
Dietary Fiber Inulin Improves Murine Imiquimod-Induced Psoriasis-like Dermatitis



Abstract
:1. Introduction
2. Results
2.1. HFD Improved Murine Imiquimod-Induced Psoriasis-like Dermatitis
2.1.1. HFD Reduced the Thickening and Total Scores of Imiquimod-Induced Dermatitis
2.1.2. HFD Reduced Acanthosis and Number of Infiltrating Inflammatory Cells
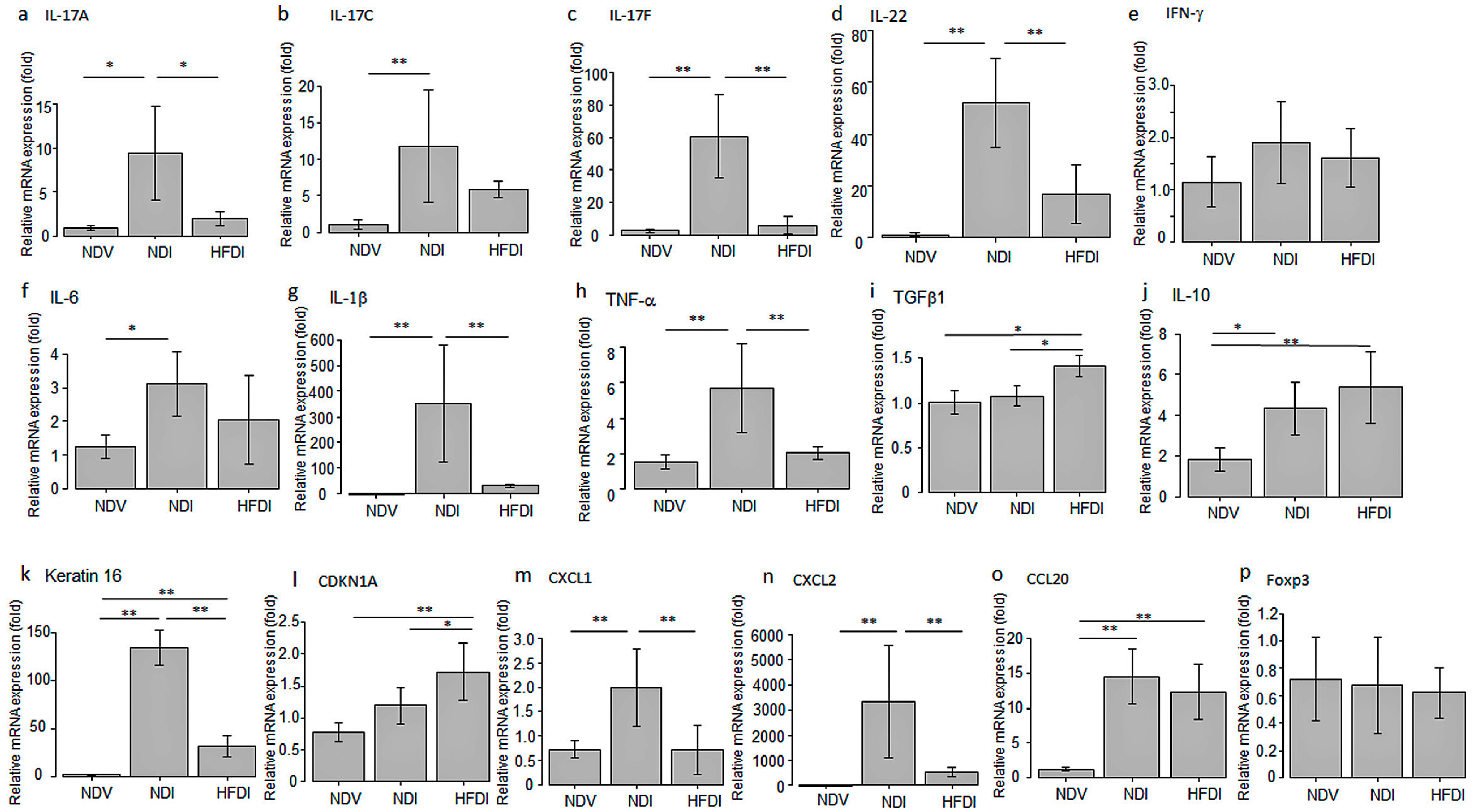
2.1.3. HFD Decreased mRNA Expression of Inflammatory Cytokines/Chemokines and Keratin 16 and Increased That of TGF-β1 and CDKN1A
2.2. Effects of Imiquimod and HFD on the Composition of the Gut Microbiome
2.3. HFD Increased Serum and Fecal Levels of Propionate
2.4. Oral Propionate Partially Attenuated Imiquimod-Induced Psoriasis-like Dermatitis
2.4.1. The Impact of Oral Propionate on Clinical and Histological Findings in Imiquimod-Induced Dermatitis
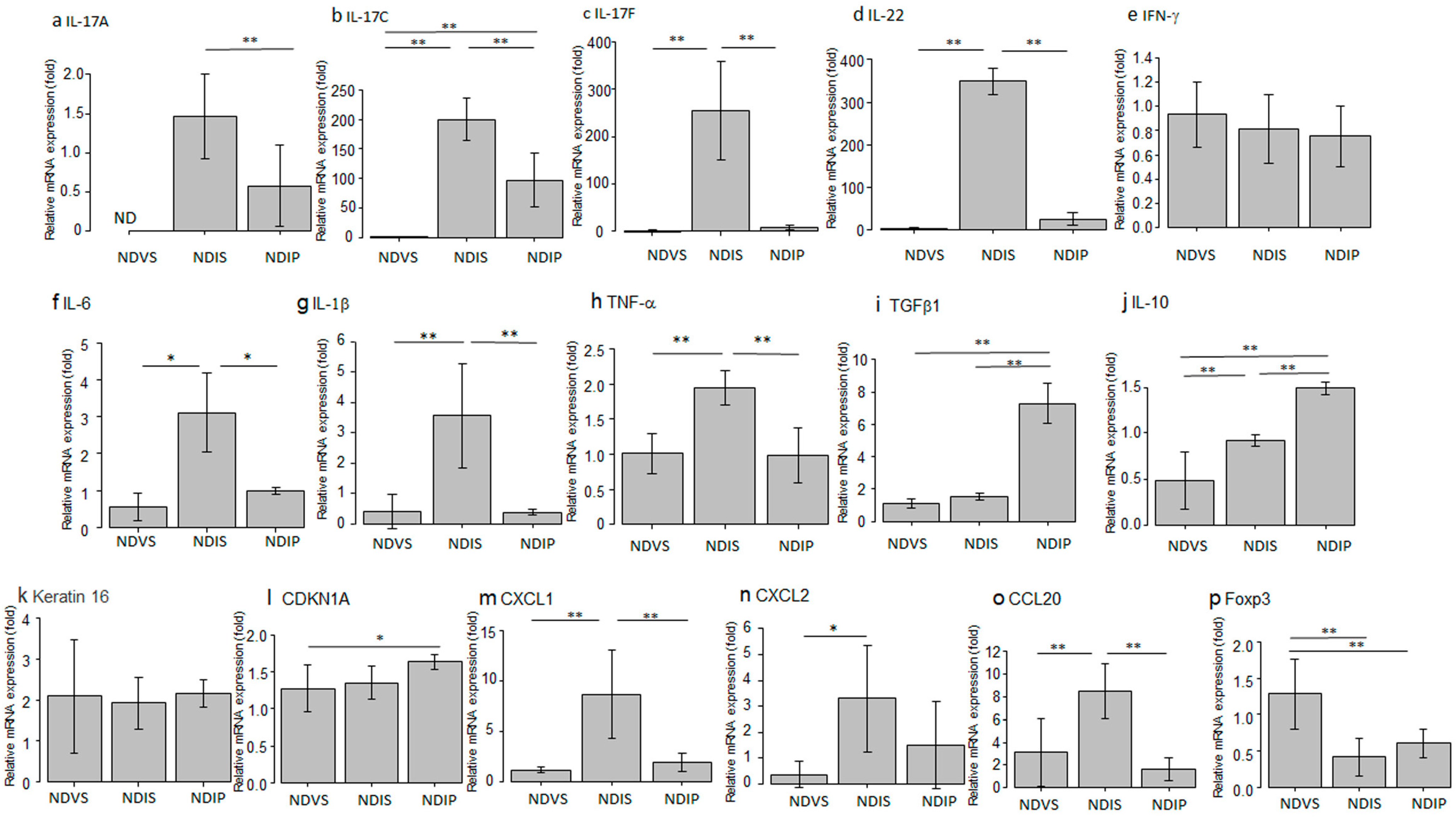
2.4.2. The Impact of Oral Propionate on mRNA Expression of Pro- or Anti-Inflammatory Cytokines/Chemokines or Proliferation-Related Molecules
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Reagents
4.3. Development of Imiquimod-Induced Psoriasis-like Dermatitis
4.4. Histological Analyses
4.5. qPCR
4.6. Measurement of the Concentrations of SCFAs in Feces and Sera
4.7. 16S rRNA Sequencing and Gut Microbial Analysis
4.8. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hu, P.; Wang, M.; Gao, H.; Zheng, A.; Li, J.; Mu, D.; Tong, J. The role of helper T cells in psoriasis. Front. Immunol. 2021, 12, 788940. [Google Scholar] [CrossRef]
- Babaie, F.; Omraninava, M.; Gorabi, A.M.; Khosrojerdi, A.; Aslani, S.; Yazdchi, A.; Torkamandi, S.; Mikaeili, H.; Sathyapalan, T.; Sahebkar, A. Etiopathogenesis of psoriasis from genetic perspective: An updated review. Curr. Genomics 2022, 23, 163–174. [Google Scholar] [PubMed]
- Yamanaka, K.; Yamamoto, O.; Honda, T. Pathophysiology of psoriasis: A review. J. Dermatol. 2021, 48, 722–731. [Google Scholar] [CrossRef] [PubMed]
- Kanda, N.; Hoashi, T.; Saeki, H. The defect in regulatory T cells in psoriasis and therapeutic approaches. J. Clin. Med. 2021, 10, 3880. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Wu, X.; Santos Rocha, C.; Rolston, M.; Garcia-Melchor, E.; Huynh, M.; Nguyen, M.; Law, T.; Haas, K.N.; Yamada, D.; et al. Short-term western diet intake promotes IL-23-mediated skin and joint inflammation accompanied by changes to the gut microbiota in mice. J. Investig. Dermatol. 2021, 141, 1780–1791. [Google Scholar] [CrossRef]
- Hidalgo-Cantabrana, C.; Gómez, J.; Delgado, S.; Requena-López, S.; Queiro-Silva, R.; Margolles, A.; Coto, E.; Sánchez, B.; Coto-Segura, P. Gut microbiota dysbiosis in a cohort of patients with psoriasis. Br. J. Dermatol. 2019, 181, 1287–1295. [Google Scholar] [CrossRef]
- Kanda, N.; Hoashi, T.; Saeki, H. Nutrition and psoriasis. Int. J. Mol. Sci. 2020, 21, 5405. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Alashkar Alhamwe, B.; Santner-Nanan, B.; Miethe, S.; Harb, H.; Renz, H.; Potaczek, D.P.; Nanan, R.K. Short-chain fatty acids augment differentiation and function of human induced regulatory T cells. Int. J. Mol. Sci. 2022, 23, 5740. [Google Scholar] [CrossRef]
- Pham, C.H.; Lee, J.E.; Yu, J.; Lee, S.H.; Yu, K.R.; Hong, J.; Cho, N.; Kim, S.; Kang, D.; Lee, S.; et al. Anticancer effects of propionic acid inducing cell death in cervical cancer cells. Molecules 2021, 26, 4951. [Google Scholar] [CrossRef]
- Roy, R.; Nguyen-Ngo, C.; Lappas, M. Short-chain fatty acids as novel therapeutics for gestational diabetes. J. Mol. Endocrinol. 2020, 65, 21–34. [Google Scholar] [CrossRef]
- Zou, F.; Qiu, Y.; Huang, Y.; Zou, H.; Cheng, X.; Niu, Q.; Luo, A.; Sun, J. Effects of short-chain fatty acids in inhibiting HDAC and activating p38 MAPK are critical for promoting B10 cell generation and function. Cell Death Dis. 2021, 12, 582. [Google Scholar] [CrossRef] [PubMed]
- Nazri, J.M.; Oikonomopoulou, K.; de Araujo, E.D.; Kraskouskaya, D.; Gunning, P.T.; Chandran, V. Histone deacetylase inhibitors as a potential new treatment for psoriatic disease and other inflammatory conditions. Crit. Rev. Clin. Lab. Sci. 2023, 60, 300–320. [Google Scholar] [CrossRef]
- Kleuskens, M.T.A.; Haasnoot, M.L.; Herpers, B.M.; Ampting, M.; Bredenoord, A.J.; Garssen, J.; Redegeld, F.A.; van Esch, B. Butyrate and propionate restore interleukin 13-compromised esophageal epithelial barrier function. Allergy 2022, 77, 1510–1521. [Google Scholar] [CrossRef] [PubMed]
- Kespohl, M.; Vachharajani, N.; Luu, M.; Harb, H.; Pautz, S.; Wolff, S.; Sillner, N.; Walker, A.; Schmitt-Kopplin, P.; Boettger, T.; et al. The microbial metabolite butyrate induces expression of Th1-associated factors in CD4(+) T Cells. Front. Immunol. 2017, 8, 1036. [Google Scholar] [CrossRef]
- Kim, C.H.; Park, J.; Kim, M. Gut microbiota-derived short-chain fatty acids, T cells, and inflammation. Immune Netw. 2014, 14, 277–288. [Google Scholar] [CrossRef]
- Valcheva, R.; Koleva, P.; Martínez, I.; Walter, J.; Gänzle, M.G.; Dieleman, L.A. Inulin-type fructans improve active ulcerative colitis associated with microbiota changes and increased short-chain fatty acids levels. Gut Microbes 2019, 10, 334–357. [Google Scholar] [CrossRef]
- Dürholz, K.; Hofmann, J.; Iljazovic, A.; Häger, J.; Lucas, S.; Sarter, K.; Strowig, T.; Bang, H.; Rech, J.; Schett, G.; et al. Dietary short-term fiber interventions in arthritis patients increase systemic SCFA levels and regulate inflammation. Nutrients 2020, 12, 3207. [Google Scholar] [CrossRef]
- Häger, J.; Bang, H.; Hagen, M.; Frech, M.; Träger, P.; Sokolova, M.V.; Steffen, U.; Tascilar, K.; Sarter, K.; Schett, G.; et al. The role of dietary fiber in rheumatoid arthritis patients: A feasibility study. Nutrients 2019, 11, 2392. [Google Scholar] [CrossRef] [PubMed]
- Haghikia, A.; Jörg, S.; Duscha, A.; Berg, J.; Manzel, A.; Waschbisch, A.; Hammer, A.; Lee, D.H.; May, C.; Wilck, N.; et al. Dietary fatty acids directly impact central nervous system autoimmunity via the small intestine. Immunity 2015, 43, 817–829. [Google Scholar] [CrossRef]
- Lewis, G.; Wang, B.; Shafiei Jahani, P.; Hurrell, B.P.; Banie, H.; Aleman Muench, G.R.; Maazi, H.; Helou, D.G.; Howard, E.; Galle-Treger, L.; et al. Dietary fiber-induced microbial short chain fatty acids suppress ILC2-dependent airway inflammation. Front. Immunol. 2019, 10, 2051. [Google Scholar] [CrossRef]
- Takahashi, M.; Takahashi, K.; Abe, S.; Yamada, K.; Suzuki, M.; Masahisa, M.; Endo, M.; Abe, K.; Inoue, R.; Hoshi, H. Improvement of psoriasis by alteration of the gut environment by oral administration of fucoidan from Cladosiphon Okamuranus. Mar. Drugs 2020, 18, 154. [Google Scholar] [CrossRef] [PubMed]
- Gangwar, R.S.; Gudjonsson, J.E.; Ward, N.L. Mouse models of psoriasis: A comprehensive review. J. Investig. Dermatol. 2022, 142 3 Pt. B, 884–897. [Google Scholar] [CrossRef]
- Then, C.K.; Paillas, S.; Wang, X.; Hampson, A.; Kiltie, A.E. Association of Bacteroides acidifaciens relative abundance with high-fibre diet-associated radiosensitisation. BMC Biol. 2020, 18, 102. [Google Scholar] [CrossRef]
- Su, X.; Yin, X.; Liu, Y.; Yan, X.; Zhang, S.; Wang, X.; Lin, Z.; Zhou, X.; Gao, J.; Wang, Z.; et al. Gut dysbiosis contributes to the imbalance of Treg and Th17 cells in Graves’ disease patients by propionic acid. J. Clin. Endocrinol. Metab. 2020, 105, 3526–3547. [Google Scholar] [CrossRef]
- Xia, Y.; Miao, J.; Zhang, Y.; Zhang, H.; Kong, L.; Seviour, R.; Kong, Y. Dietary inulin supplementation modulates the composition and activities of carbohydrate-metabolizing organisms in the cecal microbiota of broiler chickens. PLoS ONE 2021, 16, e0258663. [Google Scholar] [CrossRef]
- Shon, H.J.; Kim, Y.M.; Kim, K.S.; Choi, J.O.; Cho, S.H.; An, S.; Park, S.H.; Cho, Y.J.; Park, J.H.; Seo, S.U.; et al. Protective role of colitis in inflammatory arthritis via propionate-producing Bacteroides in the gut. Front. Immunol. 2023, 14, 1064900. [Google Scholar] [CrossRef] [PubMed]
- Han, K.H.; Tsuchihira, H.; Nakamura, Y.; Shimada, K.; Ohba, K.; Aritsuka, T.; Uchino, H.; Kikuchi, H.; Fukushima, M. Inulin-type fructans with different degrees of polymerization improve lipid metabolism but not glucose metabolism in rats fed a high-fat diet under energy restriction. Dig. Dis. Sci. 2013, 58, 2177–2186. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; Chain, F.; Martín, R.; Bermúdez-Humarán, L.G.; Courau, S.; Langella, P. Beneficial effects on host energy metabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria. Microb. Cell Fact. 2017, 16, 79. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, T.; Mori, H.; Iwata, M.; Meguro, S. Growth stimulator for bifidobacteria produced by Propionibacterium freudenreichii and several intestinal bacteria. J. Dairy. Sci. 1994, 77, 393–404. [Google Scholar] [CrossRef]
- Shapiro, J.; Cohen, N.A.; Shalev, V.; Uzan, A.; Koren, O.; Maharshak, N. Psoriatic patients have a distinct structural and functional fecal microbiota compared with controls. J. Dermatol. 2019, 46, 595–603. [Google Scholar] [CrossRef]
- Dei-Cas, I.; Giliberto, F.; Luce, L.; Dopazo, H.; Penas-Steinhardt, A. Metagenomic analysis of gut microbiota in non-treated plaque psoriasis patients stratified by disease severity: Development of a new Psoriasis-Microbiome Index. Sci. Rep. 2020, 10, 12754. [Google Scholar] [CrossRef] [PubMed]
- Codoñer, F.M.; Ramírez-Bosca, A.; Climent, E.; Carrión-Gutierrez, M.; Guerrero, M.; Pérez-Orquín, J.M.; Horga de la Parte, J.; Genovés, S.; Ramón, D.; Navarro-López, V.; et al. Gut microbial composition in patients with psoriasis. Sci. Rep. 2018, 8, 3812. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, A.; Sasaki, T.; Itoh, K.; Kitahara, T.; Takema, Y.; Hiramatsu, K.; Ishikawa, D.; Shibuya, T.; Kobayashi, O.; Osada, T.; et al. A Soluble fiber diet increases Bacteroides fragilis group abundance and immunoglobulin A production in the gut. Appl. Environ. Microbiol. 2020, 86, e00405-20. [Google Scholar] [CrossRef] [PubMed]
- Kawasoe, J.; Uchida, Y.; Kawamoto, H.; Miyauchi, T.; Watanabe, T.; Saga, K.; Tanaka, K.; Ueda, S.; Terajima, H.; Taura, K.; et al. Propionic acid, induced in gut by an inulin diet, suppresses inflammation and ameliorates liver ischemia and reperfusion injury in mice. Front. Immunol. 2022, 13, 862503. [Google Scholar] [CrossRef]
- Silva, L.G.; Ferguson, B.S.; Avila, A.S.; Faciola, A.P. Sodium propionate and sodium butyrate effects on histone deacetylase (HDAC) activity, histone acetylation, and inflammatory gene expression in bovine mammary epithelial cells. J. Anim. Sci. 2018, 96, 5244–5252. [Google Scholar] [CrossRef] [PubMed]
- Tse, C.S.; Williams, D.M. Inhibition of human endothelial cell proliferation in vitro in response to n-butyrate and propionate. J. Periodontal Res. 1992, 27, 506–510. [Google Scholar] [CrossRef]
- Leus, N.G.; Zwinderman, M.R.; Dekker, F.J. Histone deacetylase 3 (HDAC 3) as emerging drug target in NF-κB-mediated inflammation. Curr. Opin. Chem. Biol. 2016, 33, 160–168. [Google Scholar] [CrossRef]
- Place, R.F.; Noonan, E.J.; Giardina, C. HDAC inhibition prevents NF-kappa B activation by suppressing proteasome activity: Down-regulation of proteasome subunit expression stabilizes I kappa B alpha. Biochem. Pharmacol. 2005, 70, 394–406. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Martin-Gallausiaux, C.; Béguet-Crespel, F.; Marinelli, L.; Jamet, A.; Ledue, F.; Blottière, H.M.; Lapaque, N. Butyrate produced by gut commensal bacteria activates TGF-beta1 expression through the transcription factor SP1 in human intestinal epithelial cells. Sci. Rep. 2018, 8, 9742. [Google Scholar] [CrossRef]
- Pan, L.; Pan, H.; Jiang, H.; Du, J.; Wang, X.; Huang, B.; Lu, J. HDAC4 inhibits the transcriptional activation of mda-7/IL-24 induced by Sp1. Cell Mol. Immunol. 2010, 7, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Imai, K.; Okamoto, T.; Ochiai, K. Involvement of Sp1 in butyric acid-induced HIV-1 gene expression. Cell Physiol. Biochem. 2015, 37, 853–865. [Google Scholar] [CrossRef]
- Jin, H.; Kanthasamy, A.; Harischandra, D.S.; Kondru, N.; Ghosh, A.; Panicker, N.; Anantharam, V.; Rana, A.; Kanthasamy, A.G. Histone hyperacetylation up-regulates protein kinase Cδ in dopaminergic neurons to induce cell death: Relevance to epigenetic mechanisms of neurodegeneration in Parkinson disease. J. Biol. Chem. 2014, 289, 34743–34767. [Google Scholar] [CrossRef] [PubMed]
- Li, M.O.; Flavell, R.A. Contextual regulation of inflammation: A duet by transforming growth factor-beta and interleukin-10. Immunity 2008, 28, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kim, M.; Kang, S.G.; Jannasch, A.H.; Cooper, B.; Patterson, J.; Kim, C.H. Short-chain fatty acids induce both effector and regulatory T cells by suppression of histone deacetylases and regulation of the mTOR-S6K pathway. Mucosal Immunol. 2015, 8, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Wu, W.; Chen, L.; Yang, W.; Huang, X.; Ma, C.; Chen, F.; Xiao, Y.; Zhao, Y.; Ma, C.; et al. Microbiota-derived short-chain fatty acids promote Th1 cell IL-10 production to maintain intestinal homeostasis. Nat. Commun. 2018, 9, 3555. [Google Scholar] [CrossRef]
- Asquith, M.; Davin, S.; Stauffer, P.; Michell, C.; Janowitz, C.; Lin, P.; Ensign-Lewis, J.; Kinchen, J.M.; Koop, D.R.; Rosenbaum, J.T. Intestinal metabolites are profoundly altered in the context of HLA-B27 expression and functionally modulate disease in a rat model of spondyloarthritis. Arthritis Rheumatol. 2017, 69, 1984–1995. [Google Scholar] [CrossRef]
- Wu, H.; Van Der Pol, W.J.; Dubois, L.G.; Morrow, C.D.; Tollefsbol, T.O. Dietary supplementation of inulin contributes to the prevention of estrogen receptor-negative mammary cancer by alteration of gut microbial communities and epigenetic regulations. Int. J. Mol. Sci. 2023, 24, 9015. [Google Scholar] [CrossRef]
- Pöllänen, M.T.; Overman, D.O.; Salonen, J.I. Bacterial metabolites sodium butyrate and propionate inhibit epithelial cell growth in vitro. J. Periodontal Res. 1997, 32, 326–334. [Google Scholar] [CrossRef]
- Semaan, J.; El-Hakim, S.; Ibrahim, J.N.; Safi, R.; Elnar, A.A.; El Boustany, C. Comparative effect of sodium butyrate and sodium propionate on proliferation, cell cycle and apoptosis in human breast cancer cells MCF-7. Breast Cancer 2020, 27, 696–705. [Google Scholar] [CrossRef]
- Kim, K.; Kwon, O.; Ryu, T.Y.; Jung, C.R.; Kim, J.; Min, J.K.; Kim, D.S.; Son, M.Y.; Cho, H.S. Propionate of a microbiota metabolite induces cell apoptosis and cell cycle arrest in lung cancer. Mol. Med. Rep. 2019, 20, 1569–1574. [Google Scholar] [CrossRef]
- Hinnebusch, B.F.; Meng, S.; Wu, J.T.; Archer, S.Y.; Hodin, R.A. The effects of short-chain fatty acids on human colon cancer cell phenotype are associated with histone hyperacetylation. J. Nutr. 2002, 132, 1012–1017. [Google Scholar] [CrossRef]
- Park, H.S.; Han, J.H.; Park, J.W.; Lee, D.H.; Jang, K.W.; Lee, M.; Heo, K.S.; Myung, C.S. Sodium propionate exerts anticancer effect in mice bearing breast cancer cell xenograft by regulating JAK2/STAT3/ROS/p38 MAPK signaling. Acta Pharmacol. Sin. 2021, 42, 1311–1323. [Google Scholar] [CrossRef] [PubMed]
- Richon, V.M.; Sandhoff, T.W.; Rifkind, R.A.; Marks, P.A. Histone deacetylase inhibitor selectively induces p21WAF1 expression and gene-associated histone acetylation. Proc. Natl. Acad. Sci. USA 2000, 97, 10014–10019. [Google Scholar] [CrossRef] [PubMed]
- Arifuzzaman, M.; Won, T.H.; Li, T.T.; Yano, H.; Digumarthi, S.; Heras, A.F.; Zhang, W.; Parkhurst, C.N.; Kashyap, S.; Jin, W.B.; et al. Inulin fibre promotes microbiota-derived bile acids and type 2 inflammation. Nature 2022, 611, 578–584. [Google Scholar] [CrossRef]
- Wan, Y.; Yuan, J.; Li, J.; Li, H.; Zhang, J.; Tang, J.; Ni, Y.; Huang, T.; Wang, F.; Zhao, F.; et al. Unconjugated and secondary bile acid profiles in response to higher-fat, lower-carbohydrate diet and associated with related gut microbiota: A 6-month randomized controlled-feeding trial. Clin. Nutr. 2020, 39, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Wu, X.; Wu, C.Y.; Singh, S.P.; Law, T.; Yamada, D.; Huynh, M.; Liakos, W.; Yang, G.; Farber, J.M.; et al. Bile acids improve psoriasiform dermatitis through inhibition of IL-17A expression and CCL20-CCR6-mediated trafficking of T Cells. J. Investig. Dermatol. 2022, 142, 1381–1390.e11. [Google Scholar] [CrossRef]
- Vessey, D.A.; Lee, K.H.; Lau, E. Effect of bile acids on the growth and differentiation of cultured human keratinocytes. Skin. Pharmacol. 1997, 10, 265–274. [Google Scholar] [CrossRef]
- Chen, X.; Chen, Y.; Stanton, C.; Ross, R.P.; Zhao, J.; Chen, W.; Yang, B. Dose-response efficacy and mechanisms of orally administered Bifidobacterium breve CCFM683 on IMQ-induced psoriasis in mice. Nutrients 2023, 15, 1592. [Google Scholar] [CrossRef]
- Sittipo, P.; Lobionda, S.; Choi, K.; Sari, I.N.; Kwon, H.Y.; Lee, Y.K. Toll-like receptor 2-mediated suppression of colorectal cancer pathogenesis by polysaccharide A From Bacteroides fragilis. Front. Microbiol. 2018, 9, 1588. [Google Scholar] [CrossRef]
- Nakaguma, H.; Kambara, T.; Yamamoto, T. Rat ultraviolet ray B photodermatitis: An experimental model of psoriasis vulgaris. Int. J. Exp. Pathol. 1995, 76, 65–73. [Google Scholar]
- Yu, S.; Wu, X.; Zhou, Y.; Sheng, L.; Jena, P.K.; Han, D.; Wan, Y.J.Y.; Hwang, S.T. A Western Diet, but not a high-fat and low-sugar diet, predisposes mice to enhanced susceptibility to imiquimod-induced psoriasiform dermatitis. J. Investig. Dermatol. 2019, 139, 1404–1407. [Google Scholar] [CrossRef]
- Ikumi, K.; Odanaka, M.; Shime, H.; Imai, M.; Osaga, S.; Taguchi, O.; Nishida, E.; Hemmi, H.; Kaisho, T.; Morita, A.; et al. Hyperglycemia is associated with psoriatic inflammation in both humans and mice. J. Investig. Dermatol. 2019, 139, 1329–1338.e7. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, A.; Kaga, N.; Nakanishi, Y.; Ohno, H.; Miyamoto, J.; Kimura, I.; Hori, S.; Sasaki, T.; Hiramatsu, K.; Okumura, K.; et al. Maternal high fiber diet during pregnancy and lactation influences regulatory T cell differentiation in offspring in mice. J. Immunol. 2017, 199, 3516–3524. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhuo, M.; Huang, X.; Huang, Y.; Zhou, J.; Xiong, D.; Li, J.; Liu, Y.; Pan, Z.; Li, H.; et al. Altered gut microbiota associated with symptom severity in schizophrenia. PeerJ 2020, 8, e9574. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Song, J.; Ke, P.; Kong, L.; Lei, B.; Zhou, J.; Huang, Y.; Li, H.; Li, G.; Chen, J.; et al. The gut microbiome is associated with brain structure and function in schizophrenia. Sci. Rep. 2021, 11, 9743. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| List of Antibodies Used in Immunohistochemistry | |||
|---|---|---|---|
| Antibody | Clone | Company | Order number |
| Anti-CD3 | SP7 | Abcam | 67849 |
| Anti-Ly6G | 1A8 | Aviva Systems | OATA00269 |
| Anti-Foxp3 | FJK-16s | Thermo Fisher Scientific | 14-5773-82 |
| Anti-Ki67 | Polyclonal | Thermo Fisher Scientific | PA5-19462 |
| TaqMan® Gene Expression Assay IDs of genes used in quantitative real-time polymerase chain reaction | |||
| Gene | Assay ID | Gene | Assay ID |
| Ifng | Mm01168134_m1 | Il22 | Mm00444241_m1 |
| Il17a | Mm00439618_m1 | Krt16 | Mm01306670_g1 |
| Gapdh | Mm99999915_g1 | Cdkn1a | Mm04207341_m1 |
| Il10 | Mm01288386_m1 | Ccl20 | Mm01268754_m1 |
| Tgfb1 | Mm01178820_m1 | Cxcl1 | Mm04207460_m1 |
| Foxp3 | Mm00475162_m1 | Cxcl2 | Mm00436450_m1 |
| Tnfa | Mm00443258_m1 | Il1b | Mm00434228_m1 |
| Il17c | Mm00521397_m1 | Il17f | Mm00521423_m1 |
| Il6 | Mm00446190_m1 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshida, M.; Funasaka, Y.; Saeki, H.; Yamamoto, M.; Kanda, N. Dietary Fiber Inulin Improves Murine Imiquimod-Induced Psoriasis-like Dermatitis. Int. J. Mol. Sci. 2023, 24, 14197. https://doi.org/10.3390/ijms241814197
Yoshida M, Funasaka Y, Saeki H, Yamamoto M, Kanda N. Dietary Fiber Inulin Improves Murine Imiquimod-Induced Psoriasis-like Dermatitis. International Journal of Molecular Sciences. 2023; 24(18):14197. https://doi.org/10.3390/ijms241814197
Chicago/Turabian StyleYoshida, Mai, Yoko Funasaka, Hidehisa Saeki, Masami Yamamoto, and Naoko Kanda. 2023. "Dietary Fiber Inulin Improves Murine Imiquimod-Induced Psoriasis-like Dermatitis" International Journal of Molecular Sciences 24, no. 18: 14197. https://doi.org/10.3390/ijms241814197
APA StyleYoshida, M., Funasaka, Y., Saeki, H., Yamamoto, M., & Kanda, N. (2023). Dietary Fiber Inulin Improves Murine Imiquimod-Induced Psoriasis-like Dermatitis. International Journal of Molecular Sciences, 24(18), 14197. https://doi.org/10.3390/ijms241814197