun. 2015, 6, 7686. [Google Scholar] [CrossRef]
- Sugimori, M.; Sugimori, K.; Tsuchiya, H.; Suzuki, Y.; Tsuyuki, S.; Kaneta, Y.; Hirotani, A.; Sanga, K.; Tozuka, Y.; Komiyama, S.; et al. Quantitative monitoring of circulating tumor DNA in patients with advanced pancreatic cancer undergoing chemotherapy. Cancer Sci. 2020, 111, 266–278. [Google Scholar] [CrossRef] [PubMed]
- Toledano-Fonseca, M.; Cano, M.T.; Inga, E.; Rodríguez-Alonso, R.; Gómez-España, M.A.; Guil-Luna, S.; Mena-Osuna, R.; de la Haba-Rodríguez, J.R.; Rodríguez-Ariza, A.; Aranda, E. Circulating Cell-Free DNA-Based Liquid Biopsy Markers for the Non-Invasive Prognosis and Monitoring of Metastatic Pancreatic Cancer. Cancers 2020, 12, 1754. [Google Scholar] [CrossRef]
- Uesato, Y.; Sasahira, N.; Ozaka, M.; Sasaki, T.; Takatsuki, M.; Zembutsu, H. Evaluation of circulating tumor DNA as a biomarker in pancreatic cancer with liver metastasis. PLoS ONE 2020, 15, e0235623. [Google Scholar] [CrossRef]
- Wei, T.; Zhang, Q.; Li, X.; Su, W.; Li, G.; Ma, T.; Gao, S.; Lou, J.; Que, R.; Zheng, L.; et al. Monitoring Tumor Burden in Response to FOLFIRINOX Chemotherapy Via Profiling Circulating Cell-Free DNA in Pancreatic Cancer. Mol. Cancer Ther. 2019, 18, 196–203. [Google Scholar] [CrossRef]
- Koshiol, J.; Wang, E.; Zhao, Y.; Marincola, F.; Landi, M.T. Strengths and limitations of laboratory procedures for microRNA detection. Cancer Epidemiol. Biomark. Prev. 2010, 19, 907–911. [Google Scholar] [CrossRef]
- Wong, R.K.Y.; MacMahon, M.; Woodside, J.V.; Simpson, D.A. A comparison of RNA extraction and sequencing protocols for detection of small RNAs in plasma. BMC Genom. 2019, 20, 446. [Google Scholar] [CrossRef]
- Giraldez, M.D.; Spengler, R.M.; Etheridge, A.; Godoy, P.M.; Barczak, A.J.; Srinivasan, S.; De Hoff, P.L.; Tanriverdi, K.; Courtright, A.; Lu, S.; et al. Comprehensive multi-center assessment of small RNA-seq methods for quantitative miRNA profiling. Nat. Biotechnol. 2018, 36, 746–757. [Google Scholar] [CrossRef] [PubMed]
- Leshkowitz, D.; Horn-Saban, S.; Parmet, Y.; Feldmesser, E. Differences in microRNA detection levels are technology and sequence dependent. RNA 2013, 19, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Washietl, S.; Will, S.; Hendrix, D.A.; Goff, L.A.; Rinn, J.L.; Berger, B.; Kellis, M. Computational analysis of noncoding RNAs. Wiley Interdiscip. Rev. RNA 2012, 3, 759–778. [Google Scholar] [CrossRef] [PubMed]
- Godoy, P.M.; Barczak, A.J.; DeHoff, P.; Srinivasan, S.; Etheridge, A.; Galas, D.; Das, S.; Erle, D.J.; Laurent, L.C. Comparison of Reproducibility, Accuracy, Sensitivity, and Specificity of miRNA Quantification Platforms. Cell Rep. 2019, 29, 4212–4222.e5. [Google Scholar] [CrossRef]
- Yaylak, B.; Akgül, B. Experimental MicroRNA Detection Methods. In miRNomics: MicroRNA Biology and Computational Analysis; Allmer, J., Yousef, M., Eds.; Springer: New York, NY, USA, 2022; pp. 33–55. [Google Scholar]
- Jet, T.; Gines, G.; Rondelez, Y.; Taly, V. Advances in multiplexed techniques for the detection and quantification of microRNAs. Chem. Soc. Rev. 2021, 50, 4141–4161. [Google Scholar] [CrossRef]
- de Gonzalo-Calvo, D.; Martínez-Camblor, P.; Bär, C.; Duarte, K.; Girerd, N.; Fellström, B.; Schmieder, R.E.; Jardine, A.G.; Massy, Z.A.; Holdaas, H.; et al. Improved cardiovascular risk prediction in patients with end-stage renal disease on hemodialysis using machine learning modeling and circulating microribonucleic acids. Theranostics 2020, 10, 8665–8676. [Google Scholar] [CrossRef]
- Brown, C.; Mantzaris, M.; Nicolaou, E.; Karanasiou, G.; Papageorgiou, E.; Curigliano, G.; Cardinale, D.; Filippatos, G.; Memos, N.; Naka, K.K.; et al. A systematic review of miRNAs as biomarkers for chemotherapy-induced cardiotoxicity in breast cancer patients reveals potentially clinically informative panels as well as key challenges in miRNA research. Cardio-Oncology 2022, 8, 16. [Google Scholar] [CrossRef]
- Roldo, C.; Missiaglia, E.; Hagan, J.P.; Falconi, M.; Capelli, P.; Bersani, S.; Calin, G.A.; Volinia, S.; Liu, C.G.; Scarpa, A.; et al. MicroRNA expression abnormalities in pancreatic endocrine and acinar tumors are associated with distinctive pathologic features and clinical behavior. J. Clin. Oncol. 2006, 24, 4677–4684. [Google Scholar] [CrossRef]
- Zimmermann, N.; Knief, J.; Kacprowski, T.; Lazar-Karsten, P.; Keck, T.; Billmann, F.; Schmid, S.; Luley, K.; Lehnert, H.; Brabant, G.; et al. MicroRNA analysis of gastroenteropancreatic neuroendocrine tumors and metastases. Oncotarget 2018, 9, 28379–28390. [Google Scholar] [CrossRef]
- Cavalcanti, E.; Galleggiante, V.; Coletta, S.; Stasi, E.; Chieppa, M.; Armentano, R.; Serino, G. Altered miRNAs Expression Correlates With Gastroenteropancreatic Neuroendocrine Tumors Grades. Front. Oncol. 2020, 10, 1187. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Kim, H.; Kim, H.W.; Lee, J.C.; Paik, K.H.; Kang, J.; Kim, J.; Yoon, Y.S.; Han, H.S.; Sohn, I.; et al. High Expression of MicroRNA-196a Indicates Poor Prognosis in Resected Pancreatic Neuroendocrine Tumor. Medicine 2015, 94, e2224. [Google Scholar] [CrossRef] [PubMed]
- Gill, P.; Kim, E.; Chua, T.C.; Clifton-Bligh, R.J.; Nahm, C.B.; Mittal, A.; Gill, A.J.; Samra, J.S. MiRNA-3653 Is a Potential Tissue Biomarker for Increased Metastatic Risk in Pancreatic Neuroendocrine Tumours. Endocr. Pathol. 2019, 30, 128–133. [Google Scholar] [CrossRef]
- Cao, D.; Di, M.; Liang, J.; Shi, S.; Tan, Q.; Wang, Z. MicroRNA-183 in Cancer Progression. J. Cancer 2020, 11, 1315–1324. [Google Scholar] [CrossRef] [PubMed]
- Vicentini, C.; Calore, F.; Nigita, G.; Fadda, P.; Simbolo, M.; Sperandio, N.; Luchini, C.; Lawlor, R.T.; Croce, C.M.; Corbo, V.; et al. Exosomal miRNA signatures of pancreatic lesions. BMC Gastroenterol. 2020, 20, 137. [Google Scholar] [CrossRef]
- Mao, L.; Liu, S.; Hu, L.; Jia, L.; Wang, H.; Guo, M.; Chen, C.; Liu, Y.; Xu, L. miR-30 Family: A Promising Regulator in Development and Disease. Biomed. Res. Int. 2018, 2018, 9623412. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).