
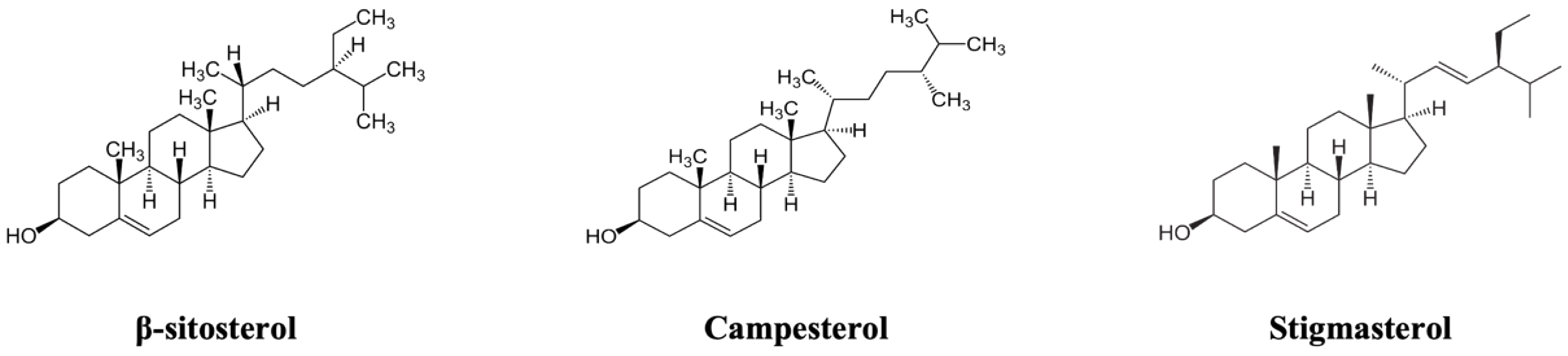
A Mixture of Dietary Plant Sterols at Nutritional Relevant Serum Concentration Inhibits Extrinsic Pathway of Eryptosis Induced by Cigarette Smoke Extract

 ,
,  ,
,  ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
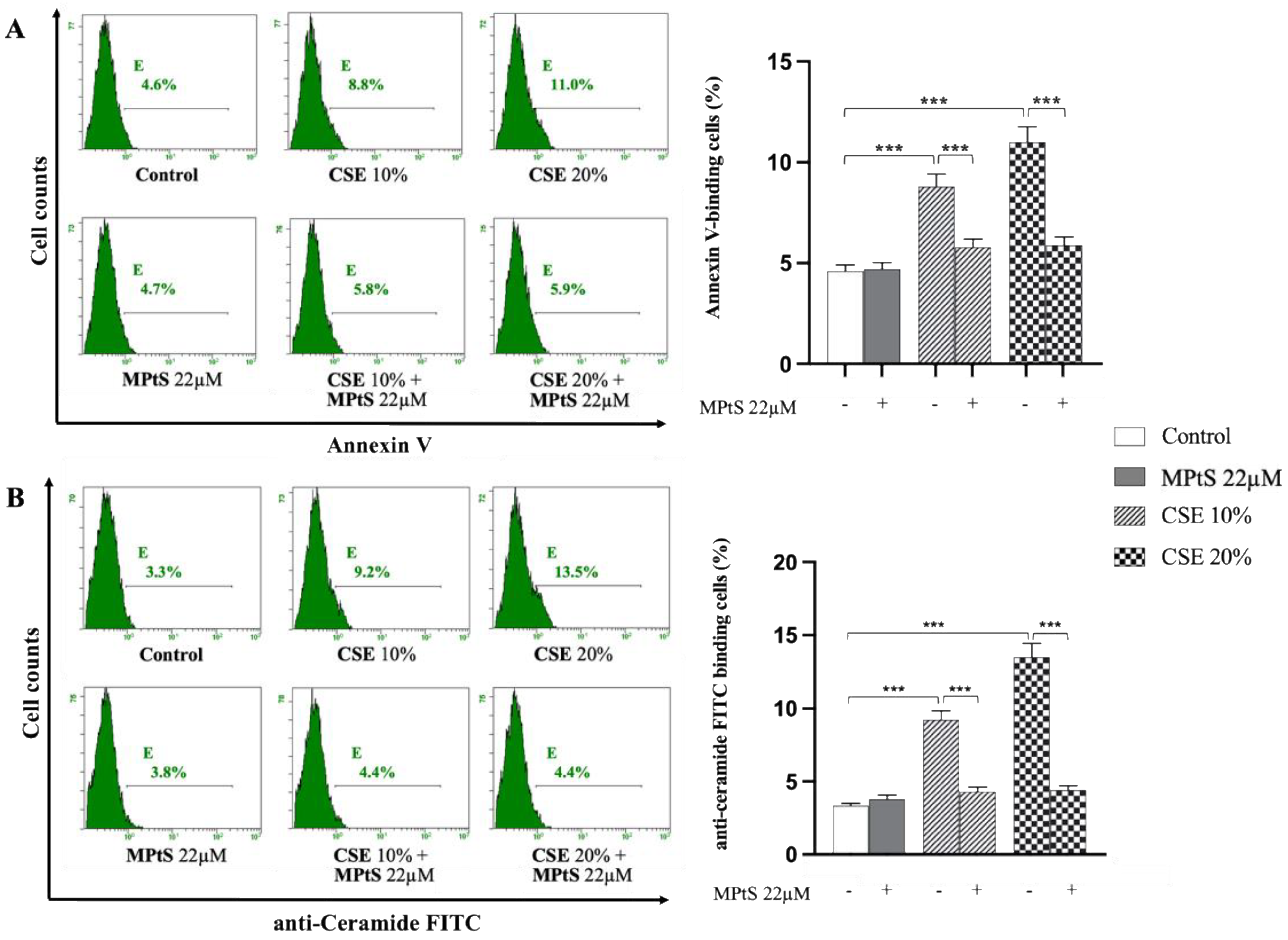
2.1. MPtS Decreases PS Exposure and Ceramide Production in CSE-Induced Eryptosis
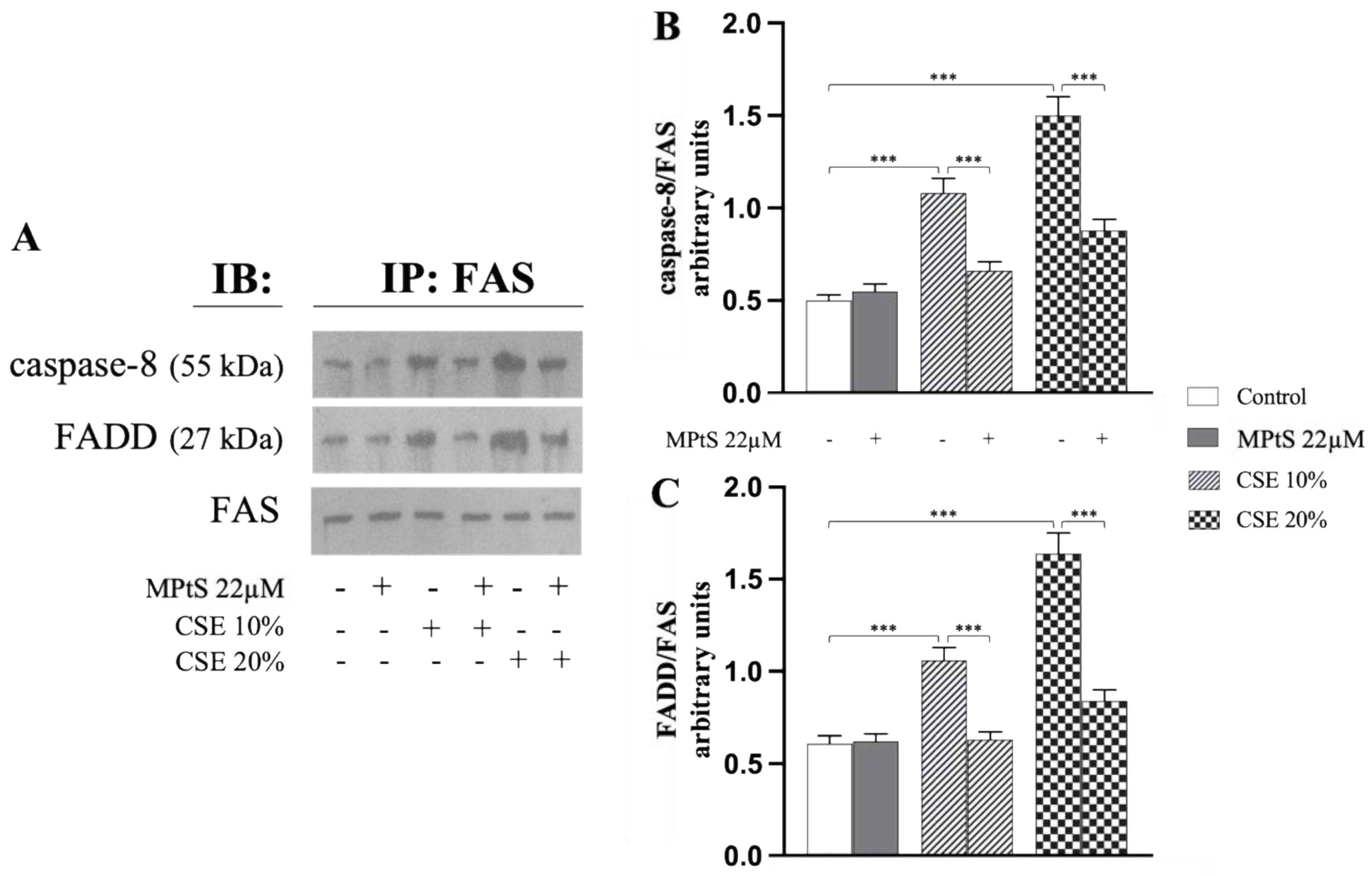
2.2. MPtS Inhibits CSE-Induced Extrinsic Eryptosis
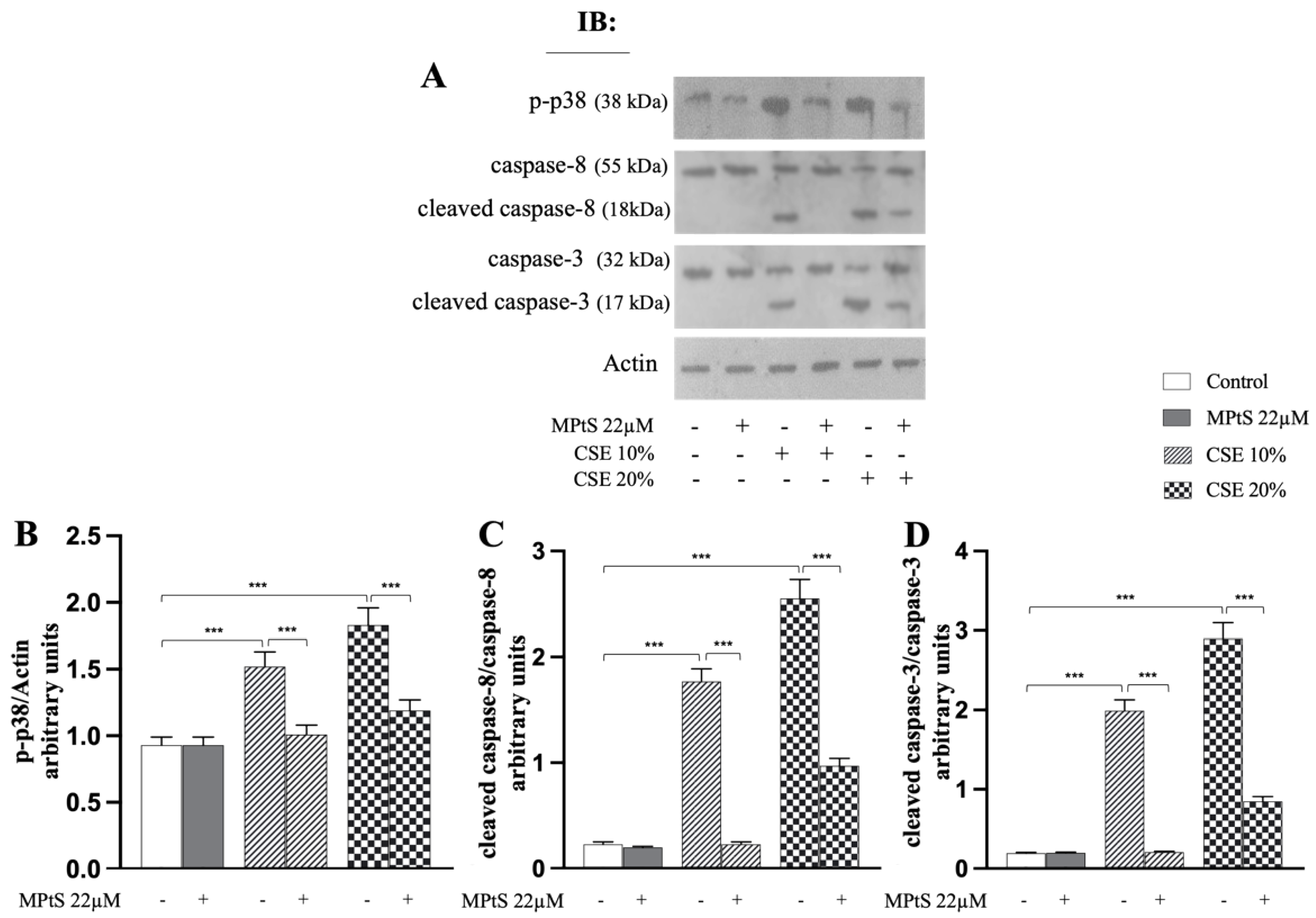
2.3. MPtS Inhibits Caspase 8/Caspase 3 Cleavage and Phosphorylation of p38 MAPK in CSE-Induced Extrinsic Eryptosis
3. Discussion
4. Materials and Methods
4.1. Preparation of CSE
4.2. Preparation of MPtS
4.3. Red Blood Cells and Treatment
4.4. Measurement of PS Externalization
4.5. Measurement of Ceramide
4.6. Immunoprecipitation
4.7. Western Blotting
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- World Health Organization. World Health Organization Global Report on Trends in Prevalence of Tobacco Smoking; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Blann, A.D.; McCollum, C.N. Adverse Influence of Cigarette Smoking on the Endothelium. Thromb. Haemost. 1993, 70, 707–711. [Google Scholar] [CrossRef] [PubMed]
- El-Mahdy, M.A.; Abdelghany, T.M.; Hemann, C.; Ewees, M.G.; Mahgoup, E.M.; Eid, M.S.; Shalaan, M.T.; Alzarie, Y.A.; Zweier, J.L. Chronic Cigarette Smoke Exposure Triggers a Vicious Cycle of Leukocyte and Endothelial-Mediated Oxidant Stress That Results in Vascular Dysfunction. Am. J. Physiol. Heart Circ. Physiol. 2020, 319, H51–H65. [Google Scholar] [CrossRef] [PubMed]
- Nowak, J.; Murray, J.J.; Oates, J.A.; FitzGerald, G.A. Biochemical Evidence of a Chronic Abnormality in Platelet and Vascular Function in Healthy Individuals Who Smoke Cigarettes. Circulation 1987, 76, 6–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attanzio, A.; Frazzitta, A.; Vasto, S.; Tesoriere, L.; Pintaudi, A.M.; Livrea, M.A.; Cilla, A.; Allegra, M. Increased Eryptosis in Smokers Is Associated with the Antioxidant Status and C-Reactive Protein Levels. Toxicology 2019, 411, 43–48. [Google Scholar] [CrossRef]
- Celermajer, D.S.; Sorensen, K.E.; Georgakopoulos, D.; Bull, C.; Thomas, O.; Robinson, J.; Deanfield, J.E. Cigarette Smoking Is Associated with Dose-Related and Potentially Reversible Impairment of Endothelium-Dependent Dilation in Healthy Young Adults. Circulation 1993, 88, 2149–2155. [Google Scholar] [CrossRef] [Green Version]
- Cilla, A.; López-García, G.; Collado-Díaz, V.; Amparo Blanch-Ruiz, M.; Garcia-Llatas, G.; Barberá, R.; Martinez-Cuesta, M.A.; Real, J.T.; Álvarez, Á.; Martínez-Hervás, S. Hypercholesterolemic Patients Have Higher Eryptosis and Erythrocyte Adhesion to Human Endothelium Independently of Statin Therapy. Int. J. Clin. Pract. 2021, 75, e14771. [Google Scholar] [CrossRef]
- Lang, F.; Qadri, S.M. Mechanisms and Significance of Eryptosis, the Suicidal Death of Erythrocytes. Blood Purif. 2012, 33, 125–130. [Google Scholar] [CrossRef]
- Qadri, S.M.; Bissinger, R.; Solh, Z.; Oldenborg, P.-A. Eryptosis in Health and Disease: A Paradigm Shift towards Understanding the (Patho)Physiological Implications of Programmed Cell Death of Erythrocytes. Blood Rev. 2017, 31, 349–361. [Google Scholar] [CrossRef]
- Lang, F.; Bissinger, R.; Abed, M.; Artunc, F. Eryptosis—The Neglected Cause of Anemia in End Stage Renal Disease. Kidney Blood Press. Res. 2017, 42, 749–760. [Google Scholar] [CrossRef]
- Restivo, I.; Attanzio, A.; Tesoriere, L.; Allegra, M. Suicidal Erythrocyte Death in Metabolic Syndrome. Antioxidants 2021, 10, 154. [Google Scholar] [CrossRef]
- Restivo, I.; Attanzio, A.; Tesoriere, L.; Allegra, M.; Garcia-Llatas, G.; Cilla, A. Anti-Eryptotic Activity of Food-Derived Phytochemicals and Natural Compounds. IJMS 2022, 23, 3019. [Google Scholar] [CrossRef] [PubMed]
- Restivo, I.; Attanzio, A.; Giardina, I.C.; Di Gaudio, F.; Tesoriere, L.; Allegra, M. Cigarette Smoke Extract Induces P38 MAPK-Initiated, Fas-Mediated Eryptosis. Int. J. Mol. Sci. 2022, 23, 14730. [Google Scholar] [CrossRef]
- Statistical office of the European Union (EUROSTAT). Causes of Death—Deats by Country of Residence and Occurrence (HLTH_CD_ARO); Statistical office of the European Union (EUROSTAT): Luxembourg, 2022. [Google Scholar]
- Gylling, H.; Plat, J.; Turley, S.; Ginsberg, H.N.; Ellegård, L.; Jessup, W.; Jones, P.J.; Lütjohann, D.; Maerz, W.; Masana, L.; et al. Plant sterols and plant stanols in the management of dyslipidaemia and prevention of cardiovascular disease. Atherosclerosis 2014, 232, 346–360. [Google Scholar] [CrossRef] [PubMed]
- Danesi, F.; Ferioli, F.; Caboni, M.F.; Boschetti, E.; Di Nunzio, M.; Verardo, V.; Valli, V.; Astolfi, A.; Pession, A.; Bordoni, A. Phytosterol supplementation reduces metabolic activity and slows cell growth in cultured rat cardiomyocytes. Br. J. Nutr. 2011, 106, 540–548. [Google Scholar] [CrossRef] [Green Version]
- Salehi, B.; Quispe, C.; Sharifi-Rad, J.; Cruz-Martins, N.; Nigam, M.; Mishra, A.P.; Konovalov, D.A.; Orobinskaya, V.; Abu-Reidah, I.M.; Zam, W.; et al. Phytosterols: From Preclinical Evidence to Potential Clinical Applications. Front. Pharmacol. 2020, 11, 599959. [Google Scholar] [CrossRef]
- Garcia-Llatas, G.; Cilla, A.; Alegría, A.; Lagarda, M.J. Bioavailability of Plant Sterol-Enriched Milk-Based Fruit Beverages: In Vivo and in Vitro Studies. J. Funct. Foods 2015, 14, 44–50. [Google Scholar] [CrossRef]
- Alvarez-Sala, A.; López-García, G.; Attanzio, A.; Tesoriere, L.; Cilla, A.; Barberá, R.; Alegría, A. Effects of Plant Sterols or β-Cryptoxanthin at Physiological Serum Concentrations on Suicidal Erythrocyte Death. J. Agric. Food Chem. 2018, 66, 1157–1166. [Google Scholar] [CrossRef]
- Allen, T.M.; Williamson, P.; Schlegel, R.A. Phosphatidylserine as a Determinant of Reticuloendothelial Recognition of Liposome Models of the Erythrocyte Surface. Proc. Natl. Acad. Sci. USA 1988, 85, 8067–8071. [Google Scholar] [CrossRef] [Green Version]
- Schroit, A.J.; Madsen, J.W.; Tanaka, Y. In Vivo Recognition and Clearance of Red Blood Cells Containing Phosphatidylserine in Their Plasma Membranes. J. Biol. Chem. 1985, 260, 5131–5138. [Google Scholar] [CrossRef]
- Boas, F.E.; Forman, L.; Beutler, E. Phosphatidylserine Exposure and Red Cell Viability in Red Cell Aging and in Hemolytic Anemia. Proc. Natl. Acad. Sci. USA 1998, 95, 3077–3081. [Google Scholar] [CrossRef]
- Lang, E.; Bissinger, R.; Gulbins, E.; Lang, F. Ceramide in the Regulation of Eryptosis, the Suicidal Erythrocyte Death. Apoptosis 2015, 20, 758–767. [Google Scholar] [CrossRef] [PubMed]
- Kuo, W.-H.; Chen, J.-H.; Lin, H.-H.; Chen, B.-C.; Hsu, J.-D.; Wang, C.-J. Induction of Apoptosis in the Lung Tissue from Rats Exposed to Cigarette Smoke Involves P38/JNK MAPK Pathway. Chem. Biol. Interact. 2005, 155, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Rubiolo, C.; Piazzolla, D.; Meissl, K.; Beug, H.; Huber, J.C.; Kolbus, A.; Baccarini, M. A Balance between Raf-1 and Fas Expression Sets the Pace of Erythroid Differentiation. Blood 2006, 108, 152–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatidis, S.; Zelenak, C.; Fajol, A.; Lang, E.; Jilani, K.; Michael, D.; Qadri, S.M.; Lang, F. P38 MAPK Activation and Function Following Osmotic Shock of Erythrocytes. Cell Physiol. Biochem. 2011, 28, 1279–1286. [Google Scholar] [CrossRef]
- Tobiume, K.; Matsuzawa, A.; Takahashi, T.; Nishitoh, H.; Morita, K.; Takeda, K.; Minowa, O.; Miyazono, K.; Noda, T.; Ichijo, H. ASK1 Is Required for Sustained Activations of JNK/P38 MAP Kinases and Apoptosis. EMBO Rep. 2001, 2, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liang, J.; Jiang, H.; Qian, M.; Zhao, W.; Bai, W. Protective Effect of Sterols Extracted from Lotus Plumule on Ethanol-Induced Injury in GES-1 Cells in Vitro. Food Funct. 2021, 12, 12659–12670. [Google Scholar] [CrossRef]
- Xu, J.; Xiao, C.; Xu, H.; Yang, S.; Chen, Z.; Wang, H.; Zheng, B.; Mao, B.; Wu, X. Anti-Inflammatory Effects of Ganoderma Lucidum Sterols via Attenuation of the P38 MAPK and NF-ΚB Pathways in LPS-Induced RAW 264.7 Macrophages. Food Chem. Toxicol. 2021, 150, 112073. [Google Scholar] [CrossRef]
- Carnevali, S.; Petruzzelli, S.; Longoni, B.; Vanacore, R.; Barale, R.; Cipollini, M.; Scatena, F.; Paggiaro, P.; Celi, A.; Giuntini, C. Cigarette Smoke Extract Induces Oxidative Stress and Apoptosis in Human Lung Fibroblasts. Am. J. Physiol. Lung Cell. Mol. Physiol. 2003, 284, L955–L963. [Google Scholar] [CrossRef] [Green Version]
- Allegra, M.; Restivo, I.; Fucarino, A.; Pitruzzella, A.; Vasto, S.; Livrea, M.A.; Tesoriere, L.; Attanzio, A. Proeryptotic Activity of 4-Hydroxynonenal: A New Potential Physiopathological Role for Lipid Peroxidation Products. Biomolecules 2020, 10, 770. [Google Scholar] [CrossRef]
- Restivo, I.; Tesoriere, L.; Frazzitta, A.; Livrea, M.A.; Attanzio, A.; Allegra, M. Anti-Proliferative Activity of A Hydrophilic Extract of Manna from Fraxinus Angustifolia Vahl through Mitochondrial Pathway-Mediated Apoptosis and Cell Cycle Arrest in Human Colon Cancer Cells. Molecules 2020, 25, E5055. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Restivo, I.; Attanzio, A.; Tesoriere, L.; Allegra, M.; Garcia-Llatas, G.; Cilla, A. A Mixture of Dietary Plant Sterols at Nutritional Relevant Serum Concentration Inhibits Extrinsic Pathway of Eryptosis Induced by Cigarette Smoke Extract. Int. J. Mol. Sci. 2023, 24, 1264. https://doi.org/10.3390/ijms24021264
Restivo I, Attanzio A, Tesoriere L, Allegra M, Garcia-Llatas G, Cilla A. A Mixture of Dietary Plant Sterols at Nutritional Relevant Serum Concentration Inhibits Extrinsic Pathway of Eryptosis Induced by Cigarette Smoke Extract. International Journal of Molecular Sciences. 2023; 24(2):1264. https://doi.org/10.3390/ijms24021264
Chicago/Turabian StyleRestivo, Ignazio, Alessandro Attanzio, Luisa Tesoriere, Mario Allegra, Guadalupe Garcia-Llatas, and Antonio Cilla. 2023. "A Mixture of Dietary Plant Sterols at Nutritional Relevant Serum Concentration Inhibits Extrinsic Pathway of Eryptosis Induced by Cigarette Smoke Extract" International Journal of Molecular Sciences 24, no. 2: 1264. https://doi.org/10.3390/ijms24021264
APA StyleRestivo, I., Attanzio, A., Tesoriere, L., Allegra, M., Garcia-Llatas, G., & Cilla, A. (2023). A Mixture of Dietary Plant Sterols at Nutritional Relevant Serum Concentration Inhibits Extrinsic Pathway of Eryptosis Induced by Cigarette Smoke Extract. International Journal of Molecular Sciences, 24(2), 1264. https://doi.org/10.3390/ijms24021264