
Epigenetic Marks as Predictors of Metabolic Response to Bariatric Surgery: Validation from an Epigenome Wide Association Study
 , ,
, ,
Abstract
:1. Introduction
2. Results
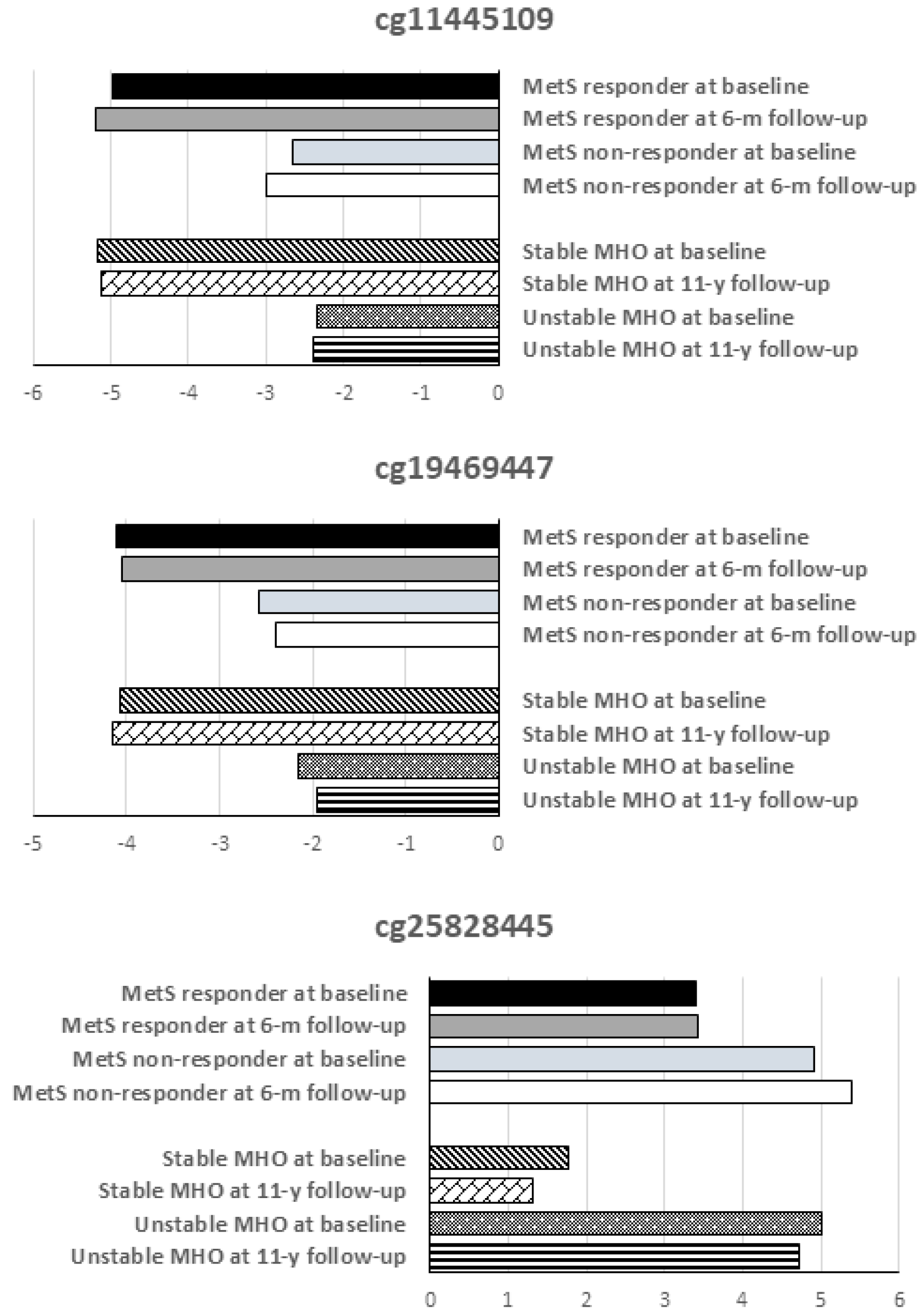
2.1. Validation of Differentially Methylated CpG Sites
2.2. Differentially Methylated Genes
2.3. Associations between DNA Methylation and Other Anthropometric and Biochemical Variables
3. Discussion
4. Material and Methods
4.1. Subjects
4.2. DNA Methylation Assay
4.3. Methylation Data Analysis
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Virani, S.S.; Alonso, A.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics-2020 Update: A Report from the American Heart Association. Circulation 2020, 141, E139–E596. [Google Scholar] [CrossRef] [PubMed]
- Fahed, G.; Aoun, L.; Bou Zerdan, M.; Allam, S.; Bou Zerdan, M.; Bouferraa, Y.; Assi, H.I. Metabolic Syndrome: Updates on Pathophysiology and Management in 2021. Int. J. Mol. Sci. 2022, 23, 786. [Google Scholar] [CrossRef] [PubMed]
- Ussar, S.; Griffin, N.W.; Bezy, O.; Fujisaka, S.; Vienberg, S.; Softic, S.; Deng, L.; Bry, L.; Gordon, J.I.; Kahn, C.R. Interactions between Gut Microbiota, Host Genetics and Diet Modulate the Predisposition to Obesity and Metabolic Syndrome. Cell Metab. 2015, 22, 516–530. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.A. Functions of DNA methylation: Islands, start sites, gene bodies and beyond. Nat. Rev. Genet. 2012, 13, 484–492. [Google Scholar] [CrossRef]
- Nuotio, M.-L.; Pervjakova, N.; Joensuu, A.; Karhunen, V.; Hiekkalinna, T.; Milani, L.; Kettunen, J.; Järvelin, M.-R.; Jousilahti, P.; Metspalu, A.; et al. An epigenome-wide association study of metabolic syndrome and its components. Sci. Rep. 2020, 10, 20567. [Google Scholar] [CrossRef]
- Akinyemiju, T.; Do, A.N.; Patki, A.; Aslibekyan, S.; Zhi, D.; Hidalgo, B.; Tiwari, H.K.; Absher, D.; Geng, X.; Arnett, D.K.; et al. Epigenome-wide association study of metabolic syndrome in African-American adults. Clin. Epigenetics 2018, 10, 49. [Google Scholar] [CrossRef]
- ElGendy, K.; Malcomson, F.C.; Bradburn, D.M.; Mathers, J.C. Effects of bariatric surgery on DNA methylation in adults: A systematic review and meta-analysis. Surg. Obes. Relat. Dis. 2020, 16, 128–136. [Google Scholar] [CrossRef]
- Faenza, M.; Benincasa, G.; Docimo, L.; Nicoletti, G.F.; Napoli, C. Clinical epigenetics and restoring of metabolic health in severely obese patients undergoing batriatric and metabolic surgery. Updates Surg. 2022, 74, 431–438. [Google Scholar] [CrossRef]
- Fraszczyk, E.; Luijten, M.; Spijkerman, A.M.W.; Snieder, H.; Wackers, P.F.K.; Bloks, V.W.; Nicoletti, C.F.; Nonino, C.B.; Crujeiras, A.B.; Buurman, W.A.; et al. The effects of bariatric surgery on clinical profile, DNA methylation, and ageing in severely obese patients. Clin. Epigenetics 2020, 12, 14. [Google Scholar] [CrossRef]
- Morcillo, S.; Macías-González, M.; Tinahones, F.J. The Effect of Metabolic and Bariatric Surgery on DNA Methylation Patterns. Curr. Atheroscler. Rep. 2017, 19, 40. [Google Scholar] [CrossRef]
- Nicoletti, C.F.; Nonino, C.B.; de Oliveira, B.A.P.; Pinhel, M.A.d.S.; Mansego, M.L.; Milagro, F.I.; Zulet, M.A.; Martinez, J.A. DNA Methylation and Hydroxymethylation Levels in Relation to Two Weight Loss Strategies: Energy-Restricted Diet or Bariatric Surgery. Obes. Surg. 2016, 26, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Coppedè, F.; Seghieri, M.; Stoccoro, A.; Santini, E.; Giannini, L.; Rossi, C.; Migliore, L.; Solini, A. DNA methylation of genes regulating appetite and prediction of weight loss after bariatric surgery in obese individuals. J. Endocrinol. Investig. 2019, 42, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Repiso, C.; Linares-Pineda, T.M.; Gonzalez-Jimenez, A.; Aguilar-Lineros, F.; Valdés, S.; Soriguer, F.; Rojo-Martínez, G.; Tinahones, F.J.; Morcillo, S. Epigenetic Biomarkers of Transition from Metabolically Healthy Obesity to Metabolically Unhealthy Obesity Phenotype: A Prospective Study. Int. J. Mol. Sci. 2021, 22, 10417. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Jiang, S.; Wang, J.; Renukuntla, J.; Sirimulla, S.; Chen, J. A comprehensive review of cytochrome P450 2E1 for xenobiotic metabolism. Drug Metab. Rev. 2019, 51, 178–195. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Lu, D.; Dong, W.; Zhang, L.; Zhang, X.; Quan, X.; Ma, C.; Lian, H.; Zhang, L. Expression of CYP2E1 increases oxidative stress and induces apoptosis of cardiomyocytes in transgenic mice. FEBS J. 2011, 278, 1484–1492. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Hall, S.D.; Maya, J.F.; Li, L.; Asghar, A.; Gorski, J.C. Diabetes mellitus increases the in vivo activity of cytochrome P450 2E1 in humans. Br. J. Clin. Pharmacol. 2003, 55, 77–85. [Google Scholar] [CrossRef]
- van Rongen, A.; Välitalo, P.A.J.; Peeters, M.Y.M.; Boerma, D.; Huisman, F.W.; van Ramshorst, B.; van Dongen, E.P.A.; Anker, J.N.v.D.; Knibbe, C.A.J. Morbidly Obese Patients Exhibit Increased CYP2E1-Mediated Oxidation of Acetaminophen. Clin. Pharmacokinet. 2016, 55, 833–847. [Google Scholar] [CrossRef]
- Zhang, L.; Tang, J.; Wang, Y.; Wang, X.; Wang, F. Association of CYP7A1 and CYP2E1 Polymorphisms with Type 2 Diabetes in the Chinese Han Populations. Pharmgenomics Pers. Med. 2022, 15, 843–855. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, L.; Xu, P.; Qin, X.; Wang, P.; Cheng, Y.; Yao, B.; Wang, X. Cytochrome P450 2E1 gene knockout or inhibition prevents obesity induced by high-fat diet via regulating energy expenditure. Biochem. Pharmacol. 2022, 202, 115160. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, T.; Wang, T.; Liu, X.; Hamada, K.; Sun, D.; Sun, Y.; Yang, Y.; Wang, J.; Takahashi, S.; et al. Crosstalk between CYP2E1 and PPAR α substrates and agonists modulate adipose browning and obesity. Acta Pharm. Sin. B 2022, 12, 2224–2238. [Google Scholar] [CrossRef]
- Kathirvel, E.; Morgan, K.; French, S.W.; Morgan, T.R. Overexpression of liver-specific cytochrome P4502E1 impairs hepatic insulin signaling in a transgenic mouse model of nonalcoholic fatty liver disease. Eur. J. Gastroenterol. Hepatol. 2009, 21, 973–983. [Google Scholar] [CrossRef] [PubMed]
- Witek, J.; Mohiuddin, S.S. Biochemistry, Pseudogenes; StatPearls: St. Petersburg, FL, USA, 2022. [Google Scholar]
- Keller, M.; Meir, A.Y.; Bernhart, S.H.; Gepner, Y.; Shelef, I.; Schwarzfuchs, D.; Tsaban, G.; Zelicha, H.; Hopp, L.; Müller, L.; et al. DNA methylation signature in blood mirrors successful weight-loss during lifestyle interventions: The CENTRAL trial. Genome Med. 2020, 12, 18. [Google Scholar] [CrossRef]
- Cleeman, J.I. Executive summary of the third report of the National Cholesterol Education Program (NCEP) expert panel on detection, evaluation, and treatment of high blood cholesterol in adults (adult treatment panel III). J. Am. Med. Assoc. 2001, 285, 2486–2497. [Google Scholar] [CrossRef]
- Aryee, M.J.; Jaffe, A.E.; Corrada-Bravo, H.; Ladd-Acosta, C.; Feinberg, A.P.; Hansen, K.D.; Irizarry, R.A. Minfi: A flexible and comprehensive Bioconductor package for the analysis of Infinium DNA methylation microarrays. Bioinformatics 2014, 30, 1363–1369. [Google Scholar] [CrossRef] [PubMed]
- Triche, T.J.; Weisenberger, D.J.; Van Den Berg, D.; Laird, P.W.; Siegmund, K.D. Low-level processing of Illumina Infinium DNA Methylation BeadArrays. Nucleic Acids Res. 2013, 41, e90. [Google Scholar] [CrossRef] [PubMed]
- Teschendorff, A.E.; Marabita, F.; Lechner, M.; Bartlett, T.; Tegner, J.; Gomez-Cabrero, D.; Beck, S. A beta-mixture quantile normalization method for correcting probe design bias in Illumina Infinium 450 k DNA methylation data. Bioinformatics 2013, 29, 189–196. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| MetS Non-Responder at Baseline | MetS Responder at Baseline | MetS Non-Responder 6 m after Surgery | MetS Responder 6 m after Surgery | |
|---|---|---|---|---|
| Sex (M/F) | 1/9 | 3/7 | ||
| Age (years) | 49.7 ± 10.29 | 47.7 ± 5.49 | ||
| Weight (kg) | 126.45 ± 22.91 | 125.32 ± 20.56 | 101.29 ± 15.91 | 94.59 ± 12.35 |
| BMI (kg/m2) | 51.54 ± 10.34 | 45.9 ± 5.03 | 41.29 ± 7.25 | 34.73 ± 2.82 # |
| Glucose (mg/dL) | 113.4 ± 18.77 | 117.1 ± 28.85 | 96.60 ± 14.85 | 89.1 ± 7.92 |
| Insulin (µUI/mL) | 19.84 ± 9.01 | 27.91 ± 18.63 | 10.80 ± 2.78 | 11.39 ± 11.18 |
| HOMA-IR | 5.6 ± 2.95 | 8.21 ± 5.55 | 2.53 ± 0.63 | 2.54 ± 2.61 |
| C-peptide (ng/mL) | 4.59 ± 1.47 | 4.31 ± 1.37 | 3.14 ± 0.59 | 2.52 ± 0.81 # |
| Cholesterol (mg/dL) | 188.3 ± 25.18 | 175.6 ± 37.76 | 207.50 ± 37.24 | 174.3 ± 29.77 # |
| Triglycerides (mg/dL) | 197.3 ± 45.59 | 137.4 ± 70.45 * | 153.60 ± 21.06 | 81.30 ± 19.39 # |
| HDL-cholesterol (mg/dL) | 38.6 ± 4.19 | 45.8 ± 8.95 | 43.20 ± 3.73 | 58.10 ± 12.83 # |
| LDL-cholesterol (mg/dL) | 111.18 ± 25.37 | 104.92 ± 31.39 | 131.26 ± 35.26 | 99.94 ± 24.88 |
| AST (U/L) | 28.5 ± 19.69 | 25.4 ± 13.38 | 17.56 ± 7.12 | 15.70 ± 3.77 |
| ALT (U/L) | 44.7 ± 26.06 | 39.3 ± 22.81 | 27.70 ± 8.90 | 21.40 ± 6.62 |
| GGT (U/L) | 46.8 ± 31.83 | 45.1 ± 17.46 | 36.00 ± 27.58 | 26.00 ± 14.53 |
| SBP (mm Hg) | 145.11 ± 24.4 | 137.8 ± 21.17 | 141.0 ± 17.45 | 124.4 ± 14.51 # |
| DBP (mm Hg) | 87.11 ± 13.14 | 83.4 ± 12.63 | 87.78 ± 11.55 | 78.6 ± 11.85 |
| Diabetes treatment (y/n) | 7/3 | 7/3 | 4/6 | 1/9 |
| HTA treatment (y/n) | 7/3 | 8/2 | 7/3 | 4/6 |
| Gene | KEGG Orthology | Differentially Methylated CpG Sites | Total Number of CpG Sites | % |
|---|---|---|---|---|
| RAP1GAP2 | Signaling and cellular processes | 21 | 88 | 23.86 |
| MUC2 | Membrane trafficking | 11 | 60 | 18.33 |
| ZFPM2 | Transcription factor | 11 | 41 | 26.83 |
| COG5 | Membrane trafficking | 8 | 63 | 12.70 |
| CYP2E1 | Metabolism | 8 | 20 | 40.00 |
| PLCZ1 | Membrane trafficking | 6 | 29 | 20.69 |
| SLC1A1 | Signaling and cellular processes | 6 | 25 | 24.00 |
| FRMD1 | Signaling and cellular processes | 5 | 42 | 11.90 |
| HLA-DRB1 | Signaling and cellular processes | 5 | 15 | 33.33 |
| ZBTB45 | Transcription factor | 5 | 27 | 18.52 |
| HLA-DQ2 | Signaling and cellular processes | 4 | 49 | 8.16 |
| HOOK3 | Signaling and cellular processes. Membrane trafficking | 4 | 35 | 11.43 |
| DENND1A | Membrane trafficking | 3 | 18 | 16.67 |
| TNFRSF4 | Signaling and cellular processes | 2 | 13 | 15.38 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez-Repiso, C.; Cantarero-Cuenca, A.; González-Jiménez, A.; Linares-Pineda, T.; Peña-Montero, N.; Ocaña-Wilhelmi, L.; Tinahones, F.J.; Morcillo, S. Epigenetic Marks as Predictors of Metabolic Response to Bariatric Surgery: Validation from an Epigenome Wide Association Study. Int. J. Mol. Sci. 2023, 24, 14778. https://doi.org/10.3390/ijms241914778
Gutiérrez-Repiso C, Cantarero-Cuenca A, González-Jiménez A, Linares-Pineda T, Peña-Montero N, Ocaña-Wilhelmi L, Tinahones FJ, Morcillo S. Epigenetic Marks as Predictors of Metabolic Response to Bariatric Surgery: Validation from an Epigenome Wide Association Study. International Journal of Molecular Sciences. 2023; 24(19):14778. https://doi.org/10.3390/ijms241914778
Chicago/Turabian StyleGutiérrez-Repiso, Carolina, Antonio Cantarero-Cuenca, Andrés González-Jiménez, Teresa Linares-Pineda, Nerea Peña-Montero, Luis Ocaña-Wilhelmi, Francisco J. Tinahones, and Sonsoles Morcillo. 2023. "Epigenetic Marks as Predictors of Metabolic Response to Bariatric Surgery: Validation from an Epigenome Wide Association Study" International Journal of Molecular Sciences 24, no. 19: 14778. https://doi.org/10.3390/ijms241914778