
Proton-Pump Inhibitors Suppress T Cell Response by Shifting Intracellular Zinc Distribution



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
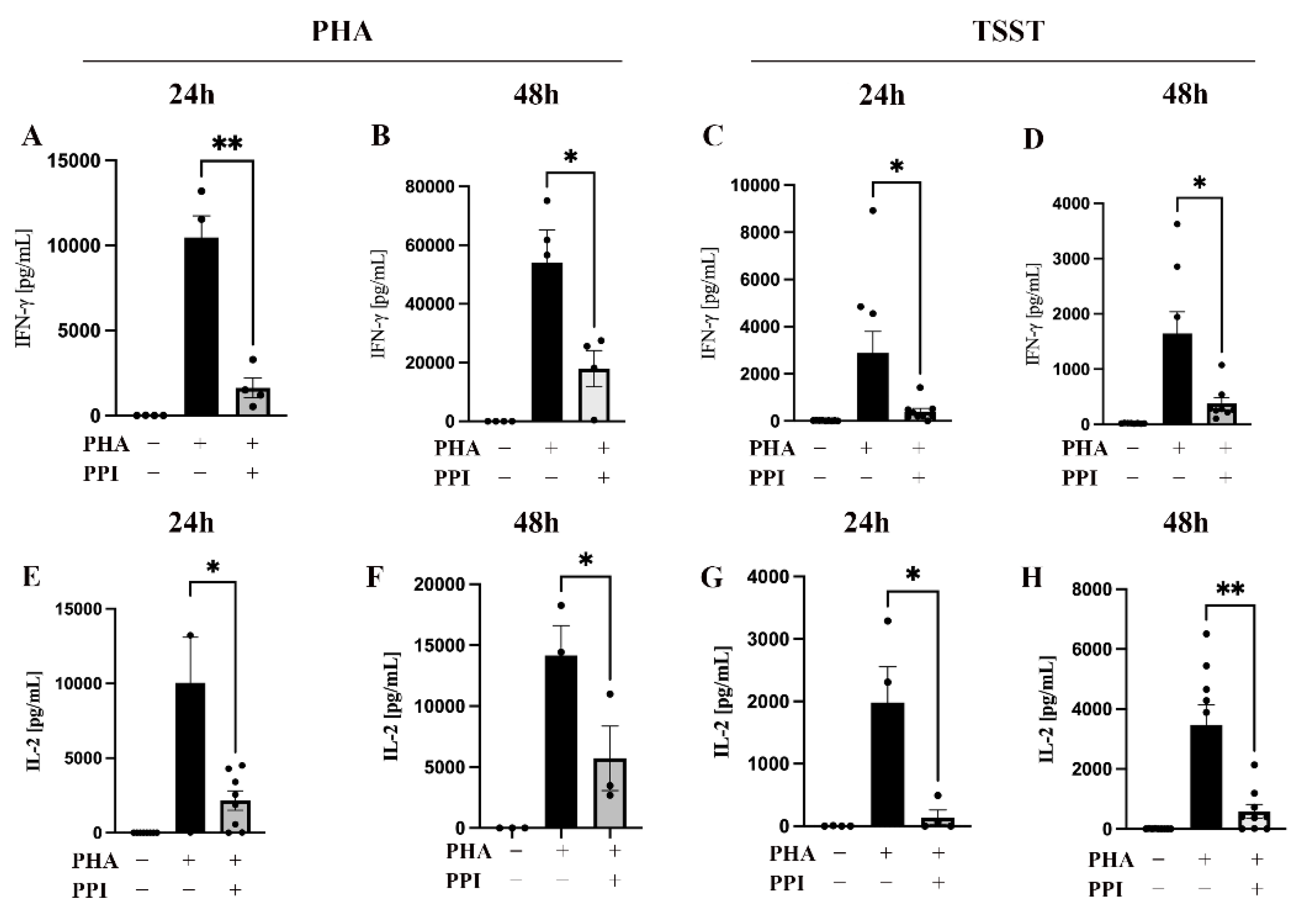
2.1. Effect of PPI on Cytokine Production
2.2. Effect of PPIs on the Distribution of Intracellular-Free Zinc
2.3. The Influence of PPIs on the Expression of Zip8
2.4. PPI Treatment Increased the Number of CREM Isoform
2.5. The Influence of PPIs on the Expression of pCREB
3. Discussion
4. Materials and Methods
4.1. Isolation of Human PBMC
4.2. Cell Culture and PPI-Treated Models
4.3. Flow Cytometric Measurement of Intracellular Free Zn2+ with FluoZin-3 AM and Zinpyr-1
4.4. IFN-γ and IL-2 Quantification
4.5. Western Blot
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nguyen, P.V.; Tamaz, R. Inappropriate Prescription of Proton Pump Inhibitors in a Community Setting. Can. J. Hosp. Pharm. 2018, 71, 267–271. [Google Scholar] [CrossRef] [Green Version]
- Kelly, O.B.; Dillane, C.; Patchett, S.E.; Harewood, G.C.; Murray, F.E. The Inappropriate Prescription of Oral Proton Pump Inhibitors in the Hospital Setting: A Prospective Cross-Sectional Study. Dig. Dis. Sci. 2015, 60, 2280–2286. [Google Scholar] [CrossRef] [PubMed]
- Maradey-Romero, C.; Fass, R. New and future drug development for gastroesophageal reflux disease. J. Neurogastroenterol. Motil. 2014, 20, 6–16. [Google Scholar] [CrossRef] [Green Version]
- Maes, M.L.; Fixen, D.R.; Linnebur, S.A. Adverse effects of proton-pump inhibitor use in older adults: A review of the evidence. Ther. Adv. Drug Saf. 2017, 8, 273–297. [Google Scholar] [CrossRef]
- De Roza, M.A.; Kai, L.; Kam, J.W.; Chan, Y.H.; Kwek, A.; Ang, T.L.; Hsiang, J.C. Proton pump inhibitor use increases mortality and hepatic decompensation in liver cirrhosis. World J. Gastroenterol. 2019, 25, 4933–4944. [Google Scholar] [CrossRef] [PubMed]
- Haas, C.M.; Maywald, M.; Goetzenich, A.; Stoppe, C.; Rink, L. Proton-pump inhibitors elevate infection rate in cardiothoracic surgery patients by influencing PMN function in vitro and in vivo. J. Leukoc. Biol. 2018, 103, 777–788. [Google Scholar] [CrossRef]
- Qin, B.D.; Jiao, X.D.; Zhou, X.C.; Shi, B.; Wang, J.; Liu, K.; Wu, Y.; Ling, Y.; Zang, Y.S. Effects of concomitant proton pump inhibitor use on immune checkpoint inhibitor efficacy among patients with advanced cancer. Oncoimmunology 2021, 10, 1929727. [Google Scholar] [CrossRef]
- Xia, B.; Yang, M.; Nguyen, L.H.; He, Q.; Zhen, J.; Yu, Y.; Di, M.; Qin, X.; Lu, K.; Kuo, Z.C.; et al. Regular Use of Proton Pump Inhibitor and the Risk of Inflammatory Bowel Disease: Pooled Analysis of 3 Prospective Cohorts. Gastroenterology 2021, 161, 1842–1852.e10. [Google Scholar] [CrossRef]
- Wessels, I.; Fischer, H.J.; Rink, L. Dietary and Physiological Effects of Zinc on the Immune System. Annu. Rev. Nutr. 2021, 41, 133–175. [Google Scholar] [CrossRef] [PubMed]
- Fraker, P.J.; King, L.E.; Laakko, T.; Vollmer, T.L. The dynamic link between the integrity of the immune system and zinc status. J. Nutr. 2000, 130, 1399S–1406S. [Google Scholar] [CrossRef] [Green Version]
- Kloubert, V.; Wessels, I.; Wolf, J.; Blaabjerg, K.; Janssens, V.; Hapala, J.; Wagner, W.; Rink, L. Zinc deficiency leads to reduced interleukin-2 production by active gene silencing due to enhanced CREMα expression in T cells. Clin. Nutr. 2021, 40, 3263–3278. [Google Scholar] [CrossRef]
- Miyai, T.; Hojyo, S.; Ikawa, T.; Kawamura, M.; Irié, T.; Ogura, H.; Hijikata, A.; Bin, B.H.; Yasuda, T.; Kitamura, H.; et al. Zinc transporter SLC39A10/ZIP10 facilitates antiapoptotic signaling during early B-cell development. Proc. Natl. Acad. Sci. USA 2014, 111, 11780–11785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Goldberg, J.M.; Lippard, S.J.; Palmer, A.E. Superiority of SpiroZin2 Versus FluoZin-3 for monitoring vesicular Zn2+ allows tracking of lysosomal Zn2+ pools. Sci. Rep. 2018, 8, 15034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marszałek, I.; Goch, W.; Bal, W. Ternary Zn(II) Complexes of Fluorescent Zinc Probes Zinpyr-1 and Zinbo-5 with the Low Molecular Weight Component of Exchangeable Cellular Zinc Pool. Inorg. Chem. 2019, 58, 14741–14751. [Google Scholar] [CrossRef]
- Panwar, A.; Wangchuk, J.; Kar, M.; Lodha, R.; Medigeshi, G.R. Dengue virus replication enhances labile zinc pools by modulation of ZIP8. Cell. Microbiol. 2021, 23, e13395. [Google Scholar] [CrossRef]
- Knoell, D.L.; Smith, D.; Bao, S.; Sapkota, M.; Wyatt, T.A.; Zweier, J.L.; Flury, J.; Borchers, M.T.; Knutson, M. Imbalance in zinc homeostasis enhances lung Tissue Loss following cigarette smoke exposure. J. Trace Elem. Med. Biol. 2020, 60, 126483. [Google Scholar] [CrossRef]
- Samuelson, D.R.; Haq, S.; Knoell, D.L. Divalent Metal Uptake and the Role of ZIP8 in Host Defense Against Pathogens. Front. Cell Dev. Biol. 2022, 10, 924820. [Google Scholar] [CrossRef]
- Aydemir, T.B.; Liuzzi, J.P.; McClellan, S.; Cousins, R.J. Zinc transporter ZIP8 (SLC39A8) and zinc influence IFN-gamma expression in activated human T cells. J. Leukoc. Biol. 2009, 86, 337–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, S.C.; Smith, D.R.; Dyavar, S.R.; Wyatt, T.A.; Samuelson, D.R.; Bailey, K.L.; Knoell, D.L. Critical Role of Zinc Transporter (ZIP8) in Myeloid Innate Immune Cell Function and the Host Response against Bacterial Pneumonia. J. Immunol. 2021, 207, 1357–1370. [Google Scholar] [CrossRef]
- Mocchegiani, E.; Fabris, N. Age-related thymus involution: Zinc reverses in vitro the thymulin secretion defect. Int. J. Immunopharmacol. 1995, 17, 745–749. [Google Scholar] [CrossRef]
- Hönscheid, A.; Rink, L.; Haase, H. T-lymphocytes: A target for stimulatory and inhibitory effects of zinc ions. Endocr. Metab. Immune Disord. Drug Targets 2009, 9, 132–144. [Google Scholar] [CrossRef]
- Baarz, B.R.; Laurentius, T.; Wolf, J.; Wessels, I.; Bollheimer, L.C.; Rink, L. Short-term zinc supplementation of zinc-deficient seniors counteracts CREMα - mediated IL-2 suppression. Immun. Ageing 2022, 19, 40. [Google Scholar] [CrossRef]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-gamma: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar] [CrossRef] [Green Version]
- Bendickova, K.; Fric, J. Roles of IL-2 in bridging adaptive and innate immunity, and as a tool for cellular immunotherapy. J. Leukoc. Biol. 2020, 108, 427–437. [Google Scholar] [CrossRef]
- Morgan, D.A.; Ruscetti, F.W.; Gallo, R. Selective in vitro growth of T lymphocytes from normal human bone marrows. Science 1976, 193, 1007–1008. [Google Scholar] [CrossRef] [PubMed]
- Molina, C.A.; Foulkes, N.S.; Lalli, E.; Sassone-Corsi, P. Inducibility and negative autoregulation of CREM: An alternative promoter directs the expression of ICER, an early response repressor. Cell 1993, 75, 875–886. [Google Scholar] [CrossRef]
- Luo, Z.; Li, M.; Li, T.W.; Lv, Z.; Ye, Z.; Cisneros, W.J.; Zhang, J.; Yuan, L.; Hultquist, J.F.; Migueles, S.A.; et al. Differential Expression of CREM/ICER Isoforms Is Associated with the Spontaneous Control of HIV Infection. mBio 2022, 13, e0197921. [Google Scholar] [CrossRef] [PubMed]
- Trifan, A.; Stanciu, C.; Girleanu, I.; Stoica, O.C.; Singeap, A.M.; Maxim, R.; Chiriac, S.A.; Ciobica, A.; Boiculese, L. Proton pump inhibitors therapy and risk of Clostridium difficile infection: Systematic review and meta-analysis. World J. Gastroenterol. 2017, 23, 6500–6515. [Google Scholar] [CrossRef]
- Cao, F.; Chen, C.X.; Wang, M.; Liao, H.R.; Wang, M.X.; Hua, S.Z.; Huang, B.; Xiong, Y.; Zhang, J.Y.; Xu, Y.L. Updated meta-analysis of controlled observational studies: Proton-pump inhibitors and risk of Clostridium difficile infection. J. Hosp. Infect. 2018, 98, 4–13. [Google Scholar] [CrossRef]
- Licastro, F.; Davis, L.J.; Morini, M.C. Lectins and superantigens: Membrane interactions of these compounds with T lymphocytes affect immune responses. Int. J. Biochem. 1993, 25, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Driessen, C.; Hirv, K.; Kirchner, H.; Rink, L. Zinc regulates cytokine induction by superantigens and lipopolysaccharide. Immunology 1995, 84, 272–277. [Google Scholar] [PubMed]
- Bjorkman, P.J.; Saper, M.A.; Samraoui, B.; Bennett, W.S.; Strominger, J.L.; Wiley, D.C. The foreign antigen binding site and T cell recognition regions of class I histocompatibility antigens. Nature 1987, 329, 512–518. [Google Scholar] [CrossRef]
- Chilson, O.P.; Kelly-Chilson, A.E. Mitogenic lectins bind to the antigen receptor on human lymphocytes. Eur. J. Immunol. 1989, 19, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Roetzer, A.; Stich, N.; Model, N.; Schwameis, M.; Firbas, C.; Jilma, B.; Eibl, M.M. High Titer Persistent Neutralizing Antibodies Induced by TSST-1 Variant Vaccine Against Toxic Shock Cytokine Storm. Toxins 2020, 12, 640. [Google Scholar] [CrossRef]
- Tanaka, S.; Akaishi, E.; Hosaka, K.; Okamura, S.; Kubohara, Y. Zinc ions suppress mitogen-activated interleukin-2 production in Jurkat cells. Biochem. Biophys. Res. Commun. 2005, 335, 162–167. [Google Scholar] [CrossRef]
- Cheng, Y.; Chen, H. Aberrance of Zinc Metalloenzymes-Induced Human Diseases and Its Potential Mechanisms. Nutrients 2021, 13, 4456. [Google Scholar] [CrossRef] [PubMed]
- Vallee, B.L.; Auld, D.S. Functional zinc-binding motifs in enzymes and DNA-binding proteins. Faraday Discuss. 1992, 47–65. [Google Scholar] [CrossRef] [PubMed]
- Maret, W.; Jacob, C.; Vallee, B.L.; Fischer, E.H. Inhibitory sites in enzymes: Zinc removal and reactivation by thionein. Proc. Natl. Acad. Sci. USA 1999, 96, 1936–1940. [Google Scholar] [CrossRef] [Green Version]
- Pan, C.Y.; Lin, F.Y.; Kao, L.S.; Huang, C.C.; Liu, P.S. Zinc oxide nanoparticles modulate the gene expression of ZnT1 and ZIP8 to manipulate zinc homeostasis and stress-induced cytotoxicity in human neuroblastoma SH-SY5Y cells. PLoS ONE 2020, 15, e0232729. [Google Scholar] [CrossRef] [PubMed]
- Laoide, B.M.; Foulkes, N.S.; Schlotter, F.; Sassone-Corsi, P. The functional versatility of CREM is determined by its modular structure. EMBO J. 1993, 12, 1179–1191. [Google Scholar] [CrossRef]
- Xu, W.D.; Zhang, Y.J.; Wang, W.; Li, R.; Pan, H.F.; Ye, D.Q. Role of CREM in systemic lupus erythematosus. Cell. Immunol. 2012, 276, 10–15. [Google Scholar] [CrossRef]
- Kuttkat, N.; Mohs, A.; Ohl, K.; Hooiveld, G.; Longerich, T.; Tenbrock, K.; Cubero, F.J.; Trautwein, C. Hepatic overexpression of cAMP-responsive element modulator α induces a regulatory T-cell response in a murine model of chronic liver disease. Gut 2017, 66, 908–919. [Google Scholar] [CrossRef]
- Jiang, Y.G.; Fang, H.Y.; Pang, W.; Liu, J.; Lu, H.; Ma, Q.; Fang, H.T. Depressed hippocampal MEK/ERK phosphorylation correlates with impaired cognitive and synaptic function in zinc-deficient rats. Nutr. Neurosci. 2011, 14, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Yuan, L.; He, W.; Yang, X. Zinc ions regulate opening of tight junction favouring efflux of macromolecules via the GSK3β/snail-mediated pathway. Metallomics 2018, 10, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Akaishi, E.; Abe, Y.; Ishikawa, R.; Tanaka, S.; Hosaka, K.; Kubohara, Y. Zinc inhibits calcineurin activity in vitro by competing with nickel. Biochem. Biophys. Res. Commun. 2003, 307, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Haase, H.; Hebel, S.; Engelhardt, G.; Rink, L. Flow cytometric measurement of labile zinc in peripheral blood mononuclear cells. Anal. Biochem. 2006, 352, 222–230. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Jakobs, J.; Rink, L. Proton-Pump Inhibitors Suppress T Cell Response by Shifting Intracellular Zinc Distribution. Int. J. Mol. Sci. 2023, 24, 1191. https://doi.org/10.3390/ijms24021191
Liu W, Jakobs J, Rink L. Proton-Pump Inhibitors Suppress T Cell Response by Shifting Intracellular Zinc Distribution. International Journal of Molecular Sciences. 2023; 24(2):1191. https://doi.org/10.3390/ijms24021191
Chicago/Turabian StyleLiu, Wenlei, Jana Jakobs, and Lothar Rink. 2023. "Proton-Pump Inhibitors Suppress T Cell Response by Shifting Intracellular Zinc Distribution" International Journal of Molecular Sciences 24, no. 2: 1191. https://doi.org/10.3390/ijms24021191
APA StyleLiu, W., Jakobs, J., & Rink, L. (2023). Proton-Pump Inhibitors Suppress T Cell Response by Shifting Intracellular Zinc Distribution. International Journal of Molecular Sciences, 24(2), 1191. https://doi.org/10.3390/ijms24021191