
Stachydrine Hydrochloride Regulates the NOX2-ROS-Signaling Axis in Pressure-Overload-Induced Heart Failure


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. STA Counteracts Stress-Loaded HF in Mice
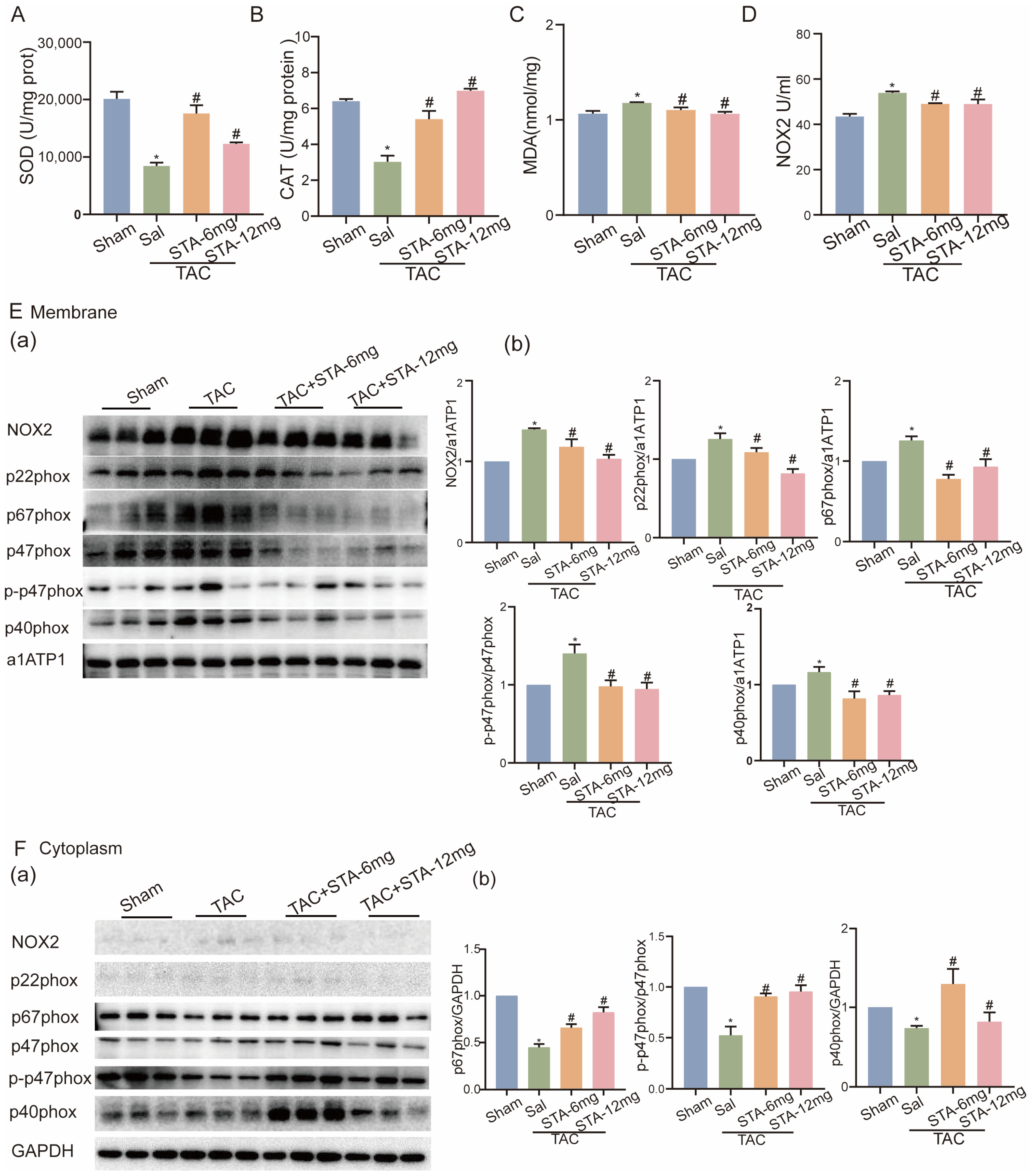
2.2. STA Relieves the ROS and ROS-Dependent Myocardial Injury in Stress-Loaded HF in Mice
2.3. STA Suppresses NOX2 Protein Expression and Activation in Stress-Loaded HF in Mice
2.4. STA Reduces the Production of ROS and Expression of NOX2 in Cardiomyocytes Cultured with H2O2 or Phenylephrine (PE)
2.5. STA Sustains NOX2-Related Ca2+ Homeostasis and Improves Contractile Function in Cardiomyocytes under Oxidative Stress
2.6. STA Inhibits NOX2-Related Ca2+-Dependent Regulatory Protein Expression in Stress-Loaded HF
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Animals
4.3. Transverse Aortic Constriction (TAC) Surgery
4.4. Echocardiography Analysis
4.5. Adult Mouse Cardiac Myocyte (AMCM) Isolation, Culture, and Treatment
4.6. Immunofluorescence Staining
4.7. H9c2 Culture and Treatment
4.8. Neonatal Rat Cardiomyocyte (NRCM) Culture and Treatment Protocol
4.9. Western Blot
4.10. ELISA
4.11. Detection of Intracellular ROS
4.12. DCFH-DA Staining
4.13. Detection of Superoxide Dismutase (SOD), Catalase (CAT), and Malondialdehyde (MDA)
4.14. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Benjamin, E.J.; Virani, S.S.; Callaway, C.W.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Chiuve, S.E.; Cushman, M.; Delling, F.N.; Deo, R.; et al. Heart Disease and Stroke Statistics-2018 Update: A Report From the American Heart Association. Circulation 2018, 137, e67–e492. [Google Scholar] [CrossRef] [PubMed]
- Van Nuys, K.E.; Xie, Z.; Tysinger, B.; Hlatky, M.A.; Goldman, D.P. Innovation in Heart Failure Treatment: Life Expectancy, Disability, and Health Disparities. JACC Heart Fail. 2018, 6, 401–409. [Google Scholar] [CrossRef]
- Writing Committee Members; ACC/AHA Joint Committee Members. 2022 AHA/ACC/HFSA Guideline for the Management of Heart Failure. J. Card. Fail. 2022, 28, e1–e167. [Google Scholar] [CrossRef]
- Chen, H.H.; Zhao, P.; Zhao, W.X.; Tian, J.; Guo, W.; Xu, M.; Zhang, C.; Lu, R. Stachydrine ameliorates pressure overload-induced diastolic heart failure by suppressing myocardial fibrosis. Am. J. Transl. Res. 2017, 9, 4250–4260. [Google Scholar]
- Hu, P.; Guo, S.; Yang, S.; Wang, S.; Wang, S.; Shan, X.; Zhao, P.; Guo, W.; Xu, M.; Zhang, C.; et al. Stachytine Hydrochloride Improves Cardiac Function in Mice with ISO-Induced Heart Failure by Inhibiting the alpha-1,6-Fucosylation on N-Glycosylation of beta1AR. Front. Pharmacol. 2021, 12, 834192. [Google Scholar] [CrossRef]
- Chen, H.H.; Wang, S.N.; Cao, T.T.; Zheng, J.L.; Tian, J.; Shan, X.L.; Zhao, P.; Guo, W.; Xu, M.; Zhang, C.; et al. Stachydrine hydrochloride alleviates pressure overload-induced heart failure and calcium mishandling on mice. J. Ethnopharmacol. 2020, 248, 112306. [Google Scholar] [CrossRef]
- van der Pol, A.; van Gilst, W.H.; Voors, A.A.; van der Meer, P. Treating oxidative stress in heart failure: Past, present and future. Eur. J. Heart Fail. 2019, 21, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Mallat, Z.; Philip, I.; Lebret, M.; Chatel, D.; Maclouf, J.; Tedgui, A. Elevated levels of 8-iso-prostaglandin F2alpha in pericardial fluid of patients with heart failure: A potential role for in vivo oxidant stress in ventricular dilatation and progression to heart failure. Circulation 1998, 97, 1536–1539. [Google Scholar] [CrossRef]
- Takimoto, E.; Champion, H.C.; Li, M.; Ren, S.; Rodriguez, E.R.; Tavazzi, B.; Lazzarino, G.; Paolocci, N.; Gabrielson, K.L.; Wang, Y.; et al. Oxidant stress from nitric oxide synthase-3 uncoupling stimulates cardiac pathologic remodeling from chronic pressure load. J. Clin. Investig. 2005, 115, 1221–1231. [Google Scholar] [CrossRef] [PubMed]
- Huynh, K.; Bernardo, B.C.; McMullen, J.R.; Ritchie, R.H. Diabetic cardiomyopathy: Mechanisms and new treatment strategies targeting antioxidant signaling pathways. Pharmacol. Ther. 2014, 142, 375–415. [Google Scholar] [CrossRef]
- Parajuli, N.; Patel, V.B.; Wang, W.; Basu, R.; Oudit, G.Y. Loss of NOX2 (gp91phox) prevents oxidative stress and progression to advanced heart failure. Clin. Sci. 2014, 127, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Sorescu, D.; Weiss, D.; Lassegue, B.; Clempus, R.E.; Szocs, K.; Sorescu, G.P.; Valppu, L.; Quinn, M.T.; Lambeth, J.D.; Vega, J.D.; et al. Superoxide production and expression of nox family proteins in human atherosclerosis. Circulation 2002, 105, 1429–1435. [Google Scholar] [CrossRef] [PubMed]
- Cao, T.T.; Chen, H.H.; Dong, Z.; Xu, Y.W.; Zhao, P.; Guo, W.; Wei, H.C.; Zhang, C.; Lu, R. Stachydrine Protects Against Pressure Overload-Induced Cardiac Hypertrophy by Suppressing Autophagy. Cell Physiol. Biochem. 2017, 42, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Murugesan, P.; Huang, K.; Cai, H. NADPH oxidases and oxidase crosstalk in cardiovascular diseases: Novel therapeutic targets. Nat. Rev. Cardiol. 2020, 17, 170–194. [Google Scholar] [CrossRef]
- Zima, A.V.; Blatter, L.A. Redox regulation of cardiac calcium channels and transporters. Cardiovasc. Res. 2006, 71, 310–321. [Google Scholar] [CrossRef]
- McNutt, N.S.; Fawcett, D.W. The ultrastructure of the cat myocardium. II. Atrial muscle. J. Cell Biol. 1969, 42, 46–67. [Google Scholar] [CrossRef]
- Bers, D.M. Cardiac sarcoplasmic reticulum calcium leak: Basis and roles in cardiac dysfunction. Annu. Rev. Physiol. 2014, 76, 107–127. [Google Scholar] [CrossRef]
- Kranias, E.G.; Hajjar, R.J. Modulation of cardiac contractility by the phospholamban/SERCA2a regulatome. Circ. Res. 2012, 110, 1646–1660. [Google Scholar] [CrossRef]
- Liu, X.; Shan, X.; Chen, H.; Li, Z.; Zhao, P.; Zhang, C.; Guo, W.; Xu, M.; Lu, R. Stachydrine Ameliorates Cardiac Fibrosis Through Inhibition of Angiotensin II/Transformation Growth Factor beta1 Fibrogenic Axis. Front. Pharmacol. 2019, 10, 538. [Google Scholar] [CrossRef]
- Wolfram, R.; Oguogho, A.; Palumbo, B.; Sinzinger, H. Enhanced oxidative stress in coronary heart disease and chronic heart failure as indicated by an increased 8-epi-PGF(2alpha). Eur. J. Heart Fail. 2005, 7, 167–172. [Google Scholar] [CrossRef]
- Nediani, C.; Raimondi, L.; Borchi, E.; Cerbai, E. Nitric oxide/reactive oxygen species generation and nitroso/redox imbalance in heart failure: From molecular mechanisms to therapeutic implications. Antioxid. Redox Signal 2011, 14, 289–331. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, L.D.; Greenstein, J.L.; O’Rourke, B.; Winslow, R.L. An integrated mitochondrial ROS production and scavenging model: Implications for heart failure. Biophys. J. 2013, 105, 2832–2842. [Google Scholar] [CrossRef] [PubMed]
- Ravi, V.; Jain, A.; Taneja, A.; Chatterjee, K.; Sundaresan, N.R. Isolation and Culture of Neonatal Murine Primary Cardiomyocytes. Curr. Protoc. 2021, 1, e196. [Google Scholar] [CrossRef]
- Wang, M.; Shu, Z.J.; Wang, Y.; Peng, W. Stachydrine hydrochloride inhibits proliferation and induces apoptosis of breast cancer cells via inhibition of Akt and ERK pathways. Am. J. Transl. Res. 2017, 9, 1834–1844. [Google Scholar] [PubMed]
- Tian, W.; Xu, H.; Fang, F.; Chen, Q.; Xu, Y.; Shen, A. Brahma-related gene 1 bridges epigenetic regulation of proinflammatory cytokine production to steatohepatitis in mice. Hepatology 2013, 58, 576–588. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, S.; Liang, Y.; Yang, S.; Fu, M.; Shan, X.; Zhang, C.; Chen, H.; Zhao, P.; Lu, R. Stachydrine Hydrochloride Regulates the NOX2-ROS-Signaling Axis in Pressure-Overload-Induced Heart Failure. Int. J. Mol. Sci. 2023, 24, 14369. https://doi.org/10.3390/ijms241814369
Lu S, Liang Y, Yang S, Fu M, Shan X, Zhang C, Chen H, Zhao P, Lu R. Stachydrine Hydrochloride Regulates the NOX2-ROS-Signaling Axis in Pressure-Overload-Induced Heart Failure. International Journal of Molecular Sciences. 2023; 24(18):14369. https://doi.org/10.3390/ijms241814369
Chicago/Turabian StyleLu, Shuang, Yueyang Liang, Songru Yang, Mengwei Fu, Xiaoli Shan, Chen Zhang, Huihua Chen, Pei Zhao, and Rong Lu. 2023. "Stachydrine Hydrochloride Regulates the NOX2-ROS-Signaling Axis in Pressure-Overload-Induced Heart Failure" International Journal of Molecular Sciences 24, no. 18: 14369. https://doi.org/10.3390/ijms241814369