
Sex Differences in Glomerular Lesions, in Atherosclerosis Progression, and in the Response to Angiotensin-Converting Enzyme Inhibitors in the ApoE−/− Mice Model

 ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Blood Lipids Profile in the ApoE−/− Mice Model
2.2. Sex-Related Differences in Blood Pressure and Renal Function
2.3. Sex-Related Differences in Control and Enalapril-Treated Mice
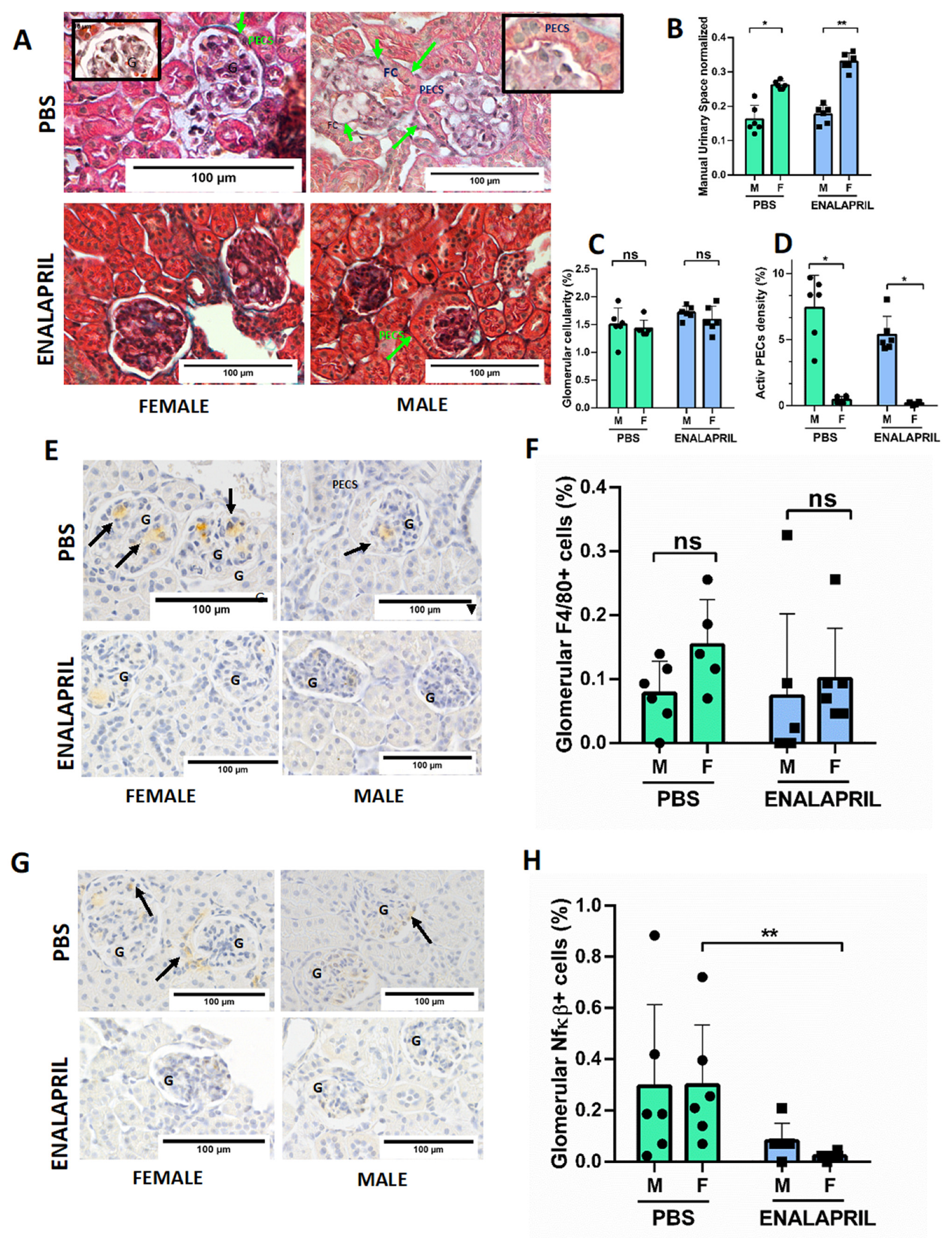
2.4. Male ApoE−/− Mice Showed Increased Number of Glomerular Foam Cells and Hypertrophy of PECs, While Enalapril Treatment Reduced Mesangial Matrix Expansion and Inflammation
3. Discussion
4. Materials and Methods
4.1. Reagents
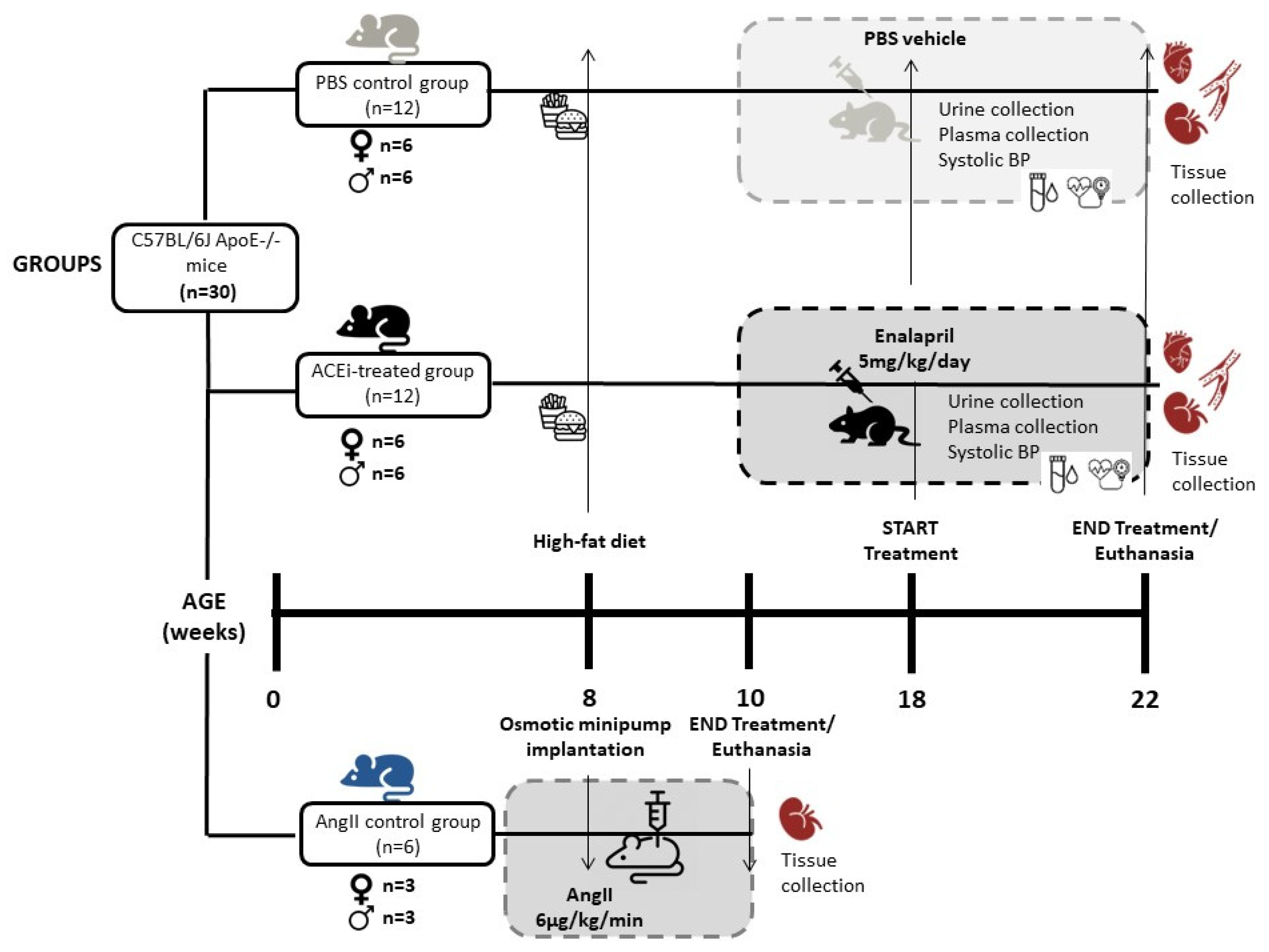
4.2. Mice and Experimental Groups
4.3. Blood Pressure Analysis
4.4. Blood and Urine Analysis
4.5. Atherosclerotic Lesions Analysis
4.6. Renal Histology Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Herrington, W.; Lacey, B.; Sherliker, P.; Armitage, J.; Lewington, S. Epidemiology of Atherosclerosis and the Potential to Reduce the Global Burden of Atherothrombotic Disease. Circ. Res. 2016, 118, 535–546. [Google Scholar] [CrossRef]
- Kon, V.; Linton, M.F.; Fazio, S. Atherosclerosis in chronic kidney disease: The role of macrophages. Nat. Rev. Nephrol. 2010, 7, 45–54. [Google Scholar] [CrossRef]
- Griffin, K.A. Hypertensive Kidney Injury and the Progression of Chronic Kidney Disease. Hypertension 2017, 70, 687–694. [Google Scholar] [CrossRef]
- Mori, T.; Cowley, A.W. Role of Pressure in Angiotensin II-Induced Renal Injury: Chronic Servo-Control of Renal Perfusion Pressure in Rats. Hypertension 2004, 43, 752–759. [Google Scholar] [CrossRef]
- Long, D.A.; Price, K.L.; Herrera-Acosta, J.; Johnson, R.J. How Does Angiotensin II Cause Renal Injury? Hypertension 2004, 43, 722–723. [Google Scholar] [CrossRef]
- Silva, G.M.; França-Falcão, M.S.; Calzerra, N.T.M.; Luz, M.S.; Gadelha, D.D.A.; Balarini, C.M.; Queiroz, T.M. Role of Renin-Angiotensin System Components in Atherosclerosis: Focus on Ang-II, ACE2, and Ang-1–7. Front. Physiol. 2020, 11, 1067. [Google Scholar] [CrossRef]
- Suganuma, E.; Zuo, Y.; Ayabe, N.; Ma, J.; Babaev, V.R.; Linton, M.F.; Fazio, S.; Ichikawa, I.; Fogo, A.B.; Kon, V. Antiatherogenic Effects of Angiotensin Receptor Antagonism in Mild Renal Dysfunction. J. Am. Soc. Nephrol. 2006, 17, 433–441. [Google Scholar] [CrossRef]
- Man, J.J.; Beckman, J.A.; Jaffe, I.Z. Sex as a Biological Variable in Atherosclerosis. Circ. Res. 2020, 126, 1297–1319. [Google Scholar] [CrossRef]
- Merz, C.N.B.; Dember, L.M.; Ingelfinger, J.R.; Vinson, A.; Neugarten, J.; Sandberg, K.L.; Sullivan, J.C.; Maric-Bilkan, C.; Rankin, T.L.; Kimmel, P.L.; et al. Star, and participants of the National Institute of Diabetes and Digestive and Kidney Diseases Workshop on “Sex and the Kidneys”. Sex and the Kidneys: Current Understanding and Research Opportunities. Nat. Rev. Nephrol. 2019, 15, 776–783. [Google Scholar] [CrossRef]
- Hilliard, L.M.; Jones, E.S.; Steckelings, U.M.; Unger, T.; Widdop, R.E.; Denton, K.M. Sex-Specific Influence of Angiotensin Type 2 Receptor Stimulation on Renal Function: A Novel Therapeutic Target for Hypertension. Hypertension 2012, 59, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Falconnet, C.; Bochud, M.; Bovet, P.; Maillard, M.; Burnier, M. Gender difference in the response to an angiotensin-converting enzyme inhibitor and a diuretic in hypertensive patients of African descent. J. Hypertens. 2004, 22, 1213–1220. [Google Scholar] [CrossRef] [PubMed]
- Hudson, M.; Rahme, E.; Behlouli, H.; Sheppard, R.; Pilote, L. Sex differences in the effectiveness of angiotensin receptor blockers and angiotensin converting enzyme inhibitors in patients with congestive heart failure—A population study. Eur. J. Heart Fail. 2007, 9, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Ruggenenti, P.; Perna, A.; Zoccali, C.; Gherardi, G.; Benini, R.; Testa, A.; Remuzzi, G. Chronic Proteinuric Nephropathies. II. Outcomes and Response to Treatment in a Prospective Cohort of 352 Patients: Differences between Women and Men in Relation to the Ace Gene Polymorphism. Gruppo Italiano Di Studi Epidemologici in Nefrologia (Gisen). J. Am. Soc. Nephrol. 2000, 11, 88–96. [Google Scholar] [CrossRef]
- Miller, J.A.; Cherney, D.Z.; Duncan, J.A.; Lai, V.; Burns, K.D.; Kennedy, C.R.; Zimpelmann, J.; Gao, W.; Cattran, D.C.; Scholey, J.W. Gender Differences in the Renal Response to Renin-Angiotensin System Blockade. J. Am. Soc. Nephrol. 2006, 17, 2554–2560. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Moeckel, G. Application of digital pathology and machine learning in the liver, kidney and lung diseases. J. Pathol. Informatics 2023, 14, 100184. [Google Scholar] [CrossRef]
- Rosenberg, A.Z.; Palmer, M.; Merlino, L.; Troost, J.P.; Gasim, A.; Bagnasco, S.; Avila-Casado, C.; Johnstone, D.; Hodgin, J.B.; Conway, C.; et al. The Application of Digital Pathology to Improve Accuracy in Glomerular Enumeration in Renal Biopsies. PLoS ONE 2016, 11, e0156441. [Google Scholar] [CrossRef] [PubMed]
- Barisoni, L.; Troost, J.P.; Nast, C.; Bagnasco, S.; Avila-Casado, C.; Hodgin, J.; Palmer, M.; Rosenberg, A.; Gasim, A.; Liensziewski, C.; et al. Reproducibility of the Neptune descriptor-based scoring system on whole-slide images and histologic and ultrastructural digital images. Mod. Pathol. 2016, 29, 671–684. [Google Scholar] [CrossRef]
- Sánchez-Jaramillo, E.A.; Gasca-Lozano, L.E.; Vera-Cruz, J.M.; Hernández-Ortega, L.D.; Salazar-Montes, A.M. Automated Computer-Assisted Image Analysis for the Fast Quantification of Kidney Fibrosis. Biology 2022, 11, 1227. [Google Scholar] [CrossRef]
- Wen, M.; Segerer, S.; Dantas, M.; Brown, P.A.; Hudkins, K.L.; Goodpaster, T.; Kirk, E.; LeBoeuf, R.C.; Alpers, C.E. Renal Injury in Apolipoprotein E–Deficient Mice. Lab. Investig. 2002, 82, 999–1006. [Google Scholar] [CrossRef]
- Belevich, I.; Jokitalo, E. DeepMIB: User-friendly and open-source software for training of deep learning network for biological image segmentation. PLoS Comput. Biol. 2021, 17, e1008374. [Google Scholar] [CrossRef]
- Lee, R.M.; Dickhout, J.G.; Sandow, S.L. Vascular structural and functional changes: Their association with causality in hypertension: Models, remodeling and relevance. Hypertens. Res. 2016, 40, 311–323. [Google Scholar] [CrossRef]
- Montecucco, F.; Pende, A.; Mach, F. The Renin-Angiotensin System Modulates Inflammatory Processes in Atherosclerosis: Evidence from Basic Research and Clinical Studies. Mediat. Inflamm. 2009, 2009, 752406. [Google Scholar] [CrossRef]
- Sata, M.; Fukuda, D. Crucial role of renin-angiotensin system in the pathogenesis of atherosclerosis. J. Med. Investig. 2010, 57, 12–25. [Google Scholar] [CrossRef]
- Vaccari, C.S.; Lerakis, S.; Hammoud, R.; Khan, B.V. Mechanisms of Benefit of Angiotensin Receptor Blockers in Coronary Atherosclerosis. Am. J. Med. Sci. 2008, 336, 270–277. [Google Scholar] [CrossRef]
- Hayashi, K.; Sasamura, H.; Azegami, T.; Itoh, H. Regression of atherosclerosis in apolipoprotein E-deficient mice is feasible using high-dose angiotensin receptor blocker, candesartan. J. Atheroscler. Thromb. 2012, 19, 736–746. [Google Scholar] [CrossRef]
- Tangirala, R.K.; Rubin, E.M.; Palinski, W. Quantitation of atherosclerosis in murine models: Correlation between lesions in the aortic origin and in the entire aorta, and differences in the extent of lesions between sexes in LDL receptor-deficient and apolipoprotein E-deficient mice. J. Lipid Res. 1995, 36, 2320–2328. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, M.E.; Polinsky, P.; Virmani, R.; Kauser, K.; Rubanyi, G.; Schwartz, S.M. Advanced Atherosclerotic Lesions in the Innominate Artery of the ApoE Knockout Mouse. Arter. Thromb. Vasc. Biol. 2000, 20, 2587–2592. [Google Scholar] [CrossRef]
- Harris, J.D.; Graham, I.R.; Schepelmann, S.; Stannard, A.K.; Roberts, M.L.; Hodges, B.L.; Hill, V.; Amalfitano, A.; Hassall, D.G.; Owen, J.S.; et al. Acute regression of advanced and retardation of early aortic atheroma in immunocompetent apolipoprotein-E (apoE) deficient mice by administration of a second generation [E1(-), E3(-), polymerase(-)] adenovirus vector expressing human apoE. Hum. Mol. Genet. 2002, 11, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Ellison, K.E.; Ingelfinger, J.R.; Pivor, M.; Dzau, V.J. Androgen regulation of rat renal angiotensinogen messenger RNA expression. J. Clin. Investig. 1989, 83, 1941–1945. [Google Scholar] [CrossRef]
- Xue, B.; Pamidimukkala, J.; Hay, M. Sex differences in the development of angiotensin II-induced hypertension in conscious mice. Am. J. Physiol.-Heart Circ. Physiol. 2005, 288, H2177–H2184. [Google Scholar] [CrossRef] [PubMed]
- Sartori-Valinotti, J.C.; Iliescu, R.; Yanes, L.L.; Dorsett-Martin, W.; Reckelhoff, J.F. Sex Differences in the Pressor Response to Angiotensin II When the Endogenous Renin-Angiotensin System Is Blocked. Hypertension 2008, 51, 1170–1176. [Google Scholar] [CrossRef] [PubMed]
- Diamond, J.R.; Karnovsky, M.J. Focal and segmental glomerulosclerosis: Analogies to atherosclerosis. Kidney Int. 1988, 33, 917–924. [Google Scholar] [CrossRef]
- Eom, M.; Hudkins, K.L.; Alpers, C.E. Foam cells and the pathogenesis of kidney disease. Curr. Opin. Nephrol. Hypertens. 2015, 24, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Ootaka, T.; Sato, H.; Furuta, T.; Sato, T.; Soma, J.; Abe, K.; Yoshinaga, K. Participation of macrophages in segmental endocapillary proliferation preceding focal glomerular sclerosis. J. Pathol. 1993, 170, 179–185. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, Y.; Chen, D.; Zeng, C.; Li, L.; Liu, Z. Presence of Foam Cells in Kidney Interstitium Is Associated with Progression of Renal Injury in Patients with Glomerular Diseases. Nephron Clin. Pract. 2009, 113, c155–c161. [Google Scholar] [CrossRef]
- Stokes, M.; Valeri, A.; Markowitz, G.; D’Agati, V. Cellular focal segmental glomerulosclerosis: Clinical and pathologic features. Kidney Int. 2006, 70, 1783–1792. [Google Scholar] [CrossRef]
- Hara, S.; Kobayashi, N.; Sakamoto, K.; Ueno, T.; Manabe, S.; Takashima, Y.; Hamada, J.; Pastan, I.; Fukamizu, A.; Matsusaka, T.; et al. Podocyte Injury–Driven Lipid Peroxidation Accelerates the Infiltration of Glomerular Foam Cells in Focal Segmental Glomerulosclerosis. Am. J. Pathol. 2015, 185, 2118–2131. [Google Scholar] [CrossRef]
- Kawaguchi, T.; Hasegawa, K.; Yasuda, I.; Muraoka, H.; Umino, H.; Tokuyama, H.; Hashiguchi, A.; Wakino, S.; Itoh, H. Diabetic condition induces hypertrophy and vacuolization in glomerular parietal epithelial cells. Sci. Rep. 2021, 11, 1515. [Google Scholar] [CrossRef] [PubMed]
- Gaut, J.P.; Hoshi, M.; Jain, S.; Liapis, H. Claudin 1 and nephrin label cellular crescents in diabetic glomerulosclerosis. Hum. Pathol. 2013, 45, 628–635. [Google Scholar] [CrossRef]
- Yadav, A.; Vallabu, S.; Kumar, D.; Ding, G.; Charney, D.N.; Chander, P.N.; Singhal, P.C. HIVAN phenotype: Consequence of epithelial mesenchymal transdifferentiation. Am. J. Physiol.-Renal Physiol. 2010, 298, F734–F744. [Google Scholar] [CrossRef] [PubMed]
- Benigni, A.; Morigi, M.; Rizzo, P.; Gagliardini, E.; Rota, C.; Abbate, M.; Ghezzi, S.; Remuzzi, A.; Remuzzi, G. Inhibiting Angiotensin-Converting Enzyme Promotes Renal Repair by Limiting Progenitor Cell Proliferation and Restoring the Glomerular Architecture. Am. J. Pathol. 2011, 179, 628–638. [Google Scholar] [CrossRef]
- Beckwith, H.; Lightstone, L.; McAdoo, S. Sex and Gender in Glomerular Disease. Semin. Nephrol. 2022, 42, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.N.; Lee, H.-W.; Osis, G.; Fang, L.; Webster, K.L.; Verlander, J.W.; Weiner, I.D. Differences in renal ammonia metabolism in male and female kidney. Am. J. Physiol.-Renal Physiol. 2018, 315, F211–F222. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.N.; Lee, H.-W.; Verlander, J.W.; Weiner, I.D. Testosterone modulates renal ammonia metabolism. Am. J. Physiol.-Renal Physiol. 2020, 318, F922–F935. [Google Scholar] [CrossRef]
- Ozluk, Y.; Blanco, P.L.; Mengel, M.; Solez, K.; Halloran, P.F.; Sis, B. Superiority of virtual microscopy versus light microscopy in transplantation pathology. Clin. Transplant. 2011, 26, 336–344. [Google Scholar] [CrossRef]
- Barisoni, L.; Nast, C.C.; Jennette, J.C.; Hodgin, J.B.; Herzenberg, A.M.; Lemley, K.V.; Conway, C.M.; Kopp, J.B.; Kretzler, M.; Lienczewski, C.; et al. Digital Pathology Evaluation in the Multicenter Nephrotic Syndrome Study Network (Neptune). Clin. J. Am. Soc. Nephrol. 2013, 8, 1449–1459. [Google Scholar] [CrossRef]
- Jha, A.; Yang, H.; Deng, R.; Kapp, M.E.; Fogo, A.B.; Huo, Y. Instance segmentation for whole slide imaging: End-to-end or detect-then-segment. J. Med. Imaging 2021, 8, 014001. [Google Scholar] [CrossRef]
- Chen, L.-C.; Papandreou, G.; Kokkinos, I.; Murphy, K.; Yuille, A.L. DeepLab: Semantic Image Segmentation with Deep Convolutional Nets, Atrous Convolution, and Fully Connected CRFs. IEEE Trans. Pattern Anal. Mach. Intell. 2018, 40, 834–848. [Google Scholar] [CrossRef]
- Hueso, M.; Mallen, A.; Ripoll, E.; de Ramon, L.; Bolaños, N.; Valera, C.; Guiteras, J.; Checa, J.; Navarro, E.; Grinyo, J.M.; et al. Cd40 Silencing by Sirna Infusion in Rodents and Evaluation by Kidney Immunostaining. Bio-Protocol 2021, 11, e4032. [Google Scholar] [CrossRef]
- Sasaki, T.; Tsuboi, N.; Haruhara, K.; Okabayashi, Y.; Kanzaki, G.; Koike, K.; Kobayashi, A.; Yamamoto, I.; Ogura, M.; Yokoo, T. Bowman Capsule Volume and Related Factors in Adults With Normal Renal Function. Kidney Int. Rep. 2018, 3, 314–320. [Google Scholar] [CrossRef]
- Su, H.; Chen, S.; He, F.-F.; Wang, Y.-M.; Bondzie, P.; Zhang, C. New Insights into Glomerular Parietal Epithelial Cell Activation and Its Signaling Pathways in Glomerular Diseases. BioMed Res. Int. 2015, 2015, 318935. [Google Scholar] [CrossRef] [PubMed]
- Herbach, N.; Schairer, I.; Blutke, A.; Kautz, S.; Siebert, A.; Göke, B.; Wolf, E.; Wanke, R.; Lieberthal, W.; Levine, J.S.; et al. Diabetic Kidney Lesions of Giprdn Transgenic Mice: Podocyte Hypertrophy and Thickening of the Gbm Precede Glomeru-lar Hypertrophy and Glomerulosclerosis. Am. J. Physiol.-Renal Physiol. 2009, 296, F819–F829. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, H.S.; Belevich, I.; Røyset, E.S.; Smistad, E.; Simpson, M.R.; Jokitalo, E.; Reinertsen, I.; Bakke, I.; Pedersen, A. Code-Free Development and Deployment of Deep Segmentation Models for Digital Pathology. Front. Med. 2022, 8, 816281. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Control | Enalapril | Enalapril | |||
|---|---|---|---|---|---|---|
| Female | Male | p | Female | Male | p | |
| n | 6 | 6 | 6 | 6 | ||
| weight (g) | 21.91 (0.58) | 30.91 (1.2) | 0.17 | 24 (3) | 31 (1.9) | 0.5 |
| LDL-cholesterol (mmol/L) | 25.96 (8.11) | 11.45 (3.96) | 0.02 | 19.92 (10.98) | 11.68 (5.17) | 0.008 |
| HDL-cholesterol (mmol/L) | 2.28 (0.45) | 2.38 (0.43) | 0.6 | 2.44 (0.13) | 2.64 (0.14) | 0.9 |
| TG (mg/dL) | 52.28 (15.89) | 84.20 (21.49) | 0.5 | 59.8 (3) | 57.7 (7.98) | 0.045 |
| Creatinine (mg/dL) | 1.1 (1.09) | 4.06 (2.1) | 0.012 | 2.04 (2.53) | 1.72 (1.46) | 0.5 |
| 24 h Proteinuria (mg/DL) | 122 (58) | 1122 (498) | 0.016 | 105 (78) | 1146 (290) | 0.037 |
| TAS (mmHg) | 137 (12) | 107 (25) | 0.2 | 108 (12) | 101 (15) | 0.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mallén, A.; Rodriguez-Urquia, R.; Alvarez, R.; Dorca-Duch, E.; Navarro, E.; Hueso, M. Sex Differences in Glomerular Lesions, in Atherosclerosis Progression, and in the Response to Angiotensin-Converting Enzyme Inhibitors in the ApoE−/− Mice Model. Int. J. Mol. Sci. 2023, 24, 13442. https://doi.org/10.3390/ijms241713442
Mallén A, Rodriguez-Urquia R, Alvarez R, Dorca-Duch E, Navarro E, Hueso M. Sex Differences in Glomerular Lesions, in Atherosclerosis Progression, and in the Response to Angiotensin-Converting Enzyme Inhibitors in the ApoE−/− Mice Model. International Journal of Molecular Sciences. 2023; 24(17):13442. https://doi.org/10.3390/ijms241713442
Chicago/Turabian StyleMallén, Adrián, Ronny Rodriguez-Urquia, Rafael Alvarez, Eduard Dorca-Duch, Estanis Navarro, and Miguel Hueso. 2023. "Sex Differences in Glomerular Lesions, in Atherosclerosis Progression, and in the Response to Angiotensin-Converting Enzyme Inhibitors in the ApoE−/− Mice Model" International Journal of Molecular Sciences 24, no. 17: 13442. https://doi.org/10.3390/ijms241713442
APA StyleMallén, A., Rodriguez-Urquia, R., Alvarez, R., Dorca-Duch, E., Navarro, E., & Hueso, M. (2023). Sex Differences in Glomerular Lesions, in Atherosclerosis Progression, and in the Response to Angiotensin-Converting Enzyme Inhibitors in the ApoE−/− Mice Model. International Journal of Molecular Sciences, 24(17), 13442. https://doi.org/10.3390/ijms241713442