
Assembling Spheroids of Rat Primary Neurons Using a Stress-Free 3D Culture System



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
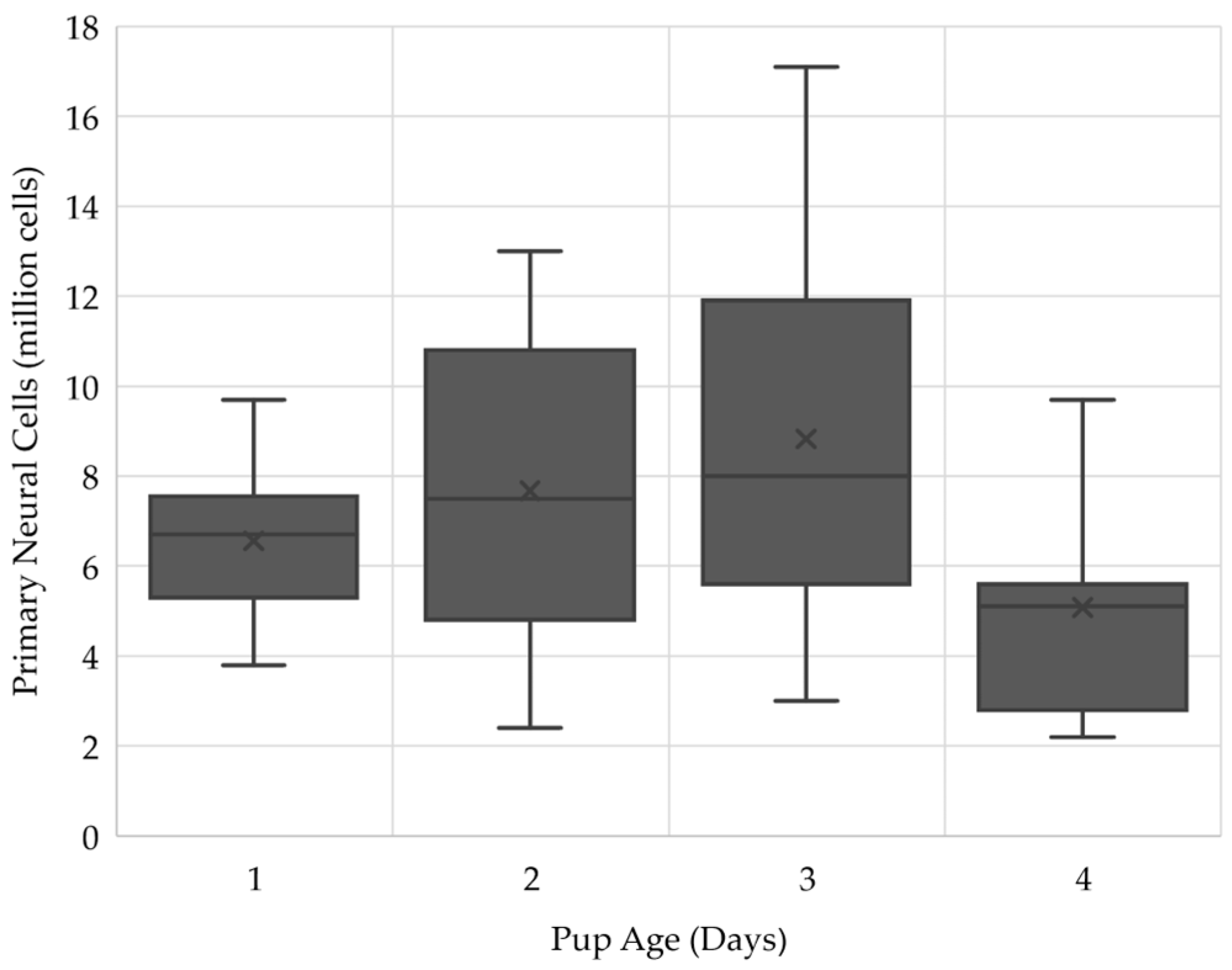
2.1. Three-Day Old Pups Provided the Largest Number of Cells for the ClinoStar
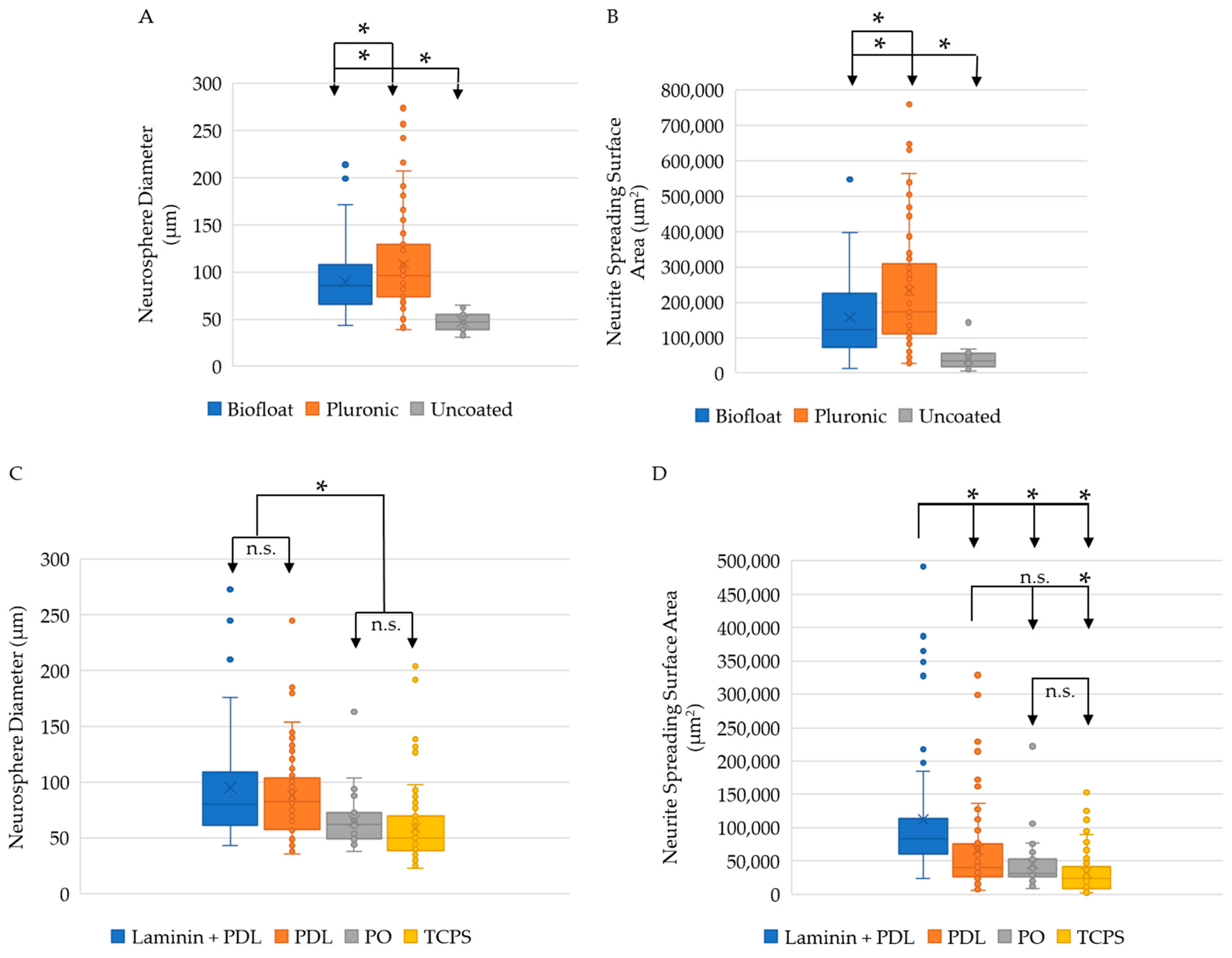
2.2. Pluronic ClinoReactor Coating and PDL Plate Coating Result in the Largest Neurosphere Size
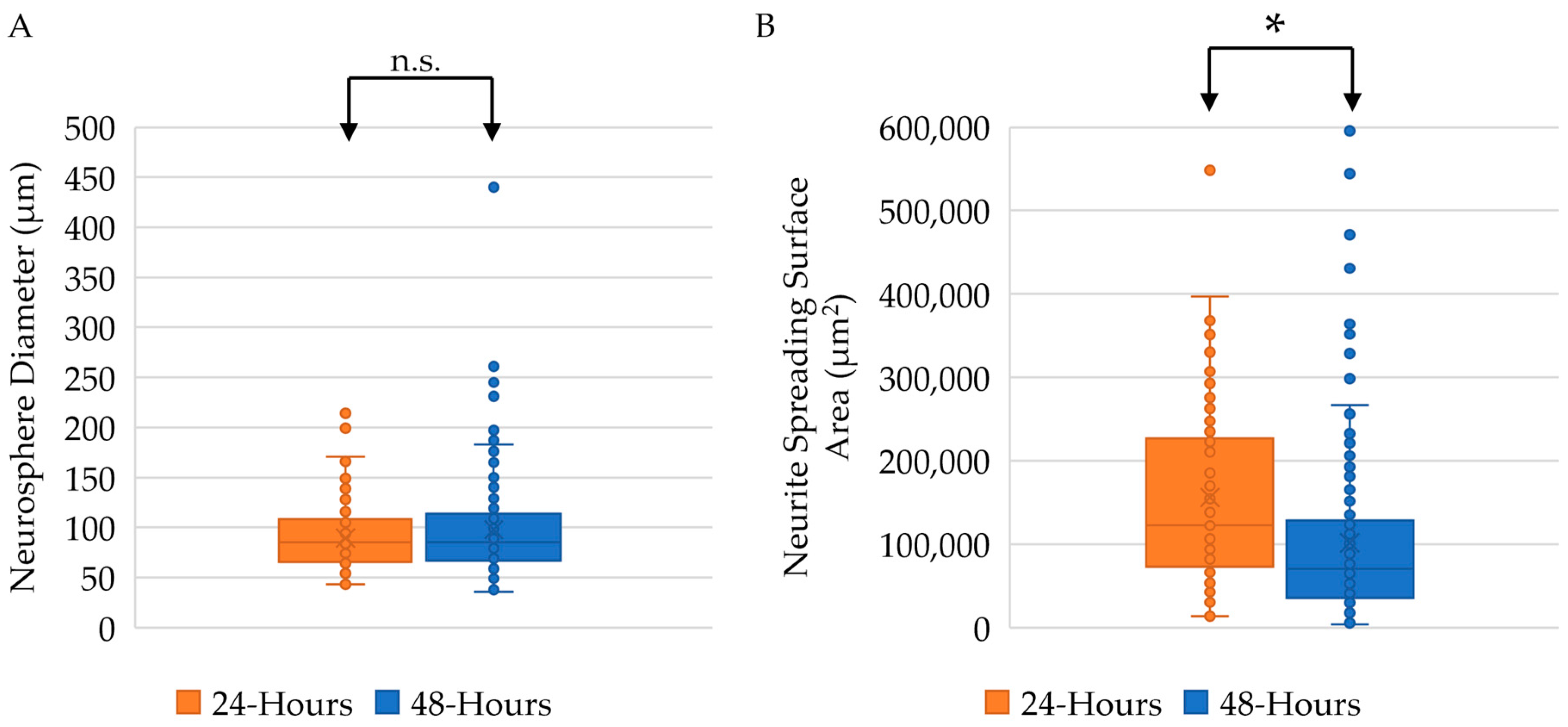
2.3. Twenty-Four-Hour Incubation Time Results in Larger Neurite Outgrowth
2.4. Supplemented Media Is Required for Appropriate Neurosphere Size and Neural Differentiation
2.5. Neurospheres Maintain Viability and Retain Neural and Glial Cell Types through 5 Days In Vitro
3. Materials and Methods
3.1. Biochemicals, Chemicals, and Disposables
3.2. Animals
3.3. Bioreactor
3.4. Primary Neural Cell Collection and Expansion
3.5. Media
3.6. Neurosphere Cultures
3.7. Viability Staining
3.8. Immunofluorescence
3.9. Image J Analyses
3.10. Statistics
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Soman, S.S.; Vijayavenkataraman, S. Perspectives on 3D Bioprinting of Peripheral Nerve Conduits. Int. J. Mol. Sci. 2020, 21, 5792. [Google Scholar] [CrossRef] [PubMed]
- Lo, J.; Chan, L.; Flynn, S. A Systematic Review of the Incidence, Prevalence, Costs, and Activity and Work Limitations of Amputation, Osteoarthritis, Rheumatoid Arthritis, Back Pain, Multiple Sclerosis, Spinal Cord Injury, Stroke, and Traumatic Brain Injury in the United States: A 2019 Update. Arch. Phys. Med. Rehabil. 2021, 102, 115–131. [Google Scholar] [CrossRef] [PubMed]
- Sulhan, S.; Lyon, K.A.; Shapiro, L.A.; Huang, J.H. Neuroinflammation and blood-brain barrier disruption following traumatic brain injury: Pathophysiology and potential therapeutic targets. J. Neurosci. Res. 2020, 98, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Erkkinen, M.G.; Kim, M.O.; Geschwind, M.D. Clinical Neurology and Epidemiology of the Major Neurodegenerative Diseases. Cold Spring Harb. Perspect. Biol. 2018, 10, a033118. [Google Scholar] [CrossRef]
- Cortesi, M.; Giordano, E. Non-destructive monitoring of 3D cell cultures: New technologies and applications. PeerJ 2022, 10, e13338. [Google Scholar] [CrossRef]
- Kim, S.J.; Kim, E.M.; Yamamoto, M.; Park, H.; Shin, H. Engineering Multi-Cellular Spheroids for Tissue Engineering and Regenerative Medicine. Adv. Healthc. Mater. 2020, 9, e2000608. [Google Scholar] [CrossRef]
- Antonova, O.Y.; Kochetkova, O.Y.; Kanev, I.L.; Shlyapnikova, E.A.; Shlyapnikov, Y.M. Rapid Generation of Neurospheres from Hippocampal Neurons Using Extracellular-Matrix-Mimetic Scaffolds. ACS Chem. Neurosci. 2021, 12, 2838–2850. [Google Scholar] [CrossRef]
- Shi, W.; Dong, P.; Kuss, M.A.; Gu, L.; Kievit, F.; Kim, H.J.; Duan, B. Design and Evaluation of an In Vitro Mild Traumatic Brain Injury Modeling System Using 3D Printed Mini Impact Device on the 3D Cultured Human iPSC Derived Neural Progenitor Cells. Adv. Healthc. Mater. 2021, 10, e2100180. [Google Scholar] [CrossRef]
- Harley-Troxell, M.E.; Dhar, M.; Anderson, D.E.; Millet, L.J. Establishing Neurospheres in vitro by a 3D Suspension Culture System and Evaluating Neurite Growth to Facilitate Nerve Repair. FASEB J. 2022, 36. [Google Scholar] [CrossRef]
- Onode, E.; Uemura, T.; Takamatsu, K.; Yokoi, T.; Shintani, K.; Hama, S.; Miyashima, Y.; Okada, M.; Nakamura, H. Bioabsorbable nerve conduits three-dimensionally coated with human induced pluripotent stem cell-derived neural stem/progenitor cells promote peripheral nerve regeneration in rats. Sci. Rep. 2021, 11, 4204. [Google Scholar] [CrossRef]
- Jahromi, M.; Razavi, S.; Bakhtiari, A. The advances in nerve tissue engineering: From fabrication of nerve conduit to in vivo nerve regeneration assays. J. Tissue Eng. Regen. Med. 2019, 13, 2077–2100. [Google Scholar] [CrossRef]
- Joseph, G.; Orme, R.P.; Kyriacou, T.; Fricker, R.A.; Roach, P. Effects of Surface Chemistry Interaction on Primary Neural Stem Cell Neurosphere Responses. ACS Omega 2021, 6, 19901–19910. [Google Scholar] [CrossRef]
- Peng, C.; Li, Y.; Lu, L.; Zhu, J.; Li, H.; Hu, J. Efficient One-Step Induction of Human Umbilical Cord-Derived Mesenchymal Stem Cells (UC-MSCs) Produces MSC-Derived Neurospheres (MSC-NS) with Unique Transcriptional Profile and Enhanced Neurogenic and Angiogenic Secretomes. Stem Cells Int. 2019, 2019, 9208173. [Google Scholar] [CrossRef]
- Peng, C.; Lu, L.; Li, Y.; Hu, J. Neurospheres Induced from Human Adipose-Derived Stem Cells as a New Source of Neural Progenitor Cells. Cell Transpl. 2019, 28, 66S–75S. [Google Scholar] [CrossRef] [PubMed]
- Decarli, M.C.; Amaral, R.; Santos, D.P.D.; Tofani, L.B.; Katayama, E.; Rezende, R.A.; Silva, J.V.L.D.; Swiech, K.; Suazo, C.A.T.; Mota, C.; et al. Cell spheroids as a versatile research platform: Formation mechanisms, high throughput production, characterization and applications. Biofabrication 2021, 13, 032002. [Google Scholar] [CrossRef]
- Xiong, F.; Gao, H.; Zhen, Y.; Chen, X.; Lin, W.; Shen, J.; Yan, Y.; Wang, X.; Liu, M.; Gao, Y. Optimal time for passaging neurospheres based on primary neural stem cell cultures. Cytotechnology 2011, 63, 621–631. [Google Scholar] [CrossRef]
- Wei, J.R.; Xiao, D.; Tang, L.; Xu, N.; Liu, R.; Shen, Y.; Xu, Z.; Sang, X.; Ge, J.; Xiang, M.; et al. Neural cell isolation from adult macaques for high-throughput analyses and neurosphere cultures. Nat. Protoc. 2023, 18, 1930–1957. [Google Scholar] [CrossRef]
- Lin, Y.J.; Lee, Y.W.; Chang, C.W.; Huang, C.C. 3D Spheroids of Umbilical Cord Blood MSC-Derived Schwann Cells Promote Peripheral Nerve Regeneration. Front. Cell Dev. Biol. 2020, 8, 604946. [Google Scholar] [CrossRef]
- Li, C.; Zhang, M.; Liu, S.Y.; Zhang, F.S.; Wan, T.; Ding, Z.T.; Zhang, P.X. Chitin Nerve Conduits with Three-Dimensional Spheroids of Mesenchymal Stem Cells from SD Rats Promote Peripheral Nerve Regeneration. Polymers 2021, 13, 3957. [Google Scholar] [CrossRef]
- He, J.; Zhang, N.; Zhu, Y.; Jin, R.; Wu, F. MSC spheroids-loaded collagen hydrogels simultaneously promote neuronal differentiation and suppress inflammatory reaction through PI3K-Akt signaling pathway. Biomaterials 2021, 265, 120448. [Google Scholar] [CrossRef]
- Oberfrank, C.; Stengel, P.; Lak, P.; Schwartz, V.; Birk, B.; Mentzel, T. The Next Generation of Low Adhesion—Increasing Flexibility and Performance. Available online: Facellitate.com (accessed on 27 July 2023).
- Chung, H.J.; Go, D.H.; Bae, J.W.; Jung, I.K.; Lee, J.W.; Park, K.D. Synthesis and characterization of Pluronic® grafted chitosan copolymer as a novel injectable biomaterial. Curr. Appl. Phys. 2005, 5, 485–488. [Google Scholar] [CrossRef]
- Kim, Y.H.; Baek, N.S.; Han, Y.H.; Chung, M.A.; Jung, S.D. Enhancement of neuronal cell adhesion by covalent binding of poly-D-lysine. J. Neurosci. Methods 2011, 202, 38–44. [Google Scholar] [CrossRef]
- Hall, P.E.; Lathia, J.D.; Caldwell, M.A.; Ffrench-Constant, C. Laminin enhances the growth of human neural stem cells in defined culture media. BMC Neurosci. 2008, 9, 71. [Google Scholar] [CrossRef]
- Millet, L.J.; Stewart, M.E.; Nuzzo, R.G.; Gillette, M.U. Guiding neuron development with planar surface gradients of substrate cues deposited using microfluidic devices. Lab Chip 2010, 10, 1525–1535. [Google Scholar] [CrossRef]
- Ge, H.; Tan, L.; Wu, P.; Yin, Y.; Liu, X.; Meng, H.; Cui, G.; Wu, N.; Lin, J.; Hu, R.; et al. Poly-L-ornithine promotes preferred differentiation of neural stem/progenitor cells via ERK signalling pathway. Sci. Rep. 2015, 5, 15535. [Google Scholar] [CrossRef]
- Fornaro, M.; Giovannelli, A.; Foggetti, A.; Muratori, L.; Geuna, S.; Novajra, G.; Perroteau, I. Role of neurotrophic factors in enhancing linear axonal growth of ganglionic sensory neurons in vitro. Neural Regen. Res. 2020, 15, 1732–1739. [Google Scholar] [CrossRef]
- Erol, O.D.; Pervin, B.; Seker, M.E.; Aerts-Kaya, F. Effects of storage media, supplements and cryopreservation methods on quality of stem cells. World J. Stem Cells 2021, 13, 1197–1214. [Google Scholar] [CrossRef]
- Hlushchuk, I.; Barut, J.; Airavaara, M.; Luk, K.; Domanskyi, A.; Chmielarz, P. Cell Culture Media, Unlike the Presence of Insulin, Affect alpha-Synuclein Aggregation in Dopaminergic Neurons. Biomolecules 2022, 12, 563. [Google Scholar] [CrossRef]
- Wang, P.Y.; Chen, J.J.; Su, H.M. Docosahexaenoic acid supplementation of primary rat hippocampal neurons attenuates the neurotoxicity induced by aggregated amyloid beta protein(42) and up-regulates cytoskeletal protein expression. J. Nutr. Biochem. 2010, 21, 345–350. [Google Scholar] [CrossRef]
- Tsui, C.T.; MacGillivray, S.R.; Weber, S.M.; McAllister, L.; Churchward, M.A.; Dennison, C.R.; Todd, K.G. Applying a novel 3D hydrogel cell culture to investigate activation of microglia due to rotational kinematics associated with mild traumatic brain injury. J. Mech. Behav. Biomed. Mater. 2021, 114, 104176. [Google Scholar] [CrossRef]
- Agalave, N.M.; Lane, B.T.; Mody, P.H.; Szabo-Pardi, T.A.; Burton, M.D. Isolation, culture, and downstream characterization of primary microglia and astrocytes from adult rodent brain and spinal cord. J. Neurosci. Methods 2020, 340, 108742. [Google Scholar] [CrossRef] [PubMed]
- Alghazali, K.M.; Newby, S.D.; Nima, Z.A.; Hamzah, R.N.; Watanabe, F.; Bourdo, S.E.; Masi, T.J.; Stephenson, S.M.; Anderson, D.E.; Dhar, M.S.; et al. Functionalized gold nanorod nanocomposite system to modulate differentiation of human mesenchymal stem cells into neural-like progenitors. Sci. Rep. 2017, 7, 16654. [Google Scholar] [CrossRef]
- Corbin, E.A.; Millet, L.J.; Keller, K.R.; King, W.P.; Bashir, R. Measuring physical properties of neuronal and glial cells with resonant microsensors. Anal. Chem. 2014, 86, 4864–4872. [Google Scholar] [CrossRef]
- Millet, L.J.; Collens, M.B.; Perry, G.L.; Bashir, R. Pattern analysis and spatial distribution of neurons in culture. Integr. Biol. 2011, 3, 1167–1178. [Google Scholar] [CrossRef]
- Millet, L.J.; Stewart, M.E.; Sweedler, J.V.; Nuzzo, R.G.; Gillette, M.U. Microfluidic devices for culturing primary mammalian neurons at low densities. Lab Chip 2007, 7, 987–994. [Google Scholar] [CrossRef]
- Newby, S.D.; Masi, T.; Griffin, C.D.; King, W.J.; Chipman, A.; Stephenson, S.; Anderson, D.E.; Biris, A.S.; Bourdo, S.E.; Dhar, M. Functionalized Graphene Nanoparticles Induce Human Mesenchymal Stem Cells to Express Distinct Extracellular Matrix Proteins Mediating Osteogenesis. Int. J. Nanomed. 2020, 15, 2501–2513. [Google Scholar] [CrossRef] [PubMed]
- Bow, A.; Newby, S.; Rifkin, R.; Jackson, B.K.; Matavosian, A.; Griffin, C.; King, W.; Alghazali, K.; Mhannawee, A.; Berryhill, S.B.; et al. Evaluation of a Polyurethane Platform for Delivery of Nanohydroxyapatite and Decellularized Bone Particles in a Porous Three-Dimensional Scaffold. ACS Appl. Bio Mater. 2019, 2, 1815–1829. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Kaminska, A.; Radoszkiewicz, K.; Rybkowska, P.; Wedzinska, A.; Sarnowska, A. Interaction of Neural Stem Cells (NSCs) and Mesenchymal Stem Cells (MSCs) as a Promising Approach in Brain Study and Nerve Regeneration. Cells 2022, 11, 1464. [Google Scholar] [CrossRef]
- Hsu, T.W.; Lu, Y.J.; Lin, Y.J.; Huang, Y.T.; Hsieh, L.H.; Wu, B.H.; Lin, Y.C.; Chen, L.C.; Wang, H.W.; Chuang, J.C.; et al. Transplantation of 3D MSC/HUVEC spheroids with neuroprotective and proangiogenic potentials ameliorates ischemic stroke brain injury. Biomaterials 2021, 272, 120765. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harley-Troxell, M.E.; Dhar, M. Assembling Spheroids of Rat Primary Neurons Using a Stress-Free 3D Culture System. Int. J. Mol. Sci. 2023, 24, 13506. https://doi.org/10.3390/ijms241713506
Harley-Troxell ME, Dhar M. Assembling Spheroids of Rat Primary Neurons Using a Stress-Free 3D Culture System. International Journal of Molecular Sciences. 2023; 24(17):13506. https://doi.org/10.3390/ijms241713506
Chicago/Turabian StyleHarley-Troxell, Meaghan E., and Madhu Dhar. 2023. "Assembling Spheroids of Rat Primary Neurons Using a Stress-Free 3D Culture System" International Journal of Molecular Sciences 24, no. 17: 13506. https://doi.org/10.3390/ijms241713506
APA StyleHarley-Troxell, M. E., & Dhar, M. (2023). Assembling Spheroids of Rat Primary Neurons Using a Stress-Free 3D Culture System. International Journal of Molecular Sciences, 24(17), 13506. https://doi.org/10.3390/ijms241713506