Abstract
Pine wilt disease (PWD) is a devastating disease that threatens pine forests worldwide, and breeding resistant pines is an important management strategy used to reduce its impact. A batch of resistant seeds of P. thunbergii was introduced from Japan. Based on the resistant materials, we obtained somatic plants through somatic embryogenesis. In this study, we performed transcriptome analysis to further understand the defense response of resistant somatic plants of P. thunbergii to PWD. The results showed that, after pine wood nematode (PWN) infection, resistant P. thunbergii stimulated more differential expression genes (DEGs) and involved more regulatory pathways than did susceptible P. thunbergii. For the first time, the alpha-linolenic acid metabolism and linoleic acid metabolism were intensively observed in pines resisting PWN infection. The related genes disease resistance protein RPS2 (SUMM2) and pathogenesis-related genes (PR1), as well as reactive oxygen species (ROS)-related genes were significantly up-expressed in order to contribute to protection against PWN inoculation in P. thunbergii. In addition, the diterpenoid biosynthesis pathway was significantly enriched only in resistant P. thunbergii. These findings provided valuable genetic information for future breeding of resistant conifers, and could contribute to the development of new diagnostic tools for early screening of resistant pine seedlings based on specific PWN-tolerance-related markers.
1. Introduction
Pinus thunbergii Parl. was the most prestigious of the coniferous bonsai trees, which was a type of positive evergreen tree native to Japan and the southern coast of Korea [1,2] It was widely used in coastal urban greening and coastal windbreaks due to its resistance to barrenness and strong adaptability, especially to sea fog and sea winds [3,4,5,6]. It has been cultivated in Qingdao City, China, since 1900 and has been widely planted in coastal cities in China for almost 120 years [7,8]. However, the population of P. thunbergii has declined dramatically since the outbreak of pine wilt disease (PWD) [9]. PWD has severely limited the development of P. thunbergii forests.
PWD was a devastating forest disease of the Pinus species, caused by the pine wood nematode (PWN) Bursaphelenchus xylophilus [10]. It was first reported in 1905 in Nagasaki, Japan, as a serious forest disease affecting the Pinus species (P. thunbergii and P. densiflora) [11,12]. Now, PWD has been reported in the USA, Canada, Mexico, China, Japan, Korea, Portugal and Spain [13,14]. Since PWD became endemic in various countries, its spread has been beyond expectations and caused huge economic losses, especially in China and Japan [15,16,17]. Among the 353 species of Pinaceae, up to 106 Pinus species could be infected by PWN, and the host range of PWN is expanding due to its adaptability [18]. In addition to the wide host range, the hazard range of PWD has been gradually expanding due to global warming [19,20]. Nowadays, PWD has become the greatest threat to the development of pine forests worldwide [21,22]. The comprehensive understanding and control of PWD is a work-in-progress. Many methods have been investigated to control PWD, and although none has been proven to completely control the disease, they have been effective in slowing down the spread of PWD [23]. Within this holistic framework, the use of tolerant/resistant P. thunbergii cultivars was probably the most efficient and environmentally friendly approach to controlling PWD. Due to the extreme difficulty of preventing the disease, there has been a growing enthusiasm for processes of breeding pine trees for resistance to PWD. Several studies have focused on finding sources of genetic resistance to PWN [24] Japan was the first country to select resistant Pinus species from PWD-endemic areas [25,26] Our group introduced thirteen resistant families (Nos. 28–40) of P. thunbergii seeds in 2004, and the seedlings from the introduced resistant families showed PWD resistance after artificial inoculation [27]. Based on the resistant materials, many somatic plants were obtained by our group [28].
Plant resistance mechanisms are important for maintaining the survival and health of organisms against adverse environments and pathogen invasions [15,29,30]. Currently, there have been more than 2000 reports of pine resistance to PWN, but the mechanism of pine resistance to PWN is still inconclusive [16]. Identifying the mechanisms of pine resistance to PWN could help us to better understand the pathogenesis and develop precise prevention or control measures [31]. This would facilitate better control of PWD epidemics in the future. Pines have enormous genomes with large amounts of repetitive DNA, making whole-genome sequencing projects difficult [32,33]. Transcriptome analysis based on next-generation sequencing data (RNA-Seq) provides information on all transcriptional activity in a cell or organism [34]. For pines, RNA-Seq was an efficient method to generate functional genomic data due to the absence of a valid reference genome sequence [35]. To comprehensively understand the pathogenic mechanism and reduce the damage caused by PWD, the genetic analysis studies had been conducted after inoculating the pines with PWN, including such plants as P. densiflora [36], P. pinaster [37], P. massoniana [38] and P. radiata [39]. However, to our knowledge, although transcriptomic studies of PWD resistance in P. thunbergii had been reported, the information on PWN resistance genes in somatic plants was rare.
In this paper, we investigated for the first time the genetic information of somatic plants in response to PWN infection. The 3-year-old somatic plants of P. thunbergii were artificially inoculated with a highly virulent strain of PWN to test their resistance to PWD, and differentially expressed genes and metabolic pathways associated with PWN resistance were revealed by comparing the responses of susceptible and resistant P. thunbergii at the transcriptome level; candidate resistant genes that explained the different levels of PWN susceptibility were identified. Our characterization of the response to PWN infection would contribute to the future molecular breeding of PWN-resistant P. thunbergii and further understanding of the mechanisms of resistance to PWN inoculation in Pinus species.
2. Results
2.1. DEGs Vary in Response to PWN Inoculation of Susceptible and Resistant P. thunbergii
The resistant and susceptible P. thunbergii showed very different regulatory processes in response to PWN infection. In PWN-resistant P. thunbergii, the more DEGs (including up- and down-regulated genes) were triggered in response to PWN infection, e.g., 611 up- and 253 down-regulated DEGs responded to PWN infection in PWN-resistant P. thunbergii from 1 to 3 dpi, whereas only 21 up- and 46 down-regulated DEGs were induced in susceptible P. thunbergii. Furthermore, the DEGs’ evolutionary trends were different, with up-regulated DEGs predominating in PWN-resistant P. thunbergii during the primary stage of PWN infection (from 1 to 3 dpi), and up- and down-regulated genes fading in and out thereafter. The opposite was true for susceptible P. thunbergii (Figure 1A,B). During the different response processes to PWN infection, the common genes involved in both resistant and susceptible P. thunbergii were relatively few. For example, 10 common DEGs were found between 1d vs. 3d and 3d vs. 7d in susceptible P. thunbergii, and 73 common DEGs existed in PWN-resistant P. thunbergii (Figure 1C,D). The common DEGs for both susceptible (10/332) and PWN-resistant P. thunbergii (73/1783) were less than 5% of the total number. In addition, some specific DEGs were present in the PWN infection process. For example, 57 and 265 specific DEGs were found in 1 d vs. 3 d and 3 d vs. 7 d stage, respectively. The number of DEGs involved in the defense response of P. thunbergii varied considerably at different stages of PWN infection development. This suggested that the defense responses of resistant and susceptible P. thunbergii were quite different as the disease progressed.
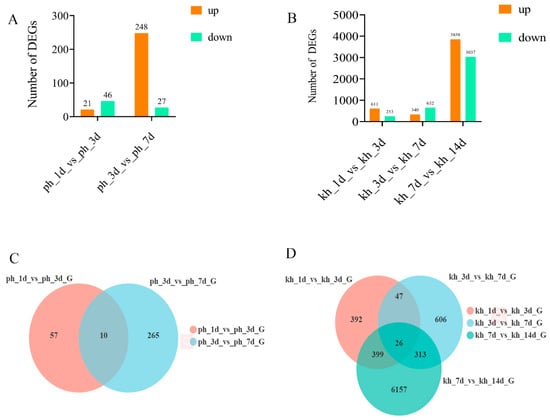
Figure 1.
Comparison of DEGs between susceptible and resistant P. thunbergii at different disease stages. (A,B) indicated the number of DEGs obtained in susceptible and resistant P. thunbergii at infection stages, respectively. (C,D) indicated the number and overlapping relationships of DEGs in venn diagram between susceptible and resistant P. thunbergii at different infection stages, respectively.
2.2. Functional Annotation and Enrichment Analysis of DEGs
To gain a functional perspective of the DEGs associated with resistance to PWN, we performed a GO annotation analysis (p < 0.05) of these DEGs in resistant and susceptible phenotypes during the disease’s progression. Four DEG sets of two PWN infection stages during disease progression were subjected to GO annotation analysis to identify over-represented GO terms. We found that the vast majority of DEGs were different in both phenotypes and that fewer GO terms were activated in susceptible P. thunbergii, including biological process, cellular component and molecular function category, than in the PWN-resistant P. thunbergii, especially in the first infection stage (1 d vs. 3 d). Many GO terms were activated only in resistant P. thunbergii, such as: developmental process, multi-organism process, organelle part and so on (Figure 2A). Furthermore, we performed a KEGG enrichment analysis (p < 0.05) of these DEGs. These results also showed that the enriched pathways associated with the DEGs were more diverse in resistant P. thunbergii. Only two pathways (mitogen-activated protein kinase (MAPK) signaling pathway—plant, biosynthesis of various secondary metabolites—part 2) associated with the DEGs were enriched in the early infection stages (1 d vs. 3 d) of susceptible P. thunbergii, compared to eight in PWN-resistant P. thunbergii. And the alpha-linolenic acid metabolism and linoleic acid metabolism pathways were enriched in susceptible P. thunbergii at later infection stages (3 d vs. 7 d), compared to the seven pathways in PWN-resistant P. thunbergii (Table 1). The alpha-linolenic acid metabolism and linoleic acid metabolism pathways were activated earlier in PWN-resistant P. thunbergii (1 d vs. 3 d) than in susceptible P. thunbergii (3 d vs. 7 d), while the “biosynthesis of various secondary metabolites—part 2” pathway was activated later in PWN-resistant P. thunbergii (3 d vs. 7 d) than in susceptible P. thunbergii (1 d vs. 3 d). In addition, the MAPK signaling pathway was active for longer in resistant P. thunbergii. In general, the findings that PWN-resistant P. thunbergii activated additional pathways to suppress PWN infection and temporal changes of gene expression involved in the defense response to PWN inoculation in susceptible and resistant P. thunbergii were clearly demonstrated. Further analysis was based on these different pathways and unigene data sets.
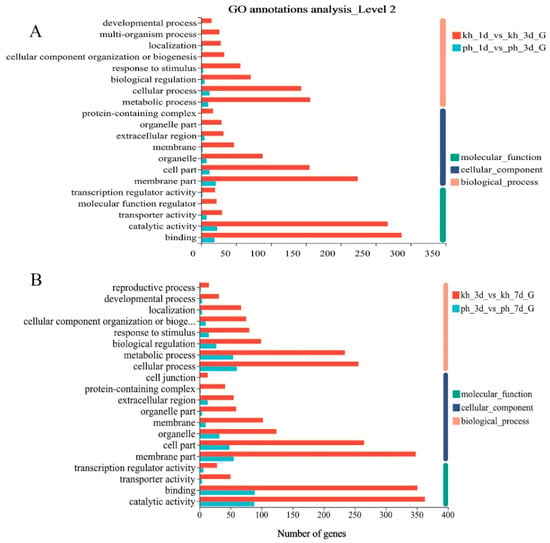
Figure 2.
Enriched biological processes in DEGs by GO annotation for susceptible and resistant P. thunbergii at different infection stages. (A) indicated GO annotation of susceptible and resistant P. thunbergii at first infection stage (1d vs 3d). (B) indicated GO annotation of susceptible and resistant P. thunbergii at second infection stage (3d vs 7d).

Table 1.
The KEGG analysis of DEGs in susceptible and resistant P. thunbergii.
2.3. The qPCR Validation
High-quality RNA (OD 260/280 = 1.8~2.2, RIN ≥ 7.9) was used to generate a sequencing library (Table S1 and Figure S1). To validate the results of the RNA-Seq data, seven DEGs (Table S2) were selected to assess their expression by qPCR. The unigenes selected for qRT-PCR analysis were mainly involved in ROS-responsive genes, resistance proteins, signal transduction and membrane components. The qPCR assay for the selected transcripts shows an expression pattern similar to that obtained by RNA-Seq data analysis. The differential expression detected by RT-PCR was consistent with the RNA-Seq expression profiles of each of the selected transcripts (Figure 3).
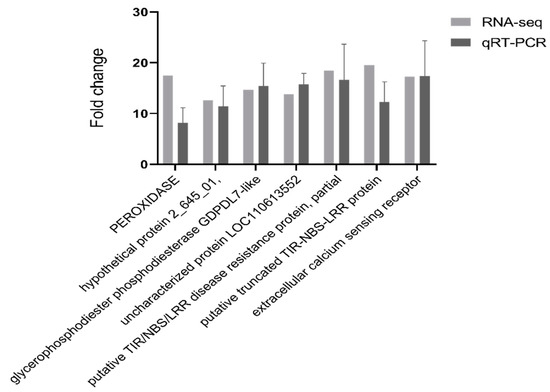
Figure 3.
qRT-PCR validation of selected transcripts for validation. Relative expression levels of qRT-PCR are calculated using elongation factor 1-alpha as the internal control. The data are expressed as the mean (±SE). Error bars represent the SE.
2.4. The Alpha-Linolenic Acid and Linoleic Acid Metabolism Were Involved in the Response to PWN
In this study, alpha-linolenic acid and linoleic acid metabolism were significantly different in susceptible (p < 0.05) and resistant P. thunbergii (p < 0.001), and they were activated earlier in resistant P. thunbergii (Figure 4A). In the linoleic acid metabolism, the expression of lindoleate 9S-lipoxygenase (LOX1-5) and 13 s-lipoxygenase (LOX2S) showed significant differences between susceptible and resistant P. thunbergii. Three unigenes encoding LOX1-5 were up-regulated in susceptible P. thunbergii. At the same time, more unigenes encoding LOX1-5 were activated in resistant P. thunbergii, and we found that six of the seven unigenes encoding LOX1-5 were up-regulated and one (TRINITY_DN3608_c0_g1) was down-regulated. The unigenes encoding LOX2S were up-regulated, and only identified in resistant P. thunbergii. In contrast to the susceptible phenotype, the LOX2S-catalyzed branch was activated in the resistant phenotype, in addition to the LOX1-5-catalyzed branch being strengthened.
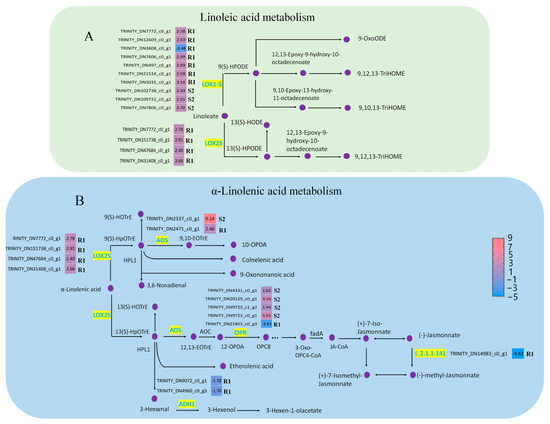
Figure 4.
DEGs involved in alpha-linolenic acid metabolism and linoleic acid metabolism in P. thunbergii. (A) indicated linoleic acid metabolism pathway. (B) indicated alpha-linolenic acid metabolism pathway. Enzymes involved in each step are shown in purple, and the green boxes represent DEGs encoding enzyme activity. R represents resistant P. thunbergii. S represents susceptible P. thunbergii. R1 represents the first stage of the resistant P. thunbergii inoculated with PWN (1 d vs. 3 d). S2 represents the second stage of susceptible P. thunbergii inoculated with PWN (3 d vs. 7 d). LOX1-5 (E5.5.1.13) represents lindoleate 9S-lipoxygenase. LOX2S (E1.14.11.13) represents lipoxygenase. AOS represents hydroperoxide dehydratase. OPR represents 12-oxophytodienoic acid reductase. ADH1 represents alcohol dehydrogenase class-P, E2.2.1.141 represents jasmonate O-methyltransferase.
In alpha-linolenic acid metabolism, five enzymes showed significantly different expression, namely, hydroperoxide dehydratase (AOS), 12-oxophytodienoic acid reductase (OPR), LOX1-5, alcohol dehydrogenase class P (ADH1) and jasmonate O-methyltransferase (E.2.1.1.141). Among these, OPR, ADH1 and jasmonate O-methyltransferase were down-regulated in resistant P. thunbergiiat stage R1, whereas AOS and LOX2S were up-regulated (Figure 4B). In susceptible P. thunbergii, only AOS and OPR were altered at the S2 stage. This suggested that PWN activated a completely different defense system in resistant P. thunbergii, involving more unigenes and metabolic branches in linoleic acid and alpha-linolenic acid metabolism.
2.5. Regulation of the “MAPK Signaling Pathway—Plant” and “Biosynthesis of Various Secondary Metabolites—Part 2” Contributed to PWN Resistance
The results showed that a total of 4 and 17 DEGs were identified in susceptible and resistant P. thunbergii, respectively (Table S3). In the MAPK signaling pathway, the disease resistance protein RPS2 (SUMM2), pathogenesis-related genes (PR1) and catalase (CAT1) were significantly up-regulated in susceptible P. thunbergii in the first stage of PWN infection (S1), whereas they were not significantly expressed in the second stage (S2) of PWN infection (Figure 5A). On the other hand, more DEGs were activated in resistant P. thunbergii, including basic endochitinase B (CHIB), nucleoside-diphosphate kinase (NDPK2), abscisic acid receptor PYL familiar (PYL) and serine/threonine-protein kinase OXI1 (OXI1), in addition to SUMM2, PR1 and CAT1. The expression of SUMM2 and CHIB was significantly up-regulated, while that of NDPK2 and PYL was significantly down-regulated. The PR1 gene was up-regulated at the S1 stage in susceptible P. thunbergii, but was significantly (p < 0.001) up-regulated at both the R1 and R2 stages in resistant P. thunbergii (Figure 5B).
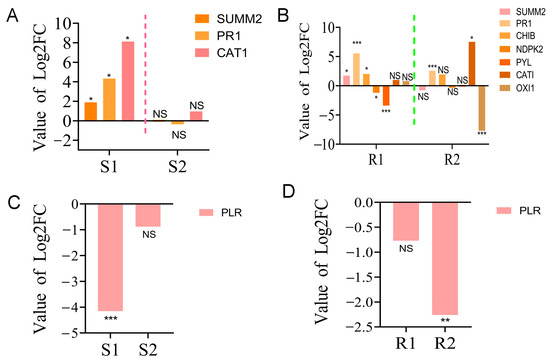
Figure 5.
The expression of partial DEGs in “MAPK signaling pathway—plant” and “biosynthesis of various secondary metabolites—part 2” pathways. (A,B) The expression of DEGs in the “MAPK signaling pathway—plant” in susceptible and PWN-resistant P. thunbergii, respectively. (C,D) Expression of DEGs in “biosynthesis of various secondary metabolites—part 2” in susceptible and PWN-resistant P. thunbergii, respectively. S1 represents the 1st stage (1 d vs. 3 d) of PWN infection in susceptible P. thunbergii. S2 represents the 2nd stage (3 d vs. 7 d) of PWN infection in susceptible P. thunbergii. R1 represents the 1st stage (1d vs. 3d) of PWN infection in resistant P. thunbergii. R2 represents the 2nd stage (3 d vs. 7 d) of PWN infection in resistant P. thunbergii. * and ** indicate significant differences at p < 0.05 and p < 0.01, respectively. *** indicate significant differences at p < 0.001.
In the biosynthesis pathway of various secondary metabolites, one unigene encoding PLR was down-regulated in susceptible P. thunbergii, while three unigenes encoding PLR were down-regulated in resistant P. thunbergii (Table S3), and the unigene expression trends were completely opposite (Figure 5C,D). These data strongly suggested that substantial changes in defense pathways were activated by PWN between the susceptible and resistant P. thunbergii, and that more genes and pathways were activated to defend against PWN infection in resistant P. thunbergii. However, the specific resistance function of DEGs needs to be further investigated.
2.6. Characterization of Gene Expression Related to Terpenoids Biosynthesis
In this study, DEGs encoding terpenoids were the most abundant, whereas DEGs encoding sesquiterpenes and triterpenes were not detected. In addition, the number of terpene-encoding DEGs was lower and decreased continuously with the PWN infection (Figure 6A). This suggested that terpenoids might play an important role in the response of P. thunbergii to PWN infection. Another result supported this conclusion. We found that three terpenoid synthesis pathways were enriched in resistant P. thunbergii by PWN infection, namely, monoterpenoid biosynthesis (p = 0.0136), diterpenoid biosynthesis (p < 0.001) and terpenoid backbone biosynthesis (p = 0.0628) (Figure S2). And only the diterpenoid biosynthesis pathway was enriched in both susceptible and resistant P. thunbergii. Nine DEGs involved in the diterpenoid biosynthesis were found in susceptible and resistant P. thunbergii. Among these DEGs, the expression levels of ent-copalyl diphosphate synthase (E5.5.1.13), (13E)-labda-7,13-dien-15-ol synthase (E3.1.7.10) and gibberellin 2beta-dioxygenase (E1.14.11.13) were significantly different (Figure 6B). The ent-copalyl diphosphate synthase was down-regulated in the resistant phenotype at the R1 stage, but significantly up-regulated in the susceptible P. thunbergii at the S2 stage. Another synthase with geranylgeranyl-pp (GGPP) as substrate varied in susceptible and resistant P. thunbergii. In addition, gibberellin 2beta-dioxygenase was only activated in the resistant phenotype. This suggested that terpenoids were involved in the resistance of P. thunbergii to PWN.
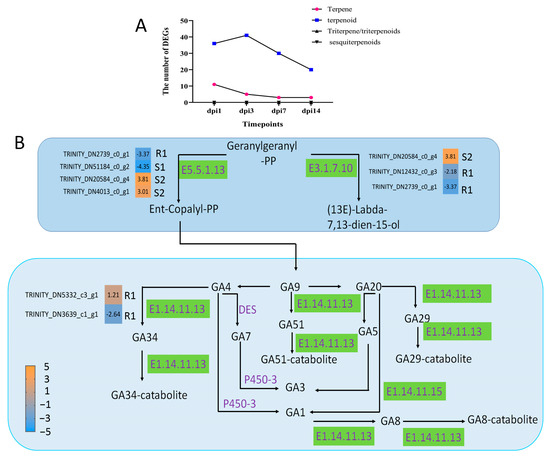
Figure 6.
DEGs involved in diterpenoid biosynthesis in the stem of P. thunbergii. (A) indicates the number of DEG terpenoid types. (B) indicated diterpenoid biosynthesis pathway. Enzymes involved in each step are shown in purple, and the green boxes represent DEGs encoding enzyme activity. R represents resistant P. thunbergii. S represents susceptible P. thunbergii. R1 represents the first stage of the resistant P. thunbergii inoculated with PWN (1 d vs. 3 d). S1 represents the first stage of the susceptible P. thunbergii inoculation with PWN (1 d vs. 3 d). S2 represents the second stage of susceptible P. thunbergii inoculated with PWN (3 d vs. 7 d). E5.5.1.13 represents ent-copalyl diphosphate synthase. E3.1.7.10 represents (13E)-labda-7,13-dien-15-ol synthase. E1.14.11.13 represents gibberellin 2beta-dioxygenase.
3. Discussion
Breeding of pines for resistance to PWN is an effective means for controlling PWD. Currently, it is rarely possible to obtain somatic pine plants from selected elite material. Fortunately, our group had previously obtained some somatic plants of P. thunbergii from resistant material [28,40]. In this study, we wanted to investigate the differences at the transcriptome level of the black pine stem segment between resistant and susceptible trees inoculated with PWN. We found that there was a remarkable difference (p < 0.05) between the two phenotypes’ stem transcriptomes; the resistant phenotype activated more DEGs in response to PWN invasion, and the number of DEGs was always more than for the susceptible phenotype at each PWN infection stage (Figure 1). This was consistent with previous study of P. thunbergii inoculation with PWN [16]. However, in the study of P. massoniana, the number of DEGs in the resistant phenotype was always lower than in the susceptible phenotype at each time point [38]. This suggested that the response mechanisms to PWN infection were different between P. massoniana and P. thunbergii, possibly due to their different susceptibility, and therefore in-depth studies on susceptible pines were needed.
According to the GO and KEGG classification of DEGs, the differences in the significant (p < 0.05) GO terms and metabolic pathways between the susceptible and resistant P. thunbergii indicated qualitative and quantitative differences in the defense genes induced in response to PWN infection (Table 1 and Figure 2). A proportion of the DEGs in susceptible trees were involved in stress/defense response categories, such as the response to stimulus, biological regulation and transcriptional regulator activity. Furthermore, a large percentage of DEGs were categorized as involving the cellular process, metabolic process, membrane part, catalytic activity and binding. These GO terms were all enriched in the resistant phenotype and many more DEGs were involved in it. This suggested that the PWN-resistant P. thunbergii has developed sophisticated defense mechanisms to combat PWN invasion, such as blocking pathogen entry and activating a range of defense responses.
The alpha-linolenic acid metabolism and linoleic acid metabolism were, for the first time, intensively observed as to their functions against PWN infection in pines. In our present study, alpha-linolenic acid metabolism and linoleic acid metabolism were activated earlier in resistant P. thunbergii (R1 stage) than in susceptible P. thunbergii (S2 stage) after inoculation with PWN. Alpha-linolenic acid can inhibit innate immune responses associated with callose deposition in wheat [41], and the activated alpha-linolenic acid metabolic pathway has resulted in increased plant (Citrus junos) growth [42]. The genes LOX, AOS and OPR were also important for plant defenses against biotic and abiotic stresses in alpha-linolenic acid metabolism [43,44]. In our study, the JA-related synthetic enzymes LOX2S and AOS were up-regulated in both resistant and susceptible P. thunbergii, but the expression of OPR was exactly opposite between resistant (down-expression) and susceptible (up-expression) P. thunbergii. In the P. pinaster study, the expression levels of AOS and LOX2S were consistent with our results, but OPR was also up-regulated in susceptible trees, contrary to our results [37] The differential expression of OPR affected the downstream regulation of JA. Also, the JA played an essential role in the response to PWN infection and might be important for resistance [45]. Alpha-linolenic acid metabolism was known to be a precursor of JA biosynthesis [46,47]. Interestingly, the jasmonate O-methyltransferase (E2.2.1.141) was down-regulated in resistant P. thunbergii in our study, which would lead to a decrease in methyl-jasmonate. Rodrigues et al. [48] reported higher levels of methyl-jasmonate in susceptible trees in response to PWN in P. pinaster at 72 dpi. Hormone quantification showed that JA levels were significantly higher in inoculated P. pinaster compared to controls observed at the same time [37] Moreover, methyl-jasmonate promoted chlorophyll degradation in plants such as Chenopodium album [49] and apple fruit [50]. Therefore, we hypothesized that down-regulated expression of the jasmonate O-methyltransferase was one of the key factors for delayed wilting in resistant P. thunbergii. These results implied that the conversion of JA to methyl jasmonate had an important effect on the resistance of pines to PWN. However, their molecular mechanisms in response to the PWN infection need to be further investigated. Taken together, these results provide evidence that PWN inoculation can promote JA signaling by stimulating alpha-linolenic acid metabolism and linoleic acid metabolism to protect pines from pathogen infection.
MAPK signaling was one of the earliest responses following pathogen attack and played a key role in plant signaling in response to various stresses [51]. In this study, the “MAPK signaling pathway—plant” was significantly activated in both resistant (p < 0.001) and susceptible (p < 0.05) P. thunbergii (Table 1). Some genes involved in plant hypersensitive response (HR) and disease resistance proteins were enriched. The resistance protein SUMM2 was significantly up-regulated in expression during the first stage (R1 and S1) in both susceptible and resistant P. thunbergii infection with PWN. In addition, the PR1 was only significantly (p < 0.05) up-regulated at the S1 stage in susceptible phenotypes, but in resistant phenotypes, the PR1 was significantly (p < 0.001) up-regulated at both the R1 and R2 stages. In plant–pathogen interactions, resistance proteins recognized pathogen effectors and then initiated effector-triggered immunity, which was often associated with (HR) cell death [52,53]. The resistance protein SUMM2 mediated immunity and HR cell death [51,54]. HR cell death at the site of infection was critical for initiating systemic signals that activate distal plant immunity and ultimately lead to systemic acquired resistance (SAR) [55,56]. In addition, HR cell death has often been described as an immune strategy for the prevention of pathogen colonization through the recognition of adapted biotrophic or hemi-biotrophic pathogens [57,58]. Therefore, we proposed that SUMM2 could be an important resistance gene functioning to prevent further spread of the pathogen in P. thunbergii during the early stages of PWN infection. Furthermore, PR1 was an important defense protein which was commonly used as a marker for SAR [59]. Plant PR proteins were initially identified as proteins that were strongly induced under biotic and abiotic stresses [60,61]. The mode of action of most PR proteins has been well characterized, with the exception of PR1, which belongs to a large superfamily of proteins which shares a common CAP structural domain. The importance of these proteins in immune defense was illustrated by the fact that PR1 overexpression in plants resulted in increased resistance to pathogens [62]. The defense response of the PR gene (PR1-10) to PWN has been reported in previous studies in P. thunbergii, and PR1 was predominantly up-regulated for expression at the early infestation stage (1 and 3 dpi) (Hirao et al., 2012). This is consistent with our findings. Our results indicated that SUMM2 and PR-1 were induced by PWN in both susceptible and resistant trees for plant immunity associated with HR cell death, and the PR1 worked longer in the resistant phenotype defending against the PWN infection. Furthermore, we found that comparatively more DEGs involved in ROS mediation were activated by the resistant phenotype in response to PWN infection, such as CAT1, NDPK2, OXI1, whereas only CAT1 was identified as a gene regulating ROS in the susceptible phenotype. Harao, et al. [16] reported that the peroxidase (PR-10) were the characteristic significantly up-regulated genes in resistant P. thunbergii inoculation with PWN. In the P. massoniana study, the catalase, peroxidase, superoxidase dismutase and glutathione reductase genes were involved in the defense response to PWN. In comparison to previous reports in P. thunbergii, we have identified more antioxidant-related genes of resistant P. thunbergii functioning against PWN. And compared to the more non-susceptible P. massoniana, both resistant trees had more complex ROS regulation relative to susceptible trees, although the antioxidant genes were not exactly the same. These results implied that ROS scavenging capacity was closely related to pine defense against PWN infestation. ROS-related genes regulating the resistance of pines to PWN infestation will be a topic worthy of further study.
In the “biosynthesis of various secondary metabolites—part 2” pathway, the pinoresinol/lariciresinol reductase (PLR) marked a significant difference between resistant and susceptible P. thunbergii. The resistant phenotype activated more DEGs involved in PLR regulation and had a different gene expression pattern than the susceptible phenotype (Figure 5). The PLR catalyzed the sequential reduction of pinoresinol to secoisolariciresinol via lariciresinol, which could lead to the structural and stereochemical diversity of lignans [63]. Lignin also affected the mechanical strength of the cell wall, which has been considered the first physical line of defense against PWN infection [9,64]. A more differential expression of the PLR might be associated with the regulation of lignin in response to PWN infection in resistant P. thunbergii, suggesting that the resistant phenotype had a more complicated regulation of lignin as a physical defense to suppress PWN infection.
Terpenoid metabolism has played an important role in the resistance of pines to PWN [38]. In our study, the DEGs encoding diterpenoids were the most numerous, and the diterpenoid biosynthetic pathway was significantly enriched in resistant P. thunbergii. After the trunk of a conifer suffered an insect attack, pathogen invasion or mechanical wounding, oleoresin could be synthesized [65,66]. Monoterpenes, sesquiterpenes and diterpene resin acids are important components of oleoresin [67,68]. Monoterpenes and sesquiterpenes in turpentine oil could directly affect herbivores through the release of toxic volatiles, and diterpene resin acids form physical barriers at the wound site [68]. In P. thunbergii, diterpenoids have the most DEGs. Therefore, we suggest that the diterpenoids metabolism is the main mode of response to PWN infestation. Furthermore, the ent-copalyl diphosphate synthase and gibberellin 2beta-dioxygenase unigenes were significantly differentially expressed in the diterpenoid biosynthetic pathway in this study. Gibberellin 2beta-dioxygenase, a key gene in the gibberellin synthesis, was only activated in resistant P. thunbergii. Tanaka, et al. [69] reported that gibberellin 2beta-dioxygenase increased rice biomass under low-nutrient conditions. Prisic, et al. [70] reported that ent-copalyl diphosphate synthases (OsCPS1ent and OsCPS2ent) from rice (Oryza sativa) were involved in GA biosynthesis and related a secondary metabolism producing defensive phytochemicals. In our study, the resistant P. thunbergii positively regulated GA signaling after inoculation with PWN, suggesting that GA biosynthesis genes were involved in the resistance of P. thunbergii to PWN.
4. Materials and Methods
4.1. Plant Material
In this experiment, the three-year-old susceptible P. thunbergii and somatic plants of PWN-resistant P. thunbergii were used as experimental materials. The somatic plants were obtained from the 1539-1 cell line by somatic embryogenesis (Sun et al., 2019). The 1539-1 cell line was initiated from the resistant family 39 of P. thunbergii. We collected immature cones (open pollination, family 39) from the seed orchard, and used their female gametophyte to induce embryogenic tissue (1539-1) (Sun et al., 2019). The somatic embryos were obtained and germinated into somatic plants, which were then transplanted in field (Sun et al., 2023); these somatic plants were the experimental material for this paper. The susceptible P. thunbergii (mortality rates were all 100% after inoculation with PWN at 28 days in the previous tests) from Suqian City, China, was selected as the control.
4.2. Pine Wood Nematode Inoculation and Sampling
The three-year-old susceptible and resistant P. thunbergii trees were transplanted into pots and grown under the same conditions for three months, and then the plants were transferred to a forcing house (30 °C constant temperature) for inoculation with PWN. The PWN used in our study was the highly virulent isolate AMA3. Three branches per tree were inoculated (1000 nematodes per branch) in susceptible and resistant P. thunbergii trees. Samples were collected at 1, 3, 7 and 14 days post-inoculation (dpi) to assess the somatic plants’ resistance to PWD. We sampled inoculated trees (both susceptible and resistant trees) at 1, 3 and 7 dpi; resistant trees were also sampled at 14 dpi. The last sampling time was chosen based on previous results, in which the needles of resistant and susceptible P. thunbergii turned yellow at 14 and 7 dpi, respectively, which was also observed in this experiment. The 2 cm long segments of the stems below the inoculation sites were cut off and immediately placed in liquid nitrogen. These samples were then stored at −80 °C for further RNA extraction. Three trees representing resistant and susceptible P. thunbergii were selected as biological replicates for each treatment.
4.3. RNA Isolation Procedure and Quantification
Following inoculation of resistant and susceptible P. thunbergii with PWN, samples were collected at the critical time point of disease development and stored at −80 °C as a reserve. Total RNA was extracted from each PWN-inoculated tree sample at four time points using TRIzol® Reagent (plant RNA purification reagent for plant tissue) according to the instructions, while genomic DNA was removed using DNase I (TaKara, Nanjing, China). The RNA degradation and contamination was monitored using 1% agarose gels, while the integrity and purity of the total RNA quality was determined using a 2100 Bioanalyser (Agilent Technologies, Shanghai, China) and quantified using an ND-2000 (NanoDrop Technologies, Wilmington, DE, USA). Only high-quality RNA samples (OD 260/280 = 1.8~2.2, RIN ≥ 7.9) were used to develop a sequencing library.
4.4. Quantitative Real-Time PCR Analysis
The RNA samples used for the qRT-PCR and transcriptome sequencing were identical. To evaluate the accuracy and reproducibility of the RNA-Seq expression profiles, quantitative real-time polymerase chain reaction (qRT-PCR) analysis was performed to analyze the expression levels of regulated genes at four different time points. Quantitative RT-PCR was run on a 7900 Real Time PCR System (Applied Biosystems, California USA) using the SYBR Green detection method to verify the transcriptome sequencing results. Quantitative real-time PCR (qRT -PCR) was performed in a 20 μL reaction volume containing 10 μL of SYBR Green Master Mix (Vazyme Biotech, Nanjing, China).
4.5. De Novo Assembly and Annotation
The raw paired end reads were trimmed and quality control was performed using fastp with default parameters. The clean data from the samples were then used to perform de novo assembly using Trinity. The assembled transcripts were then evaluated and optimized using BUSCO protein groups and the GO and KEGG databases. BLASTX was used to identify the proteins with highest sequence similarity to the given transcripts to retrieve their functional annotations, while typical cut-off E-values of less than 1.0 × 10−5 were set.
4.6. Differential Expression Analysis and Functional Enrichment
To identify the DEGs (differential expression genes) between the resistant and susceptible P. thunbergii samples, the expression levels of each gene were calculated using the transcripts-per-million-reads (TPM) method. RSEM (http://deweylab.biostat.wisc.edu/rsem/, accessed on 20 September 2022) was used to quantify gene abundance. Differential expression analysis was performed using the DESeq2/DEGseq/edgeR/Limma/, where DEGs with |log2 (foldchange)| ≥ 1 and P-adjust ≤ 0.05 were considered to be significant. Furthermore, functional-enrichment analysis, including GO (Gene Ontology, http://www.geneontology.org, accessed on 8 January 2023) and KEGG (Kyoto Encyclopedia of Genes and Genomes, http://www.genome.jp/kegg/, accessed on 8 January 2023) was also performed to identify which of the DEGs were significantly enriched in GO terms. Metabolic pathways were then compared to the whole-transcriptome background at p ≤ 0.05.
4.7. Statistical Analysis
Data were analyzed using analysis of variance (ANOVA) SPSS19 software (SPSS Inc., Chicago, IL, USA). Results were expressed as percentages using nonparametric methods. In addition, the graphs were created using Prism (Patterns & Practices., Redmond, WA, USA) and Adobe Photoshop CS6 (64 bit) software (Adobe, San Jose, CA, USA).
5. Conclusions
This work was the first to use resistant P. thunbergii somatic plants to study the defense responses of P. thunbergii to PWN; it promotes the use of somatic plants in the future. In this study, the resistant P. thunbergii activated more genes to defend against the PWN inoculation. Resistance protein SUMM2, PR1 and ROS-related genes were significantly up-expressed to contribute to the protection of P. thunbergii against PWN inoculation. In addition, alpha-linolenic acid metabolism and linoleic acid metabolism contributed to PWN resistance by regulating the JA-related synthetic enzyme genes. In conclusion, this study provided the first comprehensive report of the somatic plant transcriptomes of P. thunbergii materials which are resistant in response to PWN, and it enhanced the genetic resource database for P. thunbergii.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms241814376/s1.
Author Contributions
J.Y. and X.W. supervised the research. T.S. carried out the experiments and analyzed the data. T.S. and M.U.R. drafted the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Key Research and Development Program of China (2021YFD1400900), and the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data described in this study can be found in the article and the Supplementary Material.
Acknowledgments
We thank Weiliang Kong, Lanxiang Lu and Tongyue Wen for useful discussions; and Lin Rui and Tongyue Wen for assistance in method optimization.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Maruyama, T.E.; Hosoi, Y. Post-maturation treatment improves and synchronizes somatic embryo germination of three species of Japanese pines. Plant Cell Tissue Organ Cult. 2012, 110, 45–52. [Google Scholar] [CrossRef]
- Ho, U.H.; Song, S.R.; Pak, H.S.; Kim, K.; Ho, T.S.; Ju, I.Y. Genetic evidence of stable northward extension of Pinus thunbergii Parl. forests in the Democratic People’s Republic of Korea. Genet. Resour. Crop Evol. 2022, 69, 2105–2114. [Google Scholar] [CrossRef]
- Choi, J.; Cha, D.; Kim, D.S.; Lee, S. Review of Japanese Pine Bast Scale, Matsucoccus matsumurae (Kuwana) (Coccomorpha: Matsucoccidae), Occurring on Japanese Black Pine (Pinus thunbergii Parl.) and Japanese Red Pine (P. densiflora Siebold & Zucc.) from Korea. Forests 2019, 10, 639. [Google Scholar]
- Zhang, P.; Wen, Y.; Wang, L.; Zhang, H.; Wang, G.G.; Wu, T. Leaf Structural Carbohydrate Decreased for Pinus thunbergii along Coast–Inland Gradients. Forests 2020, 11, 449. [Google Scholar] [CrossRef]
- Todo, C.; Yamase, K.; Ikeno, H.; Tanikawa, T.; Ohashi, M.; Hirano, Y. Maximum rooting depth of Pinus thunbergii Parl. estimated with depth at the center point of rotation in a tree-pulling experiment in a coastal forest in Japan. Forests 2022, 13, 1506. [Google Scholar] [CrossRef]
- Ding, X.L.; Lin, S.X.; Zhao, R.; Ye, J. First report of needle blight on Pinus thunbergii Parl. caused by Fusarium proliferatum in China. Plant Dis. 2022, 106, 2989. [Google Scholar] [CrossRef]
- Qi, Y.; Duan, C.; Ren, L.; Wu, H. Growth dynamics of galls and chemical defence response of Pinus thunbergii Parl. to the pine needle gall midge, Thecodiplosis japonensis Uchida & Inouye (Diptera: Cecidomyiidae). Sci. Rep. 2020, 10, 12289. [Google Scholar]
- Zhang, X.; Xing, J.; Zhu, X.; Zhang, B.; Liu, C.; Hong, L.; Liu, Y.; Chen, Y.; Wen, Z. Diversity and community structure of ectomycorrhizal fungi in Pinus thunbergii coastal forests bordering the Yellow Sea of China. Braz. J. Microbiol. 2021, 52, 801–809. [Google Scholar] [CrossRef]
- Wang, X.; Wu, X.; Wen, T.; Feng, Y.Q.; Zhang, Y. Transcriptomic analysis reveals differentially expressed genes associated with pine wood nematode resistance in resistant P. thunbergii. Tree Physiol. 2023, 43, 995–1008. [Google Scholar] [CrossRef]
- Zhang, Y.; Wen, T.Y.; Wu, X.Q.; Hu, L.; Qiu, Y.; Rui, L. The Bursaphelenchus xylophilus effector BxML1 targets the cyclophilin protein (CyP) to promote parasitism and virulence in pine. BMC Plant Biol. 2022, 22, 216. [Google Scholar] [CrossRef]
- Yano, M. Investigation on the cause of pine mortality in Nagasaki. Prefect. Sanrinkoho 1913, 4, 1–14. [Google Scholar]
- Mamiya, Y. History of pine wilt disease in Japan. J. Nematol. 1988, 20, 219–226. [Google Scholar] [PubMed]
- Togashi, K.; Shigesada, N. Spread of the pinewood nematode vectored by the Japanese pine sawyer: Modeling and analytical approaches. Popul. Ecol. 2006, 48, 271–283. [Google Scholar] [CrossRef]
- Abelleira, A.; Picoaga, A.; Mansilla, J.P.; Aguin, O. Detection of Bursaphelenchus Xylophilus, causal agent of pine wilt diseaseon Pinus pinaster in northwestern Spain. Plant Dis. 2011, 96, 770–780. [Google Scholar]
- Shin HLee, H.; Woo, K.S.; Noh, E.W.; Koo, Y.B.; Lee, K.J. Identification of genes upregulated by pinewood nematode inoculation in Japanese red pine. Tree Physiol. 2009, 29, 411–421. [Google Scholar] [CrossRef]
- Hirao, T.; Fukatsu, E.; Watanabe, A. Characterization of resistance to pine wood nematode infection in Pinus thunbergii using suppression subtractive hybridization. BMC Plant Biol. 2012, 12, 207–212. [Google Scholar] [CrossRef]
- Lee, J.P.; Sekhon, S.S.; Kim, J.H.; Kim, S.C.; Cho, B.K.; Ahn, J.Y.; Kim, Y.H. The Pine Wood Nematode Bursaphelenchus xylophilus and Molecular Diagnostic Methods. Mol. Cell Toxicol. 2020, 17, 1–13. [Google Scholar] [CrossRef]
- Kong, Q.Q.; Ding, X.L.; Chen, Y.F.; Ye, J. Comparison of Morphological Indexes and the Pathogenicity of Bursaphelenchus xylophilus in Northern and Southern China. Forests 2021, 12, 310. [Google Scholar] [CrossRef]
- Ikegami, M.; Jenkins Thomas, A.R. Estimate global risks of a forest disease under current and future climates using species distribution model and simple thermal model-pine wilt disease as a model case. Forest Ecol. Manag. 2018, 409, 343–352. [Google Scholar] [CrossRef]
- Yamaguchi, R.; Matsunaga, K.; Watanabe, A. Influence of temperature on pine wilt disease progression in Pinus thunbergii seedlings. Eur. J. Plant Pathol. 2020, 156, 581–590. [Google Scholar] [CrossRef]
- Li, M.; Li, H.; Ding, X.L.; Wang, L.; Wang, X.; Chen, F. The detection of pine wilt disease: A literature review. Int. J. Mol. Sci. 2022, 23, 10797. [Google Scholar] [CrossRef]
- Lim, W.; Choi, K.; Cho, W.; Chang, B.; Ko, D.W. Efficient dead pine tree detecting method in the forest damaged by pine wood nematode (Bursaphelenchus xylophilus) through utilizing unmanned aerial vehicles and deep learning-based object detection techniques. For. Sci. Technol. 2022, 18, 36–43. [Google Scholar] [CrossRef]
- Hussain, T.; Aslam, A.; Ozair, M.; Tasneem, F.; Gomez-Aguilar, J.F. Dynamical aspects of pine wilt disease and control measures. Chaos Solitons Fractals 2021, 145, 110764. [Google Scholar] [CrossRef]
- Hirao, T.; Matsunaga, K.; Hirakawa, H.; Shirasawa, k.; Isoda, K.; Mishima, K.; Tamura, M.; Watanabe, A. Construction of genetic linkage map and identification of a novel major locus for resistance to pine wood nematode in Japanese black pine (Pinus thunbergii). BMC Plant Biol. 2019, 19, 424. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, Y. Breeding project on resistance to the pine-wood nematode, An outline of the research and the achievement of the project for ten years. Bull. For. Tree Breed. Inst. 1989, 7, 1–84. [Google Scholar]
- Toda, T. Studies on the breeding for resistance to the pine wilt disease in Pinus densiflora and P. thunbergii. Bull For. Tree Breed Cent. 2004, 20, 83–217. [Google Scholar]
- Wu, X.Q.; Zhang, Y.; Chen, W.S. Resistance and histopathological observation of wilt-resistant Pinus thunbergia families from Japan to Bursaphelenchus xylophilus. Acta Phytopathol. Sin. 2008, 38, 44–50. [Google Scholar]
- Sun, T.; Wang, Y.; Zhu, L.; Wu, X.; Ye, J. Plant regeneration by somatic embryogenesis in Pinus thunbergii resistant to the pine wood nematode. Can. J. Forest Res. 2019, 49, 1604–1612. [Google Scholar] [CrossRef]
- Parker, J. Early Leads to Mechanisms of Plant Cultivar-Specific Disease Resistance. Plant Cell 2019, 31, 1410–1411. [Google Scholar] [CrossRef]
- Jacob, P.; Hige, J.; Dangl, J.L. Is localized acquired resistance the mechanism for effector-triggered disease resistance in plants? Nat. Plants 2023, 9, 1184–1190. [Google Scholar]
- Gaspar, D.; Trindade, C.; Usié, A.; Meireles, B.; Barbosa, P.; Fortes, A.M.; Pesquita, C.; Costa, R.L.; Ramos, A.M. Expression Profiling in Pinus pinaster in response to infection with the pine wood nematode Bursaphelenchus xylophilus. Forests 2017, 8, 279. [Google Scholar] [CrossRef]
- Neale, D.B.; Savolainen, O. Association genetics of complex traits in conifers. Trends Plant Sci. 2004, 9, 325–330. [Google Scholar] [CrossRef]
- Guevara, M.A.; Soto, A.; Collada, C.; Plomion, C.; Savolainen, O.; Neale, D.B.; Gonzalez-Martinez, S.C.; Cervera, M.T. Genomics applied to the study of adaptation in pine species. Invest. Agrar. Sist. Recur. For. 2005, 14, 292–306. [Google Scholar] [CrossRef][Green Version]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Parchman, T.L.; Geist, K.S.; Grahnen, J.A.; Benkman, C.W.; Buerkle, C.A. Transcriptome sequencing in an ecologically important tree species: Assembly, annotation, and marker discovery. BMC Genom. 2010, 11, 180. [Google Scholar] [CrossRef]
- Lee, I.H.; Han, H.; Koh, Y.; Kim, I.s.; Lee, S.W.; Shim, D. Comparative transcriptome analysis of Pinus densiflora following inoculation with pathogenic (Bursaphelenchus xylophilus) or non-pathogenic nematodes (B. thailandae). Sci. Rep. 2018, 9, 12180. [Google Scholar] [CrossRef] [PubMed]
- Modesto, I.; Sterck, L.; Arbona, V.; Gomaz-Cadenas, A.; Carrasquinho, I.; Van de Peer, Y. Insights into the mechanisms implicated in Pinus pinaster resistance to pine wood nematode. Front. Plant Sci. 2021, 12, 690857. [Google Scholar] [CrossRef]
- Liu, Q.; Wei, Y.; Xu, L.; Hao, Y.; Chen, X.; Zhou, Z. Transcriptomic profiling reveals differentially expressed genes associated with pine wood nematode resistance in masson pine (Pinus massoniana Lamb.). Sci. Rep. 2018, 7, 4693. [Google Scholar] [CrossRef] [PubMed]
- Menéndez-Gutiérrez, M.; Villar, L.; Díaz, R. Virulence of seven pathogenic Bursaphelenchus xylophilus isolates in Pinus pinaster and Pinus radiata seedlings and its relation with multiplication. For. Pathol. 2021, 51, e12677. [Google Scholar] [CrossRef]
- Sun, T.; Wang, Y.; Wu, X.; Ye, J.; Cheng, F. Promoting the application of Pinus thunbergii Parl. to enhance the growth and survival rates of post-germination somatic plantlets. BMC Plant Biol. 2023, 23, 195. [Google Scholar] [CrossRef]
- Ren, J.; Zhang, Y.; Wang, Y.; Li, C.; Zhang, X.; Liu, H.; Jiang, C.; Bian, Z.; Xu, R. Deletion of all three MAP kinase genes results in severe defects in stress responses and pathogenesis in Fusarium graminearum. Stress Biol. 2022, 2, 6. [Google Scholar] [CrossRef]
- Gao, X.; Zhao, S.; Xu, Q.L.; Xiao, J. Transcriptome responses of grafted Citrus sinensis plants to inoculation with the arbuscular mycorrhizal fungus Glomus versiforme. Trees. 2016, 30, 1073–1082. [Google Scholar] [CrossRef]
- Fernandes, L.B.; Ghag, S.B. Molecular insights into the jasmonate signaling and associated defense responses against wilt caused by Fusarium oxysporum. Plant Physiol. Biochem. 2022, 174, 22–34. [Google Scholar] [CrossRef]
- Fu, J.; Ren, R.; Jin, S.; Fang, R.; Wen, Z.; Yang, M.; Wang, X.; Lu, G.; Yang, Y.; Qi, J. Overexpression of a putative 12-oxophytodienoate reductase gene, EpOPR1, enhances acetylshikonin production in Echium plantagineum. Vitr. Cell. Dev. Biol. -Plant 2022, 58, 311–320. [Google Scholar] [CrossRef]
- Modesto, I.; Mendes, A.; Carrasquinho, I.; Miguel, C.M. Molecular defense response of pine trees (Pinus spp.) to the Parasitic Nematode Bursaphelenchus xylophilus. Cells 2022, 11, 3208. [Google Scholar] [CrossRef] [PubMed]
- Pyo, Y.; Moon, H.; Nugroho, A.B.D.; Yang, S.W.; Jung, I.L.; Kim, D.H. Transcriptome analysis revealed that jasmonic acid biosynthesis/signaling is involved in plant response to Strontium stress. Ecotoxicol. Environ. Saf. 2022, 237, 113552. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Pang, J.; Zhang, F.; Sun, L.; Yang, L.; Siddique, K.H.M. Transcriptomic and metabolomics-based analysis of key biological pathways reveals the role of lipid metabolism in response to salt stress in the root system of Brassica napus. Plant Growth Regul. 2022, 97, 127–141. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; Langer, S.; Carrasquinho, I.; Bergstrom, E.; Larson, T.; Thomas-Oates, J.; Antonio, C. Pinus pinaster Early Hormonal Defense Responses to Pinewood Nematode (Bursaphelenchus xylophilus) Infection. Metabolites 2021, 11, 227. [Google Scholar] [CrossRef]
- Tsuchiya, T.; Ohta, H.; Okawa, K.; Iwamatsu, A.; Shimada, H.; Masuda, T.; Takamiya, K. Cloning of chlorophyllase, the key enzyme in chlorophyll degradation: Finding of a lipase motif and the induction by methyl jasmonate. Proc. Natl. Acad. Sci. USA 1999, 96, 15362–15367. [Google Scholar] [CrossRef]
- Lv, J.; Zhang, Y.; Tang, W.; Chen, J.; Ge, Y.; Li, J. Concentration-dependent impacts of exogenous methyl jasmonate (MeJA) on chlorophyll degradation of apple fruit during ripening. Postharvest Biol. Technol. 2023, 203, 112398. [Google Scholar] [CrossRef]
- Völz, R.; Harris, W.; Hirt, H.; Lee, Y.H. ROS homeostasis mediated by MPK4 and SUMM2 determines synergid cell death. Nat. Commun. 2022, 13, 1746. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Minaker, S.; Roth, C.; Huang, S.; Hieter, P.; Liper, V.; Wiermer, M.; Li, X. An E4 ligase facilitates polyubiquitination of plant immune receptor resistance proteins in Arabidopsis. Plant Cell 2014, 26, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Liang, C.; Wang, S.; Hou, Y.; Gao, L.; Liu, L.; He, W.; Ma, W.; Mo, B.; Chen, X. The disease resistance protein SNC1 represses the biogenesis of microRNAs and phased siRNAs. Nat. Commun. 2018, 9, 5080. [Google Scholar] [CrossRef]
- Nitta, Y.; Qiu, Y.; Yaghmaiean, H.; Zhang, Q.; Huang, J.; Adams, K.; Zhang, Y. MEKK2 inhibits activation of MAP kinases in Arabidopsis. Plant J. 2020, 103, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.Q.; Dong, X. Systemic acquired resistance: Turning local infection into global defense. Annu. Rev. Plant Biol. 2013, 64, 839–863. [Google Scholar] [CrossRef]
- Shine, M.B.; Xiao, X.; Kachroo, P.; Kachroo, A. Signaling mechanisms underlying systemic acquired resistance to microbial pathogens. Plant Sci. 2019, 279, 81–86. [Google Scholar] [CrossRef]
- Mukhtar, M.S.; McCormack, M.E.; Argueso, C.T.; Pajerowska-Mukhtar, K.M. Pathogen Tactics to Manipulate Plant Cell Death. Curr. Biol. 2016, 26, 608–619. [Google Scholar] [CrossRef]
- Pitsili, E.; Phukan, U.J.; Coll, N.S. Cell Death in Plant Immunity. Cold Spring Harb. Perspect. Biol. 2020, 12, a036483. [Google Scholar] [CrossRef]
- Seo, J.S.; Diloknawarit, P.; Park, B.S.; Chua, N.H. ELF18-induced long noncoding RNA 1 evicts fibrillarin from mediator subunit to enhance ATHOGENESIS-RELATED GENE 1 (PR1) Expression. New Phytol. 2019, 221, 2067–2079. [Google Scholar] [CrossRef]
- Breen, S.; Williams, S.J.; Winterberg, B.; Kobe, b.; Solomon, P.S. Wheat PR-1 proteins are targeted by necrotrophic pathogen effector proteins. Plant J. 2016, 88, 13–25. [Google Scholar] [CrossRef]
- Bigeard, J.; Colcombet, J.; Hirt, H. Signaling mechanisms in pattern-triggered immunity (PTI). Mol. Plant 2015, 8, 521–539. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Xiong, D.; Schneiter, R.; Tian, C. The function of plant PR1 and other members of the CAP protein superfamily in plant–pathogen interactions. Mol. Plant Pathol. 2023, 24, 651–668. [Google Scholar] [CrossRef]
- Wu, Y.; Xing, D.; Ma, G.; Dai, X.; Gao, L.; Xia, T. A variable loop involved in the substrate selectivity of pinoresinol/lariciresinol reductase from Camellia sinensis. Phytochemistry 2019, 62, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Chattopadhyay, D. Lignin: The Building Block of Defense Responses to Stress in Plants. J. Plant Growth Regul. 2023. [CrossRef]
- Ott, D.S.; Yanchuk, A.D.; Huber, D.P.W.; Wallin, K.F. Genetic variation of lodgepole pine, Pinus contorta var. latifolia, chemical and physical defenses that affect mountain pine beetle, Dendroctonus ponderosae, attack and tree mortality. J. Chem. Ecol. 2011, 37, 1002–1012. [Google Scholar] [CrossRef]
- Clark, E.L.; Huber, D.P.W.; Carroll, A.L. The legacy of attack: Implications of high phloem resin monoterpene levels in lodgepole pine following mass attack by mountain pine beetle, Dendroctonus ponderosae Hopkins. Environ. Entomol. 2012, 42, 392–398. [Google Scholar] [CrossRef]
- Trapp, S.; Croteau, R. Defensive resin biosynthesis in conifers. Annu. Rev. Plant Biol. 2001, 52, 689–724. [Google Scholar] [CrossRef]
- Martin, D.M.; Bohlmann, J. Molecular biochemistry and genomics of terpenoid defenses in conifers. Rec. Adv. Phytochem. 2005, 39, 29–56. [Google Scholar]
- Tanaka, N.; Yoshida, S.; Takagi, H.; Terauchi, R.; Shimizu, A.; Fujiwara, T. Evidence for rice heading date 16 contribution to yield increase under low-nutrient conditions. Soil Sci. Plant Nutr. 2019, 65, 589–597. [Google Scholar] [CrossRef]
- Prisic, S.; Xu, M.; Wilderman, P.R. Rice Contains Two Disparate ent-Copalyl Diphosphate Synthases with Distinct Metabolic Functions. Plant Physiol. 2004, 36, 4228–4236. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).