
Identification of Potential miRNA Biomarkers to Detect Hydrocortisone Administration in Horses
 , ,
, ,
Abstract
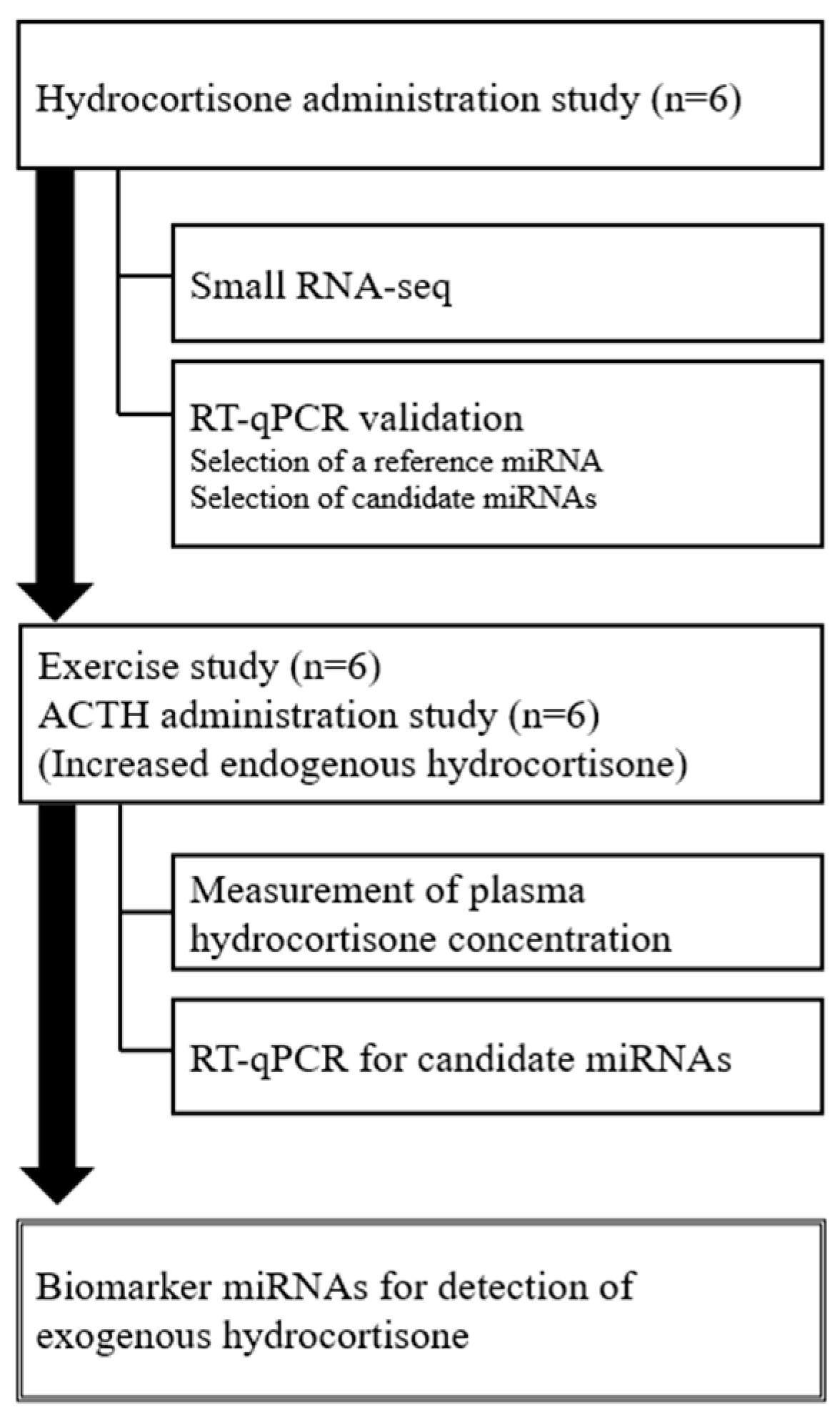
:1. Introduction
2. Results
2.1. Analysis of Circulating miRNA Profiles in Horses after Hydrocortisone Administration Using RNA-seq
2.2. Validation of miRNA Levels after Hydrocortisone Administration Using Reverse Transcription-Quantitative PCR (RT-qPCR)
2.3. Measurement of Plasma Hydrocortisone Concentration after Exercise and ACTH Administration
2.4. Analysis of miRNA Levels after Exercise and ACTH Administration Using RT-qPCR
3. Discussion
4. Materials and Methods
4.1. Animals and Sample Collection
4.2. RNA Extraction and Small RNA-seq
4.3. Bioinformatics Analyses
4.4. RT-qPCR
4.5. Evaluation of Reference miRNAs
4.6. Analysis of Hydrocortisone
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Sohel, M.H. Extracellular/circulating microRNAs: Release mechanisms, functions and challenges. Achiev. Life Sci. 2016, 10, 175–186. [Google Scholar] [CrossRef]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [PubMed]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J. Cell. Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef] [PubMed]
- Ho, P.T.B.; Clark, I.M.; Le, L.T.T. MicroRNA-based diagnosis and therapy. Int. J. Mol. Sci. 2022, 23, 7167. [Google Scholar] [CrossRef]
- Lv, Y.; Lu, C.; Ji, X.; Miao, Z.; Long, W.; Ding, H.; Lv, M. Roles of microRNAs in preeclampsia. J. Cell. Physiol. 2019, 234, 1052–1061. [Google Scholar] [CrossRef]
- Ono, K.; Okamoto, S.; Ninomiya, C.; Toji, N.; Kanazawa, T.; Ishiguro-Oonuma, T.; Takahashi, T.; Iga, K.; Kizaki, K. Analysis of circulating microRNA during early gestation in Japanese black cattle. Domest. Anim. Endocrinol. 2022, 79, 106706. [Google Scholar] [CrossRef]
- Leuenberger, N.; Schumacher, Y.O.; Pradervand, S.; Sander, T.; Saugy, M.; Pottgiesser, T. Circulating microRNAs as biomarkers for detection of autologous blood transfusion. PLoS ONE 2013, 8, e66309. [Google Scholar] [CrossRef]
- Salamin, O.; Jaggi, L.; Baume, N.; Robinson, N.; Saugy, M.; Leuenberger, N. Circulating microRNA-122 as potential biomarker for detection of testosterone abuse. PLoS ONE 2016, 11, e0155248. [Google Scholar] [CrossRef]
- Leuenberger, N.; Jan, N.; Pradervand, S.; Robinson, N.; Saugy, M. Circulating microRNAs as long-term biomarkers for the detection of erythropoiesis-stimulating agent abuse. Drug Test. Anal. 2011, 3, 771–776. [Google Scholar] [CrossRef]
- Kelly, B.N.; Haverstick, D.M.; Lee, J.K.; Thorner, M.O.; Vance, M.L.; Xin, W.; Bruns, D.E. Circulating microRNA as a biomarker of human growth hormone administration to patients. Drug Test. Anal. 2014, 6, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Igaz, I.; Nyírő, G.; Nagy, Z.; Butz, H.; Nagy, Z.; Perge, P.; Sahin, P.; Tóth, M.; Rácz, K.; Igaz, P.; et al. Analysis of Circulating microRNAs in vivo following Administration of dexamethasone and adrenocorticotropin. Int. J. Endocrinol. 2015, 2015, 589230. [Google Scholar] [CrossRef]
- Loup, B.; André, F.; Avignon, J.; Lhuaire, M.; Delcourt, V.; Barnabé, A.; Garcia, P.; Popot, M.A.; Bailly-Chouriberry, L. miRNAs detection in equine plasma by quantitative polymerase chain reaction for doping control: Assessment of blood sampling and study of eca-miR-144 as potential erythropoiesis stimulating agent biomarker. Drug Test. Anal. 2022, 14, 953–962. [Google Scholar] [CrossRef]
- Blackburn-Munro, G.; Blackburn-Munro, R. Pain in the brain: Are hormones to blame? Trends Endocrinol. Metab. 2003, 14, 20–27. [Google Scholar] [CrossRef]
- Jankord, R.; Herman, J.P. Limbic regulation of hypothalamo-pituitary-adrenocortical function during acute and chronic stress. Ann. N. Y. Acad. Sci. 2008, 1148, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Spiga, F.; Walker, J.J.; Terry, J.R.; Lightman, S.L. HPA axis-rhythms. Compr. Physiol. 2014, 4, 1273–1298. [Google Scholar]
- Leclere, M. Corticosteroids and immune suppressive therapies in horses. Vet. Clin. N. Am. Equine Pract. 2017, 33, 17–27. [Google Scholar] [CrossRef]
- Bohák, Z.; Szabó, F.; Beckers, J.F.; Melo de Sousa, N.; Kutasi, O.; Nagy, K.; Szenci, O. Monitoring the circadian rhythm of serum and salivary cortisol concentrations in the horse. Domest. Anim. Endocrinol. 2013, 45, 38–42. [Google Scholar] [CrossRef]
- Larsson, M.; Edqvist, L.E.; Ekman, L.; Persson, S. Plasma cortisol in the horse, diurnal rhythm and effects of exogenous ACTH. Acta Vet. Scand. 1979, 20, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, M.; Nagata, S.; Ishige, T.; Minamijima, Y.; Hirota, K.; Tozaki, T.; Kakoi, H.; Kizaki, K. Evaluation of the effect of glucocorticoid treatment on adrenocortical functions by monitoring endogenous hydrocortisone in horses. J. Vet. Med. Sci. 2023, 85, 647–652. [Google Scholar] [CrossRef]
- Lardizábal, M.N.; Nocito, A.L.; Daniele, S.M.; Ornella, L.A.; Palatnik, J.F.; Veggi, L.M. Reference genes for real-time PCR quantification of microRNAs and messenger RNAs in rat models of hepatotoxicity. PLoS ONE 2012, 7, e36323. [Google Scholar] [CrossRef]
- Pagacz, K.; Kucharski, P.; Smyczynska, U.; Grabia, S.; Chowdhury, D.; Fendler, W. A systemic approach to screening high-throughput RT-qPCR data for a suitable set of reference circulating miRNAs. BMC Genom. 2020, 21, 111. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Lee, S.; Hwang, S.; Yu, H.J.; Oh, D.; Choi, Y.J.; Kim, M.C.; Kim, Y.; Ryu, D.Y. Expression of microRNAs in horse plasma and their characteristic nucleotide composition. PLoS ONE 2016, 11, e0146374. [Google Scholar] [CrossRef]
- Tiedt, S.; Prestel, M.; Malik, R.; Schieferdecker, N.; Duering, M.; Kautzky, V.; Stoycheva, I.; Böck, J.; Northoff, B.H.; Klein, M.; et al. RNA-seq identifies circulating miR-125a-5p, miR-125b-5p, and miR-143-3p as potential biomarkers for acute ischemic stroke. Circ. Res. 2017, 121, 970–980. [Google Scholar] [CrossRef]
- Mestdagh, P.; Hartmann, N.; Baeriswyl, L.; Andreasen, D.; Bernard, N.; Chen, C.; Cheo, D.; D’Andrade, P.; DeMayo, M.; Dennis, L.; et al. Evaluation of quantitative miRNA expression platforms in the microRNA quality control (miRQC) study. Nat. Methods 2014, 11, 809–815. [Google Scholar] [CrossRef]
- Foreman, J.H.; Ferlazzo, A. Physiological responses to stress in the horse. Pferdeheilkunde Equine Med. 1996, 12, 401–404. [Google Scholar] [CrossRef]
- Schmidt, A.; Biau, S.; Möstl, E.; Becker-Birck, M.; Morillon, B.; Aurich, J.; Faure, J.M.; Aurich, C. Changes in cortisol release and heart rate variability in sport horses during long-distance road transport. Domest. Anim. Endocrinol. 2010, 38, 179–189. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhao, J.; Tuazon, J.P.; Borlongan, C.V.; Yu, G. MicroRNA-133a and myocardial infarction. Cell Transplant. 2019, 28, 831–838. [Google Scholar] [CrossRef]
- Liang, H.W.; Yang, X.; Wen, D.Y.; Gao, L.; Zhang, X.Y.; Ye, Z.H.; Luo, J.; Li, Z.Y.; He, Y.; Pang, Y.Y.; et al. Utility of miR-133a-3p as a diagnostic indicator for hepatocellular carcinoma: An investigation combined with GEO, TCGA, meta-analysis and bioinformatics. Mol. Med. Rep. 2018, 17, 1469–1484. [Google Scholar] [CrossRef]
- Xu, M.; Wang, Y.Z. miR-133a suppresses cell proliferation, migration and invasion in human lung cancer by targeting MMP-14. Oncol. Rep. 2013, 30, 1398–1404. [Google Scholar] [CrossRef]
- Li, Z.; Xu, W.; Ren, X.; Xu, J.; Chen, J. Puerarin promotes DUSP1 expression by regulating miR-133a-3p in breast cancer. Mol. Med. Rep. 2019, 19, 205–212. [Google Scholar] [CrossRef]
- Yu, A.M.; Tian, Y.; Tu, M.J.; Ho, P.Y.; Jilek, J.L. MicroRNA pharmacoepigenetics: Posttranscriptional regulation mechanisms behind variable drug disposition and strategy to develop more effective therapy. Drug Metab. Dispos. 2016, 44, 308–319. [Google Scholar] [CrossRef]
- Gould, D. Gene doping: Gene delivery for Olympic victory. Br. J. Clin. Pharmacol. 2013, 76, 292–298. [Google Scholar] [CrossRef]
- Sugasawa, T.; Nakano, T.; Fujita, S.I.; Matsumoto, Y.; Ishihara, G.; Aoki, K.; Yanazawa, K.; Ono, S.; Tamai, S.; Manevich, L.; et al. Proof of gene doping in a mouse model with a human erythropoietin gene transferred using an adenoviral vector. Genes 2021, 12, 1249. [Google Scholar] [CrossRef]
- Tozaki, T.; Ohnuma, A.; Kikuchi, M.; Ishige, T.; Kakoi, H.; Hirota, K.; Kusano, K.; Nagata, S. Microfluidic quantitative PCR detection of 12 transgenes from horse plasma for gene doping control. Genes 2020, 11, 457. [Google Scholar] [CrossRef]
- Nagata, S.; Takeda, F.; Kurosawa, M.; Mima, K.; Hiraga, A.; Kai, M.; Taya, K. Plasma adrenocorticotropin, cortisol and catecholamines response to various exercises. Equine Vet. J. Suppl. 1999, 31, 570–574. [Google Scholar] [CrossRef]
- Caloni, F.; Spotti, M.; Villa, R.; Mariani, C.; Montana, M.; Pompa, G. Hydrocortisone levels in the urine and blood of horses treated with ACTH. Equine Vet. J. 1999, 31, 273–276. [Google Scholar] [CrossRef]
- Kirchmeier, A.; van Herwaarden, A.E.; van der Kolk, J.H.; Sauer, F.J.; Gerber, V. Plasma steroid profiles before and after ACTH stimulation test in healthy horses. Domest. Anim. Endocrinol. 2020, 72, 106419. [Google Scholar] [CrossRef]
- Babraham Bioinformatics—FastQC A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 27 July 2023).
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [PubMed]
- Friedländer, M.R.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples 1 | Average Raw Reads | Average Clean Reads | Mapped Reads | Number of Novel miRNAs | Number of Known miRNAs |
|---|---|---|---|---|---|
| Pre-09 | 14,709,992 | 7,270,876 | 2,645,358 | 324 | 319 |
| Pre-10 | 11,895,794 | 6,370,296 | 2,855,883 | 338 | 316 |
| Pre-12 | 12,523,047 | 6,438,134 | 2,785,224 | 334 | 323 |
| Pre-15 | 17,089,367 | 9,692,756 | 4,379,242 | 353 | 316 |
| Pre-18 | 14,515,643 | 7,707,896 | 3,839,293 | 345 | 330 |
| Pre-21 | 16,575,098 | 9,078,428 | 4,830,487 | 353 | 333 |
| H-1 | 15,258,351 | 8,375,329 | 5,082,109 | 357 | 326 |
| H-3 | 15,244,208 | 8,685,837 | 5,436,409 | 353 | 330 |
| H-6 | 15,476,810 | 8,622,097 | 4,919,080 | 347 | 330 |
| H-9 | 16,308,131 | 9,678,295 | 5,057,211 | 348 | 331 |
| H-12 | 16,497,648 | 9,086,488 | 4,410,764 | 352 | 318 |
| H-24 | 14,614,984 | 8,399,419 | 4,391,252 | 342 | 308 |
| H-36 | 17,752,135 | 10,470,181 | 5,682,381 | 348 | 324 |
| H-48 | 16,278,917 | 9,159,001 | 5,045,230 | 340 | 330 |
| miRNA | cel-miR-39 | miR-191a | let-7g | miR-128 | miR-146a |
|---|---|---|---|---|---|
| SD 1 | 0.506565 | 0.70237 | 0.814206 | 0.718547 | 0.527002 |
| Coefficient of correlation 1 | 0.852 | 0.861 | 0.87 | 0.899 | 0.922 |
| miRNA 1 | Time after the Administration (h) | Small RNA-seq | RT-qPCR | |||
|---|---|---|---|---|---|---|
| Level 2 | log2 Fold-Change | Level 2 | log2 Fold-Change | p-Value | ||
| chr3-33188 | 12 | ↑ | 8.66 | ↑ | 0.78 | 0.028 |
| chrX-47614 | 48 | ↑ | 4.19 | ↑ | 1.10 | 0.028 |
| chr20-23348 | 12 | ↑ | 3.48 | - | −0.17 | 0.249 |
| chrX-47606 | 36 | ↑ | 3.34 | - | −0.04 | 0.753 |
| miR-133a | 3 | ↑ | 2.98 | ↑ | 1.25 | 0.028 |
| miR-206 | 9 | ↑ | 2.75 | - | −0.09 | 0.463 |
| miR-1 | 3 | ↑ | 2.63 | ↑ | 1.29 | 0.046 |
| chr3-32616 | 6 | ↑ | 2.48 | - | −0.12 | 0.249 |
| miR-122 | 48 | ↑ | 2.32 | ↑ | 2.88 | 0.028 |
| chr7-41424 | 9 | ↑ | 2.12 | - | 0.40 | 0.116 |
| chr3-33188 | 3 | ↓ | −11.37 | - | 0.13 | 0.345 |
| chr25-30007 | 12 | ↓ | −4.14 | ↓ | −2.69 | 0.028 |
| miR-532-3p | 9 | ↓ | −2.04 | - | 0.33 | 0.116 |
| miR-451 | 6 | ↓ | −1.93 | ↓ | −1.42 | 0.028 |
| miR-7 | 6 | ↓ | −1.80 | ↓ | −0.78 | 0.046 |
| miR-144 | 6 | ↓ | −1.69 | - | −0.22 | 0.249 |
| chr7-42879 | 12 | ↓ | −1.59 | ↓ | −2.90 | 0.028 |
| miR-142-3p | 36 | ↓ | −1.48 | - | −0.30 | 0.075 |
| let-7a | 48 | ↓ | −1.47 | - | −0.61 | 0.075 |
| miR-200b | 12 | ↓ | −1.46 | - | −0.43 | 0.173 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kikuchi, M.; Ishige, T.; Minamijima, Y.; Hirota, K.-i.; Nagata, S.-i.; Tozaki, T.; Kakoi, H.; Ishiguro-Oonuma, T.; Kizaki, K. Identification of Potential miRNA Biomarkers to Detect Hydrocortisone Administration in Horses. Int. J. Mol. Sci. 2023, 24, 14515. https://doi.org/10.3390/ijms241914515
Kikuchi M, Ishige T, Minamijima Y, Hirota K-i, Nagata S-i, Tozaki T, Kakoi H, Ishiguro-Oonuma T, Kizaki K. Identification of Potential miRNA Biomarkers to Detect Hydrocortisone Administration in Horses. International Journal of Molecular Sciences. 2023; 24(19):14515. https://doi.org/10.3390/ijms241914515
Chicago/Turabian StyleKikuchi, Mio, Taichiro Ishige, Yohei Minamijima, Kei-ichi Hirota, Shun-ichi Nagata, Teruaki Tozaki, Hironaga Kakoi, Toshina Ishiguro-Oonuma, and Keiichiro Kizaki. 2023. "Identification of Potential miRNA Biomarkers to Detect Hydrocortisone Administration in Horses" International Journal of Molecular Sciences 24, no. 19: 14515. https://doi.org/10.3390/ijms241914515