
Administration of Recombinant TAPBPL Protein Ameliorates Collagen-Induced Arthritis in Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. TAPBPL Protein Reduces CIA Incidence and Symptoms
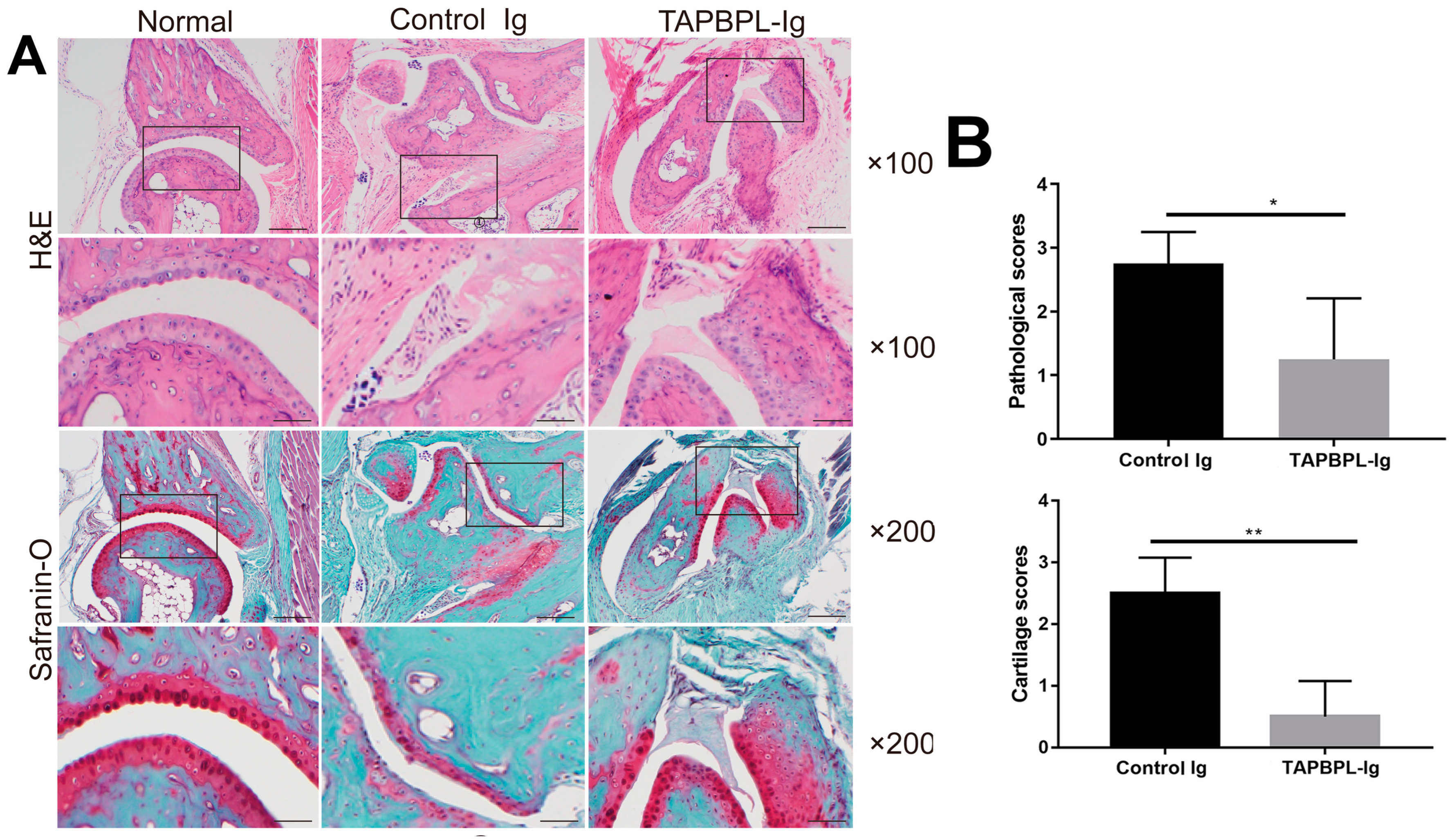
2.2. TAPBPL Protein Ameliorates CIA Pathology
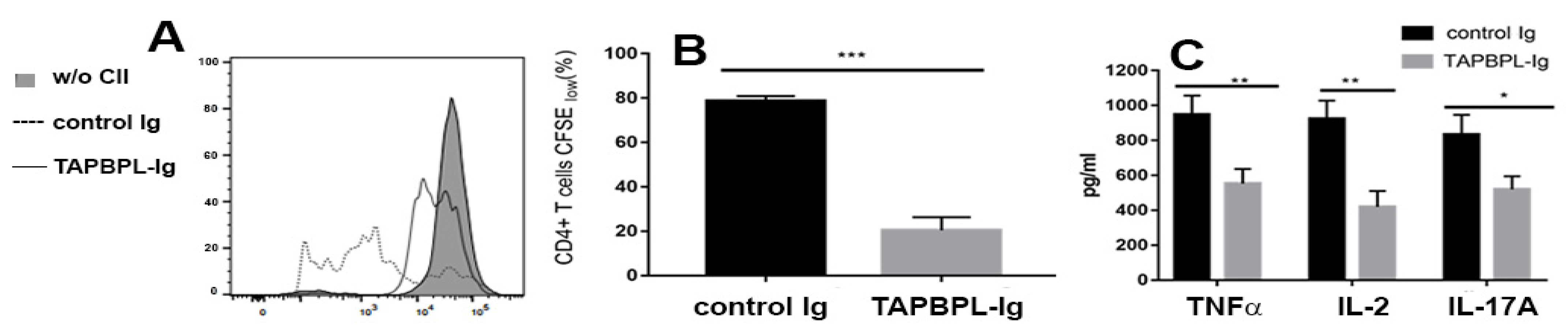
2.3. TAPBPL Protein Decreases the Production of Th1/Th17 Proinflammatory Cytokines in Joints and Serum
2.4. TAPBPL Protein Reduces the Percentage and Activation of CD4 T Cells, but Augments the Percentage of Tregs in the Spleen
2.5. TAPBPL-Ig Protein Supresses CII-Specific T Cell Proliferation and Cytokine Production
2.6. TAPBPL-Ig Protein Deminishes CII-Specific Autoantibody Production
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Production of Recombinant Human TAPBPL-Ig Fusion Protein
4.3. CIA Induction
4.4. Histological Assessment of Arthritis
4.5. CII-Specific T Cell Proliferation and Cytokine Production
4.6. Flow Cytometry Analysis
4.7. qRT-PCR
4.8. Measurement of Serum Anti-CII Antibody and Cytokine Levels
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Firestein, G.S. Evolving concepts of rheumatoid arthritis. Nature 2003, 423, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Feldmann, M.; Brennan, F.M.; Maini, R.N. Rheumatoid arthritis. Cell 1996, 85, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Smolen, J.S.; Aletaha, D.; McInnes, I.B. Rheumatoid arthritis. Lancet 2016, 388, 2023–2038. [Google Scholar] [CrossRef] [PubMed]
- Sharif, K.; Sharif, A.; Jumah, F.; Oskouian, R.; Tubbs, R.S. Rheumatoid arthritis in review: Clinical, anatomical, cellular and molecular points of view. Clin. Anat. 2018, 31, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Brand, D.D.; Latham, K.A.; Rosloniec, E.F. Collagen-induced arthritis. Nat. Protoc. 2007, 2, 1269–1275. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Zhu, G.; Xu, H.; Yao, S.; Zhou, G.; Zhu, Y.; Tamada, K.; Huang, L.; Flies, A.D.; Broadwater, M.; et al. B7-H3 Promotes Pathogenesis of Autoimmune Disease and Inflammation by Regulating the Activity of Different T Cell Subsets. PLoS ONE 2015, 10, e0130126. [Google Scholar] [CrossRef]
- Hegen, M.; Keith, J.C., Jr.; Collins, M.; Nickerson-Nutter, C.L. Utility of animal models for identification of potential therapeutics for rheumatoid arthritis. Ann. Rheum. Dis. 2008, 67, 1505–1515. [Google Scholar] [CrossRef]
- Brand, D.D.; Kang, A.H.; Rosloniec, E.F. Immunopathogenesis of collagen arthritis. Springer Semin. Immunopathol. 2003, 25, 3–18. [Google Scholar] [CrossRef]
- VanderBorght, A.; Geusens, P.; Raus, J.; Stinissen, P. The autoimmune pathogenesis of rheumatoid arthritis: Role of autoreactive T cells and new immunotherapies. Semin. Arthritis Rheum. 2001, 31, 160–175. [Google Scholar] [CrossRef]
- Cope, A.P.; Schulze-Koops, H.; Aringer, M. The central role of T cells in rheumatoid arthritis. Clin. Exp. Rheumatol. 2007, 25 (Suppl. S46), S4–S11. [Google Scholar]
- Weyand, C.M.; Bryl, E.; Goronzy, J.J. The role of T cells in rheumatoid arthritis. Arch. Immunol. Ther. Exp. 2000, 48, 429–435. [Google Scholar]
- Cui, C.; Su, M.; Lin, Y.; Lai, L. A CD300c-Fc Fusion Protein Inhibits T Cell Immunity. Front. Immunol. 2018, 9, 2657. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Cui, C.; Su, M.; Tian, X.; Huang, Y.; Zhao, J.; Lai, L. Skint8, a Novel B7 Family-Related Molecule, Negatively Regulates T Cell Responses. J. Immunol. 2019, 203, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Lin, Y.; Cui, C.; Tian, X.; Lai, L. ERMAP is a B7 family-related molecule that negatively regulates T cell and macrophage responses. Cell. Mol. Immunol. 2021, 18, 1920–1933. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Cui, C.; Su, M.; Silbart, L.K.; Liu, H.; Zhao, J.; He, L.; Huang, Y.; Xu, D.; Wei, X.; et al. Identification of TAPBPL as a novel negative regulator of T-cell function. EMBO Mol. Med. 2021, 13, e13404. [Google Scholar] [CrossRef] [PubMed]
- Feldmann, M.; Brennan, F.M.; Maini, R.N. Role of cytokines in rheumatoid arthritis. Annu. Rev. Immunol. 1996, 14, 397–440. [Google Scholar] [CrossRef]
- Müssener, A.; Litton, M.J.; Lindroos, E.; Klareskog, L. Cytokine production in synovial tissue of mice with collagen-induced arthritis (CIA). Clin. Exp. Immunol. 1997, 107, 485–493. [Google Scholar] [CrossRef]
- Iwai, H.; Kozono, Y.; Hirose, S.; Akiba, H.; Yagita, H.; Okumura, K.; Kohsaka, H.; Miyasaka, N.; Azuma, M. Amelioration of collagen-induced arthritis by blockade of inducible costimulator-B7 homologous protein costimulation. J. Immunol. 2002, 169, 4332–4339. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Yamaguchi, T.; Nomura, T.; Ono, M. Regulatory T cells and immune tolerance. Cell 2008, 133, 775–787. [Google Scholar] [CrossRef]
- Myers, L.K.; Rosloniec, E.F.; Cremer, M.A.; Kang, A.H. Collagen-induced arthritis, an animal model of autoimmunity. Life Sci. 1997, 61, 1861–1878. [Google Scholar] [CrossRef]
- Bishop, G.A.; Hostager, B.S. B lymphocyte activation by contact-mediated interactions with T lymphocytes. Curr. Opin. Immunol. 2001, 13, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ma, Y.; Liu, D.; Zhang, L.; Wei, W. The roles of B cells and their interactions with fibroblast-like synoviocytes in the pathogenesis of rheumatoid arthritis. Int. Arch. Allergy Immunol. 2011, 155, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Parker, D.C. The functions of antigen recognition in T cell-dependent B cell activation. Semin. Immunol. 1993, 5, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Parker, D.C. T cell-dependent B cell activation. Annu. Rev. Immunol. 1993, 11, 331–360. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wang, J.; Xu, Z.; Huang, F.; Qian, W.; Ma, J.; Wee, H.B.; Lewis, G.S.; June, R.R.; Schafer, P.H.; et al. Apremilast Ameliorates Experimental Arthritis via Suppression of Th1 and Th17 Cells and Enhancement of CD4+Foxp3+ Regulatory T Cells Differentiation. Front. Immunol. 2018, 9, 1662. [Google Scholar] [CrossRef]
- Han, Z.; Chang, L.; Yamanishi, Y.; Karin, M.; Firestein, G.S. Joint damage and inflammation in c-Jun N-terminal kinase 2 knockout mice with passive murine collagen-induced arthritis. Arthritis Rheum. 2002, 46, 818–823. [Google Scholar] [CrossRef]
- Jin, J.; Goldschneider, I.; Lai, L. In vivo administration of the recombinant IL-7/hepatocyte growth factor beta hybrid cytokine efficiently restores thymopoiesis and naive T cell generation in lethally irradiated mice after syngeneic bone marrow transplantation. J. Immunol. 2011, 186, 1915–1922. [Google Scholar] [CrossRef]
- Lai, L.; Zhang, M.; Goldschneider, I. Recombinant IL-7/HGFbeta efficiently induces transplantable murine hematopoietic stem cells. J. Clin. Investig. 2012, 122, 3552–3562. [Google Scholar] [CrossRef]
- Lai, L.; Zhang, M.; Song, Y.; Rood, D. Recombinant IL-7/HGFbeta Hybrid Cytokine Enhances T Cell Recovery in Mice Following Allogeneic Bone Marrow Transplantation. PLoS ONE 2013, 8, e82998. [Google Scholar] [CrossRef]
- Song, Y.; Su, M.; Panchatsharam, P.; Rood, D.; Lai, L. c-Met signalling is required for efficient postnatal thymic regeneration and repair. Immunology 2015, 144, 245–253. [Google Scholar] [CrossRef]
- Yan, Y.; Su, M.; Song, Y.; Tang, Y.; Tian, C.; Rood, D.; Lai, L. Tbx1 Modulates Endodermal and Mesodermal Differentiation from Mouse Induced Pluripotent Stem cells. Stem Cells Dev. 2014, 23, 1491–1500. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Zhao, J.; Lai, K.C.; Lai, L. Administration of Recombinant TAPBPL Protein Ameliorates Collagen-Induced Arthritis in Mice. Int. J. Mol. Sci. 2023, 24, 13772. https://doi.org/10.3390/ijms241813772
Zhang Z, Zhao J, Lai KC, Lai L. Administration of Recombinant TAPBPL Protein Ameliorates Collagen-Induced Arthritis in Mice. International Journal of Molecular Sciences. 2023; 24(18):13772. https://doi.org/10.3390/ijms241813772
Chicago/Turabian StyleZhang, Zhenzhen, Jin Zhao, Kuan Chen Lai, and Laijun Lai. 2023. "Administration of Recombinant TAPBPL Protein Ameliorates Collagen-Induced Arthritis in Mice" International Journal of Molecular Sciences 24, no. 18: 13772. https://doi.org/10.3390/ijms241813772
APA StyleZhang, Z., Zhao, J., Lai, K. C., & Lai, L. (2023). Administration of Recombinant TAPBPL Protein Ameliorates Collagen-Induced Arthritis in Mice. International Journal of Molecular Sciences, 24(18), 13772. https://doi.org/10.3390/ijms241813772